
APOE ε4 in Depression-Associated Memory Impairment—Evidence from Genetic and MicroRNA Analyses

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Penotype Measures
2.2.1. Verbal Memory
2.2.2. Depression
2.2.3. Covariates
2.3. Omics Data
2.3.1. Genome-Wide SNP Chip
2.3.2. APOE Status
2.3.3. miRNA Measurement
2.4. Statistical Analyses
2.4.1. Main Analyses in TREND
2.4.2. Replication in LEGEND
2.4.3. Sensitivity Analyses for miRNAs in TREND Subsample
3. Results
3.1. Direct Effects of Depression and APOE ε4 Status on Verbal Memory in TREND
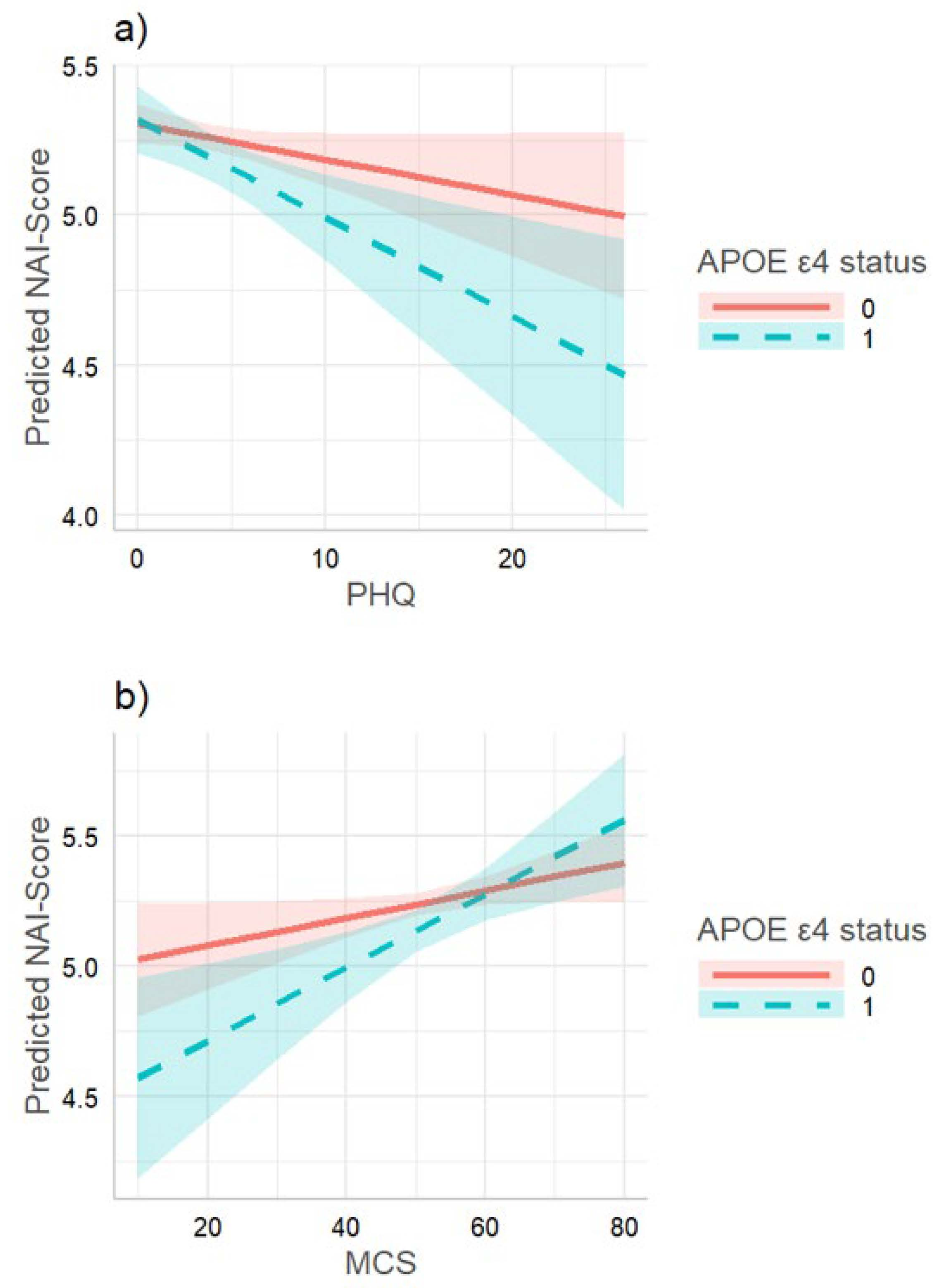
3.2. Interaction Effects between Depression and APOE ε4 Status on Verbal Memory in TREND
3.3. Replication of the Direct Effects and Interactions in LEGEND
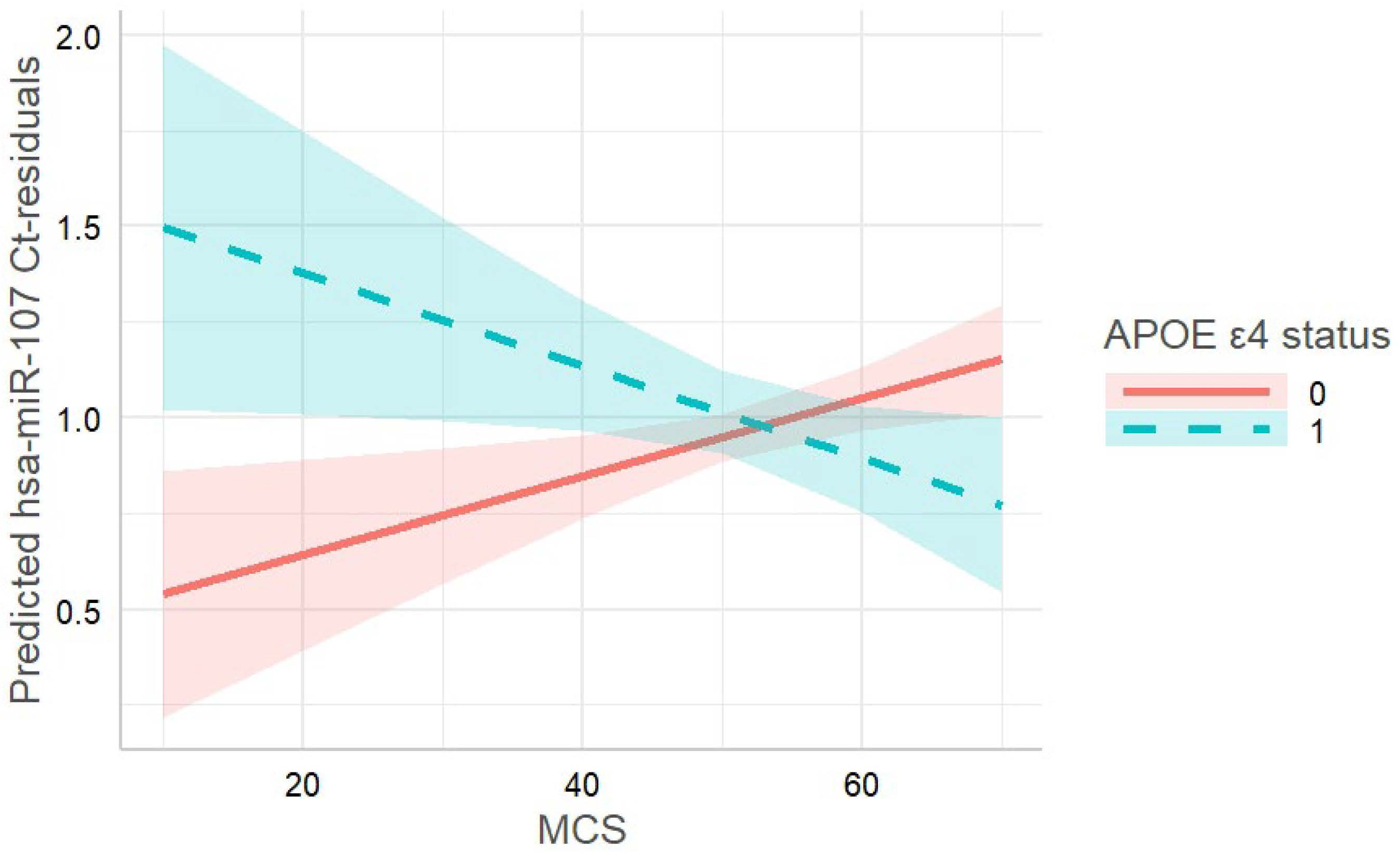
3.4. miRNA Analyses in the TREND Subsample
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hasin, D.S.; Sarvet, A.L.; Meyers, J.L.; Saha, T.D.; Ruan, W.J.; Stohl, M.; Grant, B.F. Epidemiology of Adult DSM-5 Major Depressive Disorder and Its Specifiers in the United States. JAMA Psychiatry 2018, 75, 336–346. [Google Scholar] [CrossRef] [Green Version]
- Fratelli, C.; Siqueira, J.; Silva, C.; Ferreira, E.; Silva, I. 5HTTLPR Genetic Variant and Major Depressive Disorder: A Review. Genes 2020, 11, 1260. [Google Scholar] [CrossRef] [PubMed]
- Woolf, C.; Lampit, A.; Shahnawaz, Z.; Sabates, J.; Norrie, L.M.; Burke, D.; Naismith, S.L.; Mowszowski, L. A Systematic Review and Meta-Analysis of Cognitive Training in Adults with Major Depressive Disorder. Neuropsychol. Rev. 2021, 32, 419–437. [Google Scholar] [CrossRef] [PubMed]
- Varghese, S.; Frey, B.N.; Schneider, M.A.; Kapczinski, F.; de Azevedo Cardoso, T. Functional and cognitive impairment in the first episode of depression: A systematic review. Acta Psychiatr. Scand. 2022, 145, 156–185. [Google Scholar] [CrossRef] [PubMed]
- Schmaal, L.; Hibar, D.P.; Sämann, P.G.; Hall, G.B.; Baune, B.T.; Jahanshad, N.; Cheung, J.W.; van Erp, T.G.M.; Bos, D.; Ikram, M.A.; et al. Cortical abnormalities in adults and adolescents with major depression based on brain scans from 20 cohorts worldwide in the ENIGMA Major Depressive Disorder Working Group. Mol. Psychiatry 2017, 22, 900–909. [Google Scholar] [CrossRef] [Green Version]
- El Haj, M.; Antoine, P.; Amouyel, P.; Lambert, J.-C.; Pasquier, F.; Kapogiannis, D. Apolipoprotein E (APOE) ε4 and episodic memory decline in Alzheimer’s disease: A review. Ageing Res. Rev. 2016, 27, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.-W.A.; Zhou, B.; Wernig, M.; Südhof, T.C. ApoE2, ApoE3, and ApoE4 Differentially Stimulate APP Transcription and Aβ Secretion. Cell 2017, 168, 427–441.e21. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, T.; Serrano-Pozo, A.; Hori, Y.; Adams, K.W.; Takeda, S.; Banerji, A.O.; Mitani, A.; Joyner, D.; Thyssen, D.H.; Bacskai, B.J.; et al. Apolipoprotein E, especially apolipoprotein E4, increases the oligomerization of amyloid β peptide. J. Neurosci. 2012, 32, 15181–15192. [Google Scholar] [CrossRef] [Green Version]
- Altuna, M.; Olmedo-Saura, G.; Carmona-Iragui, M.; Fortea, J. Mechanisms Involved in Epileptogenesis in Alzheimer’s Disease and Their Therapeutic Implications. Int. J. Mol. Sci. 2022, 23, 4307. [Google Scholar] [CrossRef]
- Forero, D.A.; Lopez-Leon, S.; Gonzales-Giraldo, Y.; Dries, D.R.; Pereira-Morales, A.J.; Jimenez, K.M.; Franco-Restrepo, J.E. APOE gene and neuropsychiatric disorders and endophenotypes: A comprehensive review. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2018, 177, 126–142. [Google Scholar] [CrossRef] [Green Version]
- Debette, S.; Ibrahim Verbaas, C.A.; Bressler, J.; Schuur, M.; Smith, A.; Bis, J.C.; Davies, G.; Wolf, C.; Gudnason, V.; Chibnik, L.B.; et al. Genome-wide studies of verbal declarative memory in nondemented older people: The Cohorts for Heart and Aging Research in Genomic Epidemiology consortium. Biol. Psychiatry 2015, 77, 749–763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, T.; Lee, H.; Young, A.H.; Aarsland, D.; Thuret, S. Adult Hippocampal Neurogenesis in Major Depressive Disorder and Alzheimer’s Disease. Trends Mol. Med. 2020, 26, 803–818. [Google Scholar] [CrossRef] [PubMed]
- Piers, R.J.; Liu, Y.; Ang, T.F.A.; Tao, Q.; Au, R.; Qiu, W.Q. Association between Elevated Depressive Symptoms and Cognitive Function Moderated by APOE4 Status: Framingham Offspring Study. J. Alzheimers Dis. 2021, 80, 1269–1279. [Google Scholar] [CrossRef] [PubMed]
- Camkurt, M.A.; Güneş, S.; Coşkun, S.; Fındıklı, E. Peripheral Signatures of Psychiatric Disorders: MicroRNAs. Clin. Psychopharmacol. Neurosci. 2017, 15, 313–319. [Google Scholar] [CrossRef] [Green Version]
- Place, R.F.; Li, L.-C.; Pookot, D.; Noonan, E.J.; Dahiya, R. MicroRNA-373 induces expression of genes with complementary promoter sequences. Proc. Natl. Acad. Sci. USA 2018, 115, E3325. [Google Scholar] [CrossRef] [Green Version]
- Mendes-Silva, A.P.; Pereira, K.S.; Tolentino-Araujo, G.T.; Nicolau, E.D.S.; Silva-Ferreira, C.M.; Teixeira, A.L.; Diniz, B.S. Shared Biologic Pathways between Alzheimer Disease and Major Depression: A Systematic Review of MicroRNA Expression Studies. Am. J. Geriatr. Psychiatry 2016, 24, 903–912. [Google Scholar] [CrossRef]
- Wingo, T.S.; Yang, J.; Fan, W.; Min Canon, S.; Gerasimov, E.S.; Lori, A.; Logsdon, B.; Yao, B.; Seyfried, N.T.; Lah, J.J.; et al. Brain microRNAs associated with late-life depressive symptoms are also associated with cognitive trajectory and dementia. NPJ Genom. Med. 2020, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Ogonowski, N.; Salcidua, S.; Leon, T.; Chamorro-Veloso, N.; Valls, C.; Avalos, C.; Bisquertt, A.; Rentería, M.E.; Orellana, P.; Duran-Aniotz, C. Systematic Review: MicroRNAs as Potential Biomarkers in Mild Cognitive Impairment Diagnosis. Front. Aging Neurosci. 2021, 13, 807764. [Google Scholar] [CrossRef]
- Völzke, H.; Schössow, J.; Schmidt, C.O.; Jürgens, C.; Richter, A.; Werner, A.; Werner, N.; Radke, D.; Teumer, A.; Ittermann, T.; et al. Cohort Profile Update: The Study of Health in Pomerania (SHIP). Int. J. Epidemiol. 2022, dyac034. [Google Scholar] [CrossRef]
- Schulz, A.; Becker, M.; van der Auwera, S.; Barnow, S.; Appel, K.; Mahler, J.; Schmidt, C.O.; John, U.; Freyberger, H.J.; Grabe, H.J. The impact of childhood trauma on depression: Does resilience matter? Population-based results from the Study of Health in Pomerania. J. Psychosom. Res. 2014, 77, 97–103. [Google Scholar] [CrossRef]
- Oswald, W.D.; Fleischmann, U.M. Nürnberger-Alters-Inventar: (NAI) NAI-Testmanual und Textband; Hogrefe: Göttingen, Germany, 1999. [Google Scholar]
- Van der Auwera, S.; Terock, J.; Teumer, A.; Schomerus, G.; Homuth, G.; Grabe, H.J. Sex effects for the interaction of dopamine related genetic variants for COMT and BDNF on declarative memory performance. Genes Brain Behav. 2021, 20, e12737. [Google Scholar] [CrossRef] [PubMed]
- Helmstaedter, C.; Durwen, H.F. VLMT: Verbaler Lern- und Merkfähigkeitstest: Ein Praktikables und Differenziertes Instrumentarium zur Prüfung der Verbalen Gedächtnisleistungen; Schweizer Archiv für Neurologie, Neurochirurgie und Psychiatrie: Muttenz, Switzerland, 1990. [Google Scholar]
- Wittchen, H.-L. Reliability and validity studies of the WHO-Composite International Diagnostic Interview (CIDI): A critical review. J. Psychiatr. Res. 1994, 28, 57–84. [Google Scholar] [CrossRef]
- Wittchen, H.U.; Lachner, G.; Wunderlich, U.; Pfister, H. Test-retest reliability of the computerized DSM-IV version of the Munich-Composite International Diagnostic Interview (M-CIDI). Soc. Psychiatry Psychiatr. Epidemiol. 1998, 33, 568–578. [Google Scholar] [CrossRef]
- Kroenke, K.; Spitzer, R.L.; Williams, J.B.W. The PHQ-9: Validity of a Brief Depression Severity Measure. J. Gen. Intern. Med. 2001, 16, 606–613. [Google Scholar] [CrossRef] [PubMed]
- Beck, A.T.; Steer, R.A.; Brown, G. Manual for the Beck Depression Inventory-II; Psychological Corporation: San Antonio, TX, USA, 1996. [Google Scholar]
- Vilagut, G.; Forero, C.G.; Pinto-Meza, A.; Haro, J.M.; de Graaf, R.; Bruffaerts, R.; Kovess, V.; de Girolamo, G.; Matschinger, H.; Ferrer, M.; et al. The mental component of the short-form 12 health survey (SF-12) as a measure of depressive disorders in the general population: Results with three alternative scoring methods. Value Health 2013, 16, 564–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ware, J.; Kosinski, M.; Keller, S.D. A 12-Item Short-Form Health Survey: Construction of Scales and Preliminary Tests of Reliability and Validity. Med. Care 1996, 34, 220–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habes, M.; Toledo, J.B.; Resnick, S.M.; Doshi, J.; van der Auwera, S.; Erus, G.; Janowitz, D.; Hegenscheid, K.; Homuth, G.; Völzke, H.; et al. Relationship between APOE Genotype and Structural MRI Measures throughout Adulthood in the Study of Health in Pomerania Population-Based Cohort. AJNR Am. J. Neuroradiol. 2016, 37, 1636–1642. [Google Scholar] [CrossRef] [Green Version]
- Pitchika, A.; Ricardo Paulista Markus, M.; Schipf, S.; Teumer, A.; van der Auwera, S.; Nauck, M.; Dörr, M.; Felix, S.; Jörgen Grabe, H.; Völzke, H.; et al. Longitudinal association of Apolipoprotein E polymorphism with lipid profile, type 2 diabetes and metabolic syndrome: Results from a 15 year follow-up study. Diabetes Res. Clin. Pract. 2022, 185, 109778. [Google Scholar] [CrossRef]
- Ameling, S.; Kacprowski, T.; Chilukoti, R.K.; Malsch, C.; Liebscher, V.; Suhre, K.; Pietzner, M.; Friedrich, N.; Homuth, G.; Hammer, E.; et al. Associations of circulating plasma microRNAs with age, body mass index and sex in a population-based study. BMC Med. Genom. 2015, 8, 61. [Google Scholar] [CrossRef] [Green Version]
- Van der Auwera, S.; Ameling, S.; Nauck, M.; Völzke, H.; Völker, U.; Grabe, H.J. Plasma circulating micro-RNAs associated with alexithymia reflect a high overlap on neuropsychiatric outcomes. J. Affect. Disord. 2022, 305, 206–212. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Keller, A.; Gröger, L.; Tschernig, T.; Solomon, J.; Laham, O.; Schaum, N.; Wagner, V.; Kern, F.; Schmartz, G.P.; Li, Y.; et al. miRNATissueAtlas2: An update to the human miRNA tissue atlas. Nucleic Acids Res. 2022, 50, D211–D221. [Google Scholar] [CrossRef] [PubMed]
- Karisetty, B.C.; Bhatnagar, A.; Armour, E.M.; Beaver, M.; Zhang, H.; Elefant, F. Amyloid-β Peptide Impact on Synaptic Function and Neuroepigenetic Gene Control Reveal New Therapeutic Strategies for Alzheimer’s Disease. Front. Mol. Neurosci. 2020, 13, 577622. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, U.; Nilson, A.N.; Kayed, R. The Role of Amyloid-β Oligomers in Toxicity, Propagation, and Immunotherapy. EBioMedicine 2016, 6, 42–49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayed, R.; Lasagna-Reeves, C.A. Molecular mechanisms of amyloid oligomers toxicity. J. Alzheimers. Dis. 2013, 33 (Suppl. 1), S67–S78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padmanabhan, P.; Kneynsberg, A.; Götz, J. Super-resolution microscopy: A closer look at synaptic dysfunction in Alzheimer disease. Nat. Rev. Neurosci. 2021, 22, 723–740. [Google Scholar] [CrossRef]
- Holderbach, R.; Clark, K.; Moreau, J.-L.; Bischofberger, J.; Normann, C. Enhanced long-term synaptic depression in an animal model of depression. Biol. Psychiatry 2007, 62, 92–100. [Google Scholar] [CrossRef]
- Wang, W.; Wei, C.; Quan, M.; Li, T.; Jia, J. Sulforaphane Reverses the Amyloid-β Oligomers Induced Depressive-like Behavior. J. Alzheimers. Dis. 2020, 78, 127–137. [Google Scholar] [CrossRef]
- Cirrito, J.R.; Wallace, C.E.; Yan, P.; Davis, T.A.; Gardiner, W.D.; Doherty, B.M.; King, D.; Yuede, C.M.; Lee, J.-M.; Sheline, Y.I. Effect of escitalopram on Aβ levels and plaque load in an Alzheimer mouse model. Neurology 2020, 95, e2666–e2674. [Google Scholar] [CrossRef]
- Keller, J.; Gomez, R.; Williams, G.; Lembke, A.; Lazzeroni, L.; Murphy, G.M.; Schatzberg, A.F. HPA axis in major depression: Cortisol, clinical symptomatology and genetic variation predict cognition. Mol. Psychiatry 2017, 22, 527–536. [Google Scholar] [CrossRef]
- Yan, T.; Qiu, Y.; Yu, X.; Yang, L. Glymphatic Dysfunction: A Bridge between Sleep Disturbance and Mood Disorders. Front. Psychiatry 2021, 12, 658340. [Google Scholar] [CrossRef]
- Kumar, A.; Sidhu, J.; Goyal, A.; Tsao, J.W. Alzheimer Disease; StatPearls: Treasure Island, FL, USA, 2022. [Google Scholar]
- Norris, D. Short-term memory and long-term memory are still different. Psychol. Bull. 2017, 143, 992–1009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Jaber, V.; Alexandrov, P.N.; Vergallo, A.; Lista, S.; Hampel, H.; Lukiw, W.J. microRNA-Based Biomarkers in Alzheimer’s Disease (AD). Front. Neurosci. 2020, 14, 585432. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zhu, Y.; Chen, W.; Tang, Y. Emerging role of microRNAs in major depressive disorder and its implication on diagnosis and therapeutic response. J. Affect. Disord. 2021, 286, 80–86. [Google Scholar] [CrossRef]
- Van den Berg, M.M.J.; Krauskopf, J.; Ramaekers, J.G.; Kleinjans, J.C.S.; Prickaerts, J.; Briedé, J.J. Circulating microRNAs as potential biomarkers for psychiatric and neurodegenerative disorders. Prog. Neurobiol. 2020, 185, 101732. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.-X.; Rajeev, B.W.; Stromberg, A.J.; Ren, N.; Tang, G.; Huang, Q.; Rigoutsos, I.; Nelson, P.T. The expression of microRNA miR-107 decreases early in Alzheimer’s disease and may accelerate disease progression through regulation of beta-site amyloid precursor protein-cleaving enzyme 1. J. Neurosci. 2008, 28, 1213–1223. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Kong, L.; Yao, Y.; Li, S.; Tao, Z.; Yan, Y.; Yang, J. Osthole decreases beta amyloid levels through up-regulation of miR-107 in Alzheimer’s disease. Neuropharmacology 2016, 108, 332–344. [Google Scholar] [CrossRef] [PubMed]
- Das, B.; Singh, N.; Yao, A.Y.; Zhou, J.; He, W.; Hu, X.; Yan, R. BACE1 controls synaptic function through modulating release of synaptic vesicles. Mol. Psychiatry 2021, 26, 6394–6410. [Google Scholar] [CrossRef]
- Turk, A.; Kunej, T.; Peterlin, B. MicroRNA-Target Interaction Regulatory Network in Alzheimer’s Disease. J. Pers. Med. 2021, 11, 1275. [Google Scholar] [CrossRef]
- Thalamuthu, A.; Mills, N.T.; Berger, K.; Minnerup, H.; Grotegerd, D.; Dannlowski, U.; Meinert, S.; Opel, N.; Repple, J.; Gruber, M.; et al. Genome-wide interaction study with major depression identifies novel variants associated with cognitive function. Mol. Psychiatry 2021, 27, 1111–1119. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Variable | TREND (n = 3964) | LEGEND (n = 2322) | Comparison |
|---|---|---|---|
| Age | 51.9 (15.4), [20–83] | 55.9 (14.3), [29–89] | T = −9.06, p < 2.2 × 10−16 |
| Sex | Chi2 = 0.45, p = 0.50 | ||
| Males | 1919 (48.4%) | 1103 (47.5%) | |
| Females | 2045 (51.6%) | 1219 (52.5%) | |
| Education | Chi2 = 45.07, p = 1.64 × 10−10 | ||
| <10 years | 909 (22.9%) | 677 (29.2%) | |
| =10 years | 2052 (51.8%) | 1196 (51.5%) | |
| >10 years | 1003 (25.3%) | 449 (19.3%) | |
| APOE ε4 status | Chi2 = 3.55, p = 0.06 | ||
| ε4 carrier | 951 (24%) | 508 (21.9%) | |
| non carrier | 3013 (76%) | 1814 (78.1%) | |
| MDD | Chi2 = 2.38, p = 0.12 | ||
| No | 3153 (81.5%) | 1882 (83.1%) | |
| Yes | 718 (18.5%) | 384 (16.9%) | |
| NA | 93 | 56 | |
| Mental health score (MCS) | 52.5 (8.6), [16.6–70.3] | 51.9 (9.3), [15.6–70.8] | T = 2.72, p = 0.007 |
| NA | 93 | 0 | |
| Depressive symptoms (PHQ–9) | 3.9 (3.6),[0–26] | - | |
| NA | 208 | ||
| Depressive symptoms (BDI-II) | - | 6.5 (7.3), [0–54.6] | |
| NA | 91 | ||
| NAI | - | ||
| Immediate recall | 5.2 (1.4), [0–8] | ||
| NA | 3 | ||
| NAI | - | ||
| Delayed recall | 5.7 (1.7), [−3–8] | ||
| NA | 28 | ||
| VLMT | - | ||
| Immediate recall | 24.4 (6.4), [1–45] | ||
| NA | 142 | ||
| VLMT | - | ||
| Delayed recall | 7.9 (3.1), [0–15] | ||
| NA | 236 | ||
| Smoking status | Chi2 = 32.26, p = 9.9 × 10∓8 | ||
| Never | 1444 (36.4%) | 720 (38.3%) | |
| Former | 1461 (36.9%) | 783 (41.7%) | |
| Current | 1059 (26.7%) | 376 (20.0%) | |
| Diabetes | Chi2 = 10.74, p = 0.001 | ||
| No | 1968 (85.1%) | ||
| Yes | 3490 (88%) | 344 (14.9%) | |
| NA | 474 (12%) | 10 | |
| Alcohol (beverage last 12 month) | - | ||
| Never | 430 (10.8%) | ||
| once per month | 1093 (27.6%) | ||
| 2–4 per month | 1247 (31.5%) | ||
| 2–3 per week | 791 (20.0%) | ||
| >3 per week | 403 (10.8%) | ||
| Hypertension | Chi2 = 34.74, p = 3.8 × 10−9 | ||
| No | 1046 (55.9%) | ||
| Yes | 2076 (52.4%) | 824 (44.1%) | |
| NA | 1888 (47.6%) | 452 | |
| Heart disease | Chi2 = 1026, p < 2.2 × 10−16 | ||
| No | 929 (45.4%) | ||
| Yes | 3363 (84.8%) | 1117 (54.6%) | |
| NA | 601 (15.2%) | 443 | |
| Time between SHIP-START-2 and LEGEND in years | 1.6 (1.2), [−1.8–4.9] |
| Adjustment | Predictor | Immediate Recall n, β, p-Value | Delayed Recall n, β, p-Value |
|---|---|---|---|
| Covariate set 1 | PHQ-9 | 3753, −0.0198, 0.00026 | 3731, −0.022, 0.0027 |
| MCS | 3869, 0.008, 0.00024 | 3845, 0.008, 0.0088 | |
| MDD | 3868, −0.012, 0.812 | 3844, −0.052, 0.443 | |
| APOE ε4 | 3964, −0.08, 0.073 | 3936, −0.10, 0.097 | |
| Covariate set 2 | PHQ-9 | 3753, −0.018, 0.0127 | 3731, −0.021, 0.005 |
| MCS | 3869, 0.0073, 0.0023 | 3845, 0.007, 0.017 | |
| MDD | 3868, 0.00012, 0.998 | 3844, −0.052, 0.436 | |
| APOE ε4 | 3964, −0.083, 0.0596 | 3936, −0.1, 0.094 |
| Adjustment | Interaction Term | Immediate Recall n, β, p-Value | Delayed Recall n, β, p-Value |
|---|---|---|---|
| Covariate set 1 | PHQ-9 × APOE ε4 | 3753, −0.019, 0.106 | 3731, −0.006, 0.72 |
| MCS × APOE ε4 | 3869, 0.009, 0.097 | 3845, −0.003, 0.704 | |
| MDD × APOE ε4 | 3868, 0.013, 0.91 | 3844, −0.033, 0.83 | |
| Covariate set 2 | PHQ-9 × APOE ε4 | 3753, −0.021, 0.083 | 3731, −0.008, 0.633 |
| MCS × APOE ε4 | 3869, 0.009, 0.087 | 3845, −0.002, 0.77 | |
| MDD × APOE ε4 | 3868, 0.007, 0.951 | 3844, −0042, 0.78 |
| Predictor | Immediate Recall n, β, p-Value | Delayed Recall n, β, p-Value |
|---|---|---|
| BDI-II | 2132, −0.06, 0.00016 | 2046, −0.025, 0.0045 |
| MCS | 2180, 0.032, 0.01 | 2086, 0.016, 0.017 |
| MDD | 2162, 0.244, 0.42 | 2078, −0.06, 0.73 |
| APOE ε4 | 2180, 0.23, 0.41 | 2086, 0.066, 0.677 |
| Interaction term | ||
| BDI × APOE ε4 | 2132, −0.066, 0.075 | 2046, −0.027, 0.167 |
| MCS × APOE ε4 | 2180, 0.0041, 0.88 | 2086, −0.0045, 0.76 |
| MDD × APOE ε4 | 2162, −1.22, 0.12 | 2078, −0.53, 0.21 |
| Predictor | Immediate Recall n, β, p-Value | Delayed Recall n, β, p-Value |
|---|---|---|
| BDI-II | 1755, −0.04, 0.019 | 1685, −0.021, 0.041 |
| MCS | 1778, 0.025, 0.078 | 1704, 0.015, 0.047 |
| MDD | 1767, 0.36, 0.31 | 1697, −0.19, 0.32 |
| APOE ε4 | 1778, 0.11, 0.72 | 1704, −0.034, 0.83 |
| Interaction term | ||
| BDI × APOE ε4 | 1755, −0.057, 0.172 | 1685, −0.013, 0.56 |
| MCS × APOE ε4 | 1778, 0.005, 0.86 | 1704, 0.0003, 0.99 |
| MDD × APOE ε4 | 1767, −1.16, 0.17 | 1697, −0.64, 0.18 |
| Micro RNA | APOE ε4*PHQ-9 β, p-Value | APOE ε4*MCS β, p-Value |
|---|---|---|
| hsa-miR-221-3p | −0.094, 0.002 | 0.027, 0.029 |
| hsa-miR-376a-3p | −0.094, 0.005 | 0.036, 0.012 |
| hsa-let-7d-3p | −0.042, 0.004 | 0.015, 0.018 |
| hsa-miR-107 | 0.024, 0.016 | −0.027, 0.0001 |
| hsa-miR-382-5p | −0.084,0.011 | 0.043, 0.002 |
| hsa-miR-181a-5p | 0.041, 0.069 | −0.028, 0.005 |
| hsa-miR-99a-5p | 0.036, 0.098 | −0.026, 0.007 |
| hsa-miR-222-3p | 0.006, 0.678 | −0.017, 0.009 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bonk, S.; Kirchner, K.; Ameling, S.; Garvert, L.; Völzke, H.; Nauck, M.; Völker, U.; Grabe, H.J.; Van der Auwera, S. APOE ε4 in Depression-Associated Memory Impairment—Evidence from Genetic and MicroRNA Analyses. Biomedicines 2022, 10, 1560. https://doi.org/10.3390/biomedicines10071560
Bonk S, Kirchner K, Ameling S, Garvert L, Völzke H, Nauck M, Völker U, Grabe HJ, Van der Auwera S. APOE ε4 in Depression-Associated Memory Impairment—Evidence from Genetic and MicroRNA Analyses. Biomedicines. 2022; 10(7):1560. https://doi.org/10.3390/biomedicines10071560
Chicago/Turabian StyleBonk, Sarah, Kevin Kirchner, Sabine Ameling, Linda Garvert, Henry Völzke, Matthias Nauck, Uwe Völker, Hans J. Grabe, and Sandra Van der Auwera. 2022. "APOE ε4 in Depression-Associated Memory Impairment—Evidence from Genetic and MicroRNA Analyses" Biomedicines 10, no. 7: 1560. https://doi.org/10.3390/biomedicines10071560