
Deletion of Toxoplasma Rhoptry Protein 38 (PruΔrop38) as a Vaccine Candidate for Toxoplasmosis in a Murine Model


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Ethical Approval
2.2. Parasites
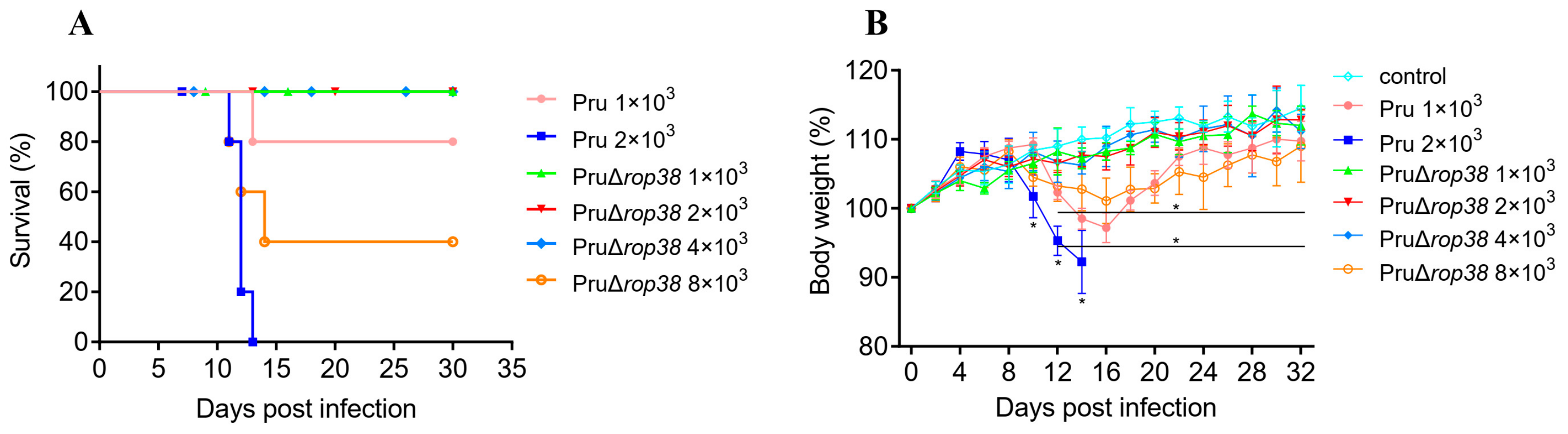
2.3. Virulence Comparison of PruΔrop38 and Pru Strain
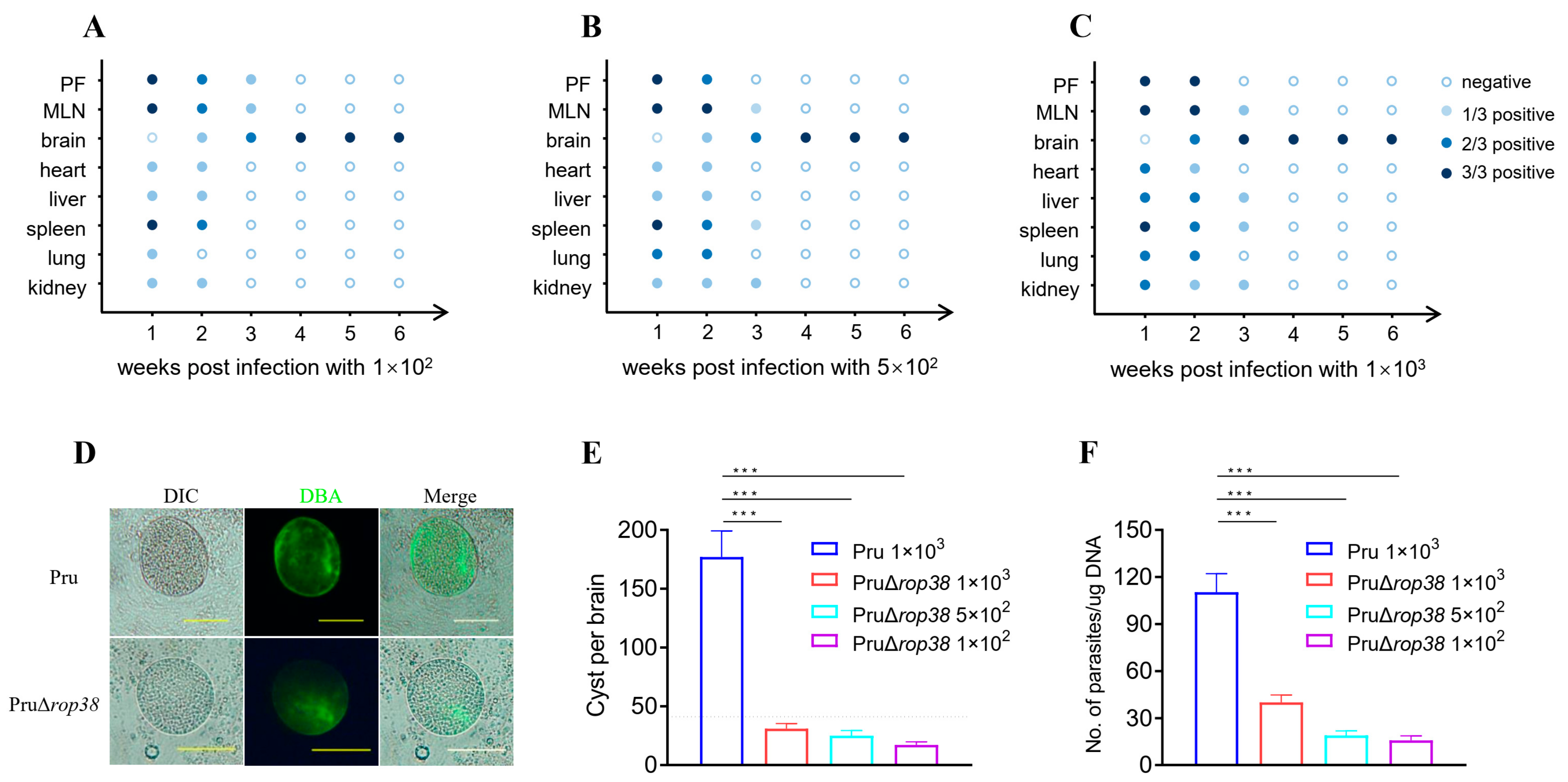
2.4. Dynamic Distribution of PruΔrop38 In Vivo
2.5. Brain Parasite Burden Assay
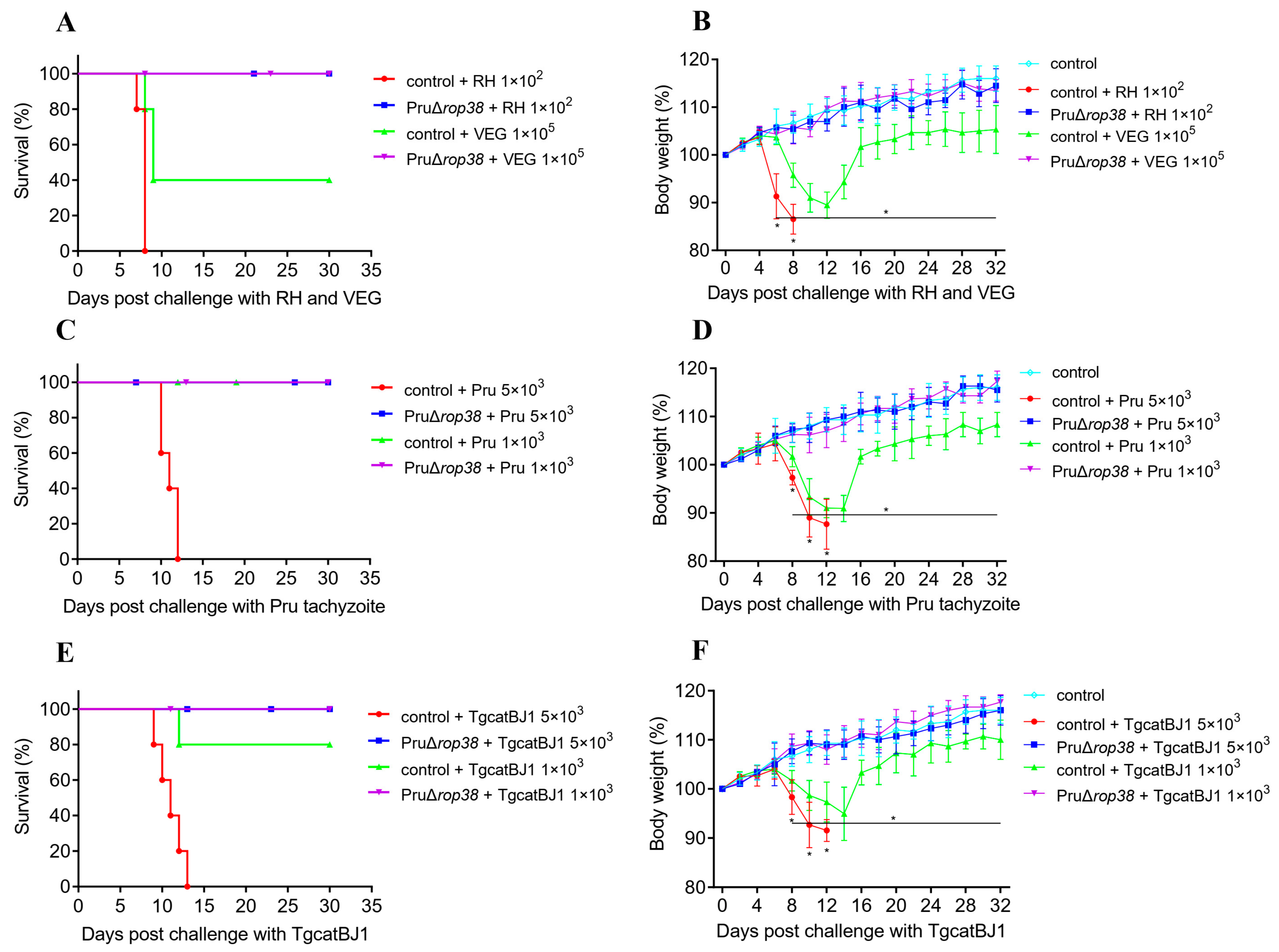
2.6. Immunization and Challenges
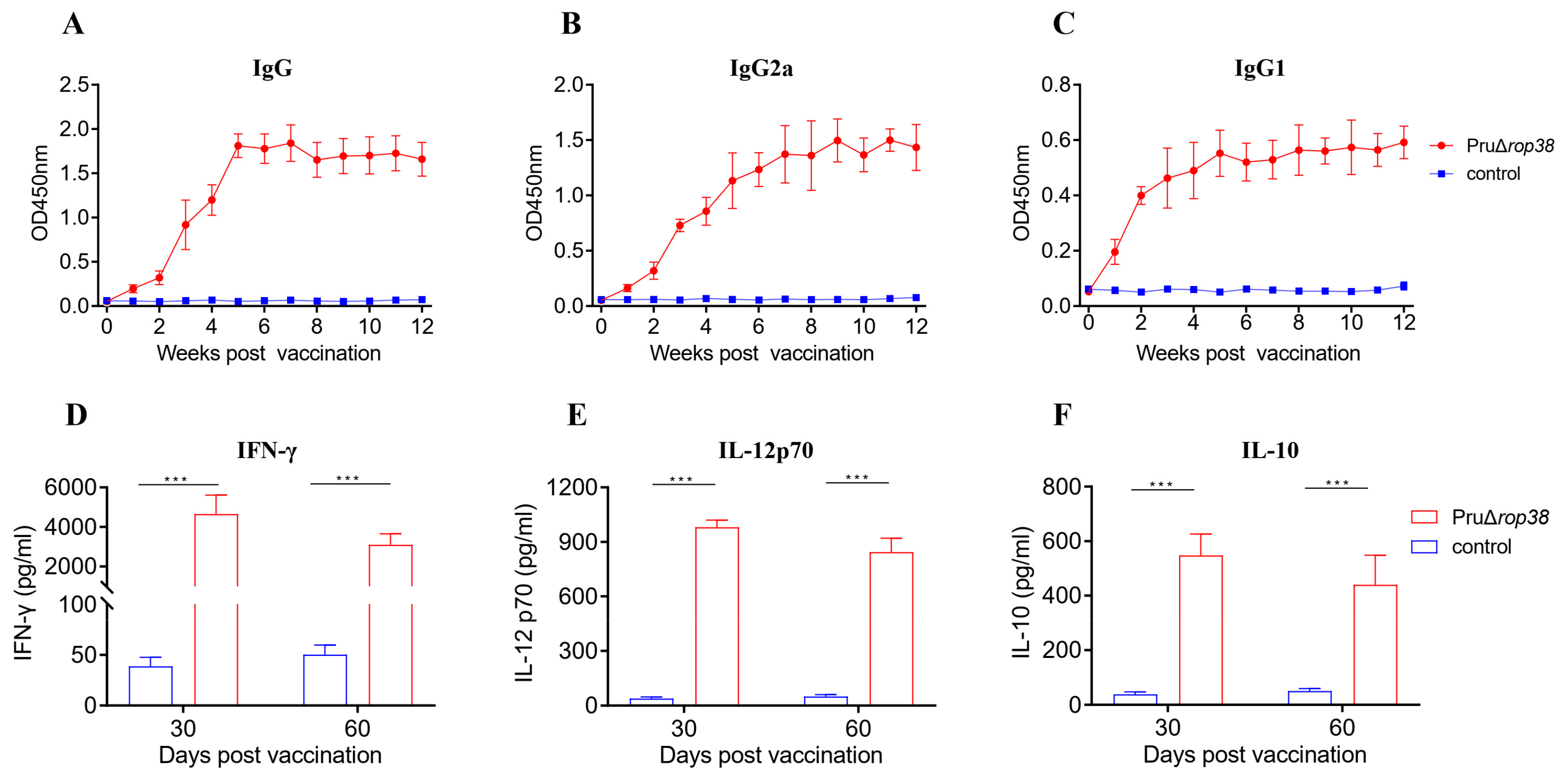
2.7. Toxoplasma IgG Antibody Assay
2.8. Cytokine Production Assay
2.9. Statistical Analysis
3. Results
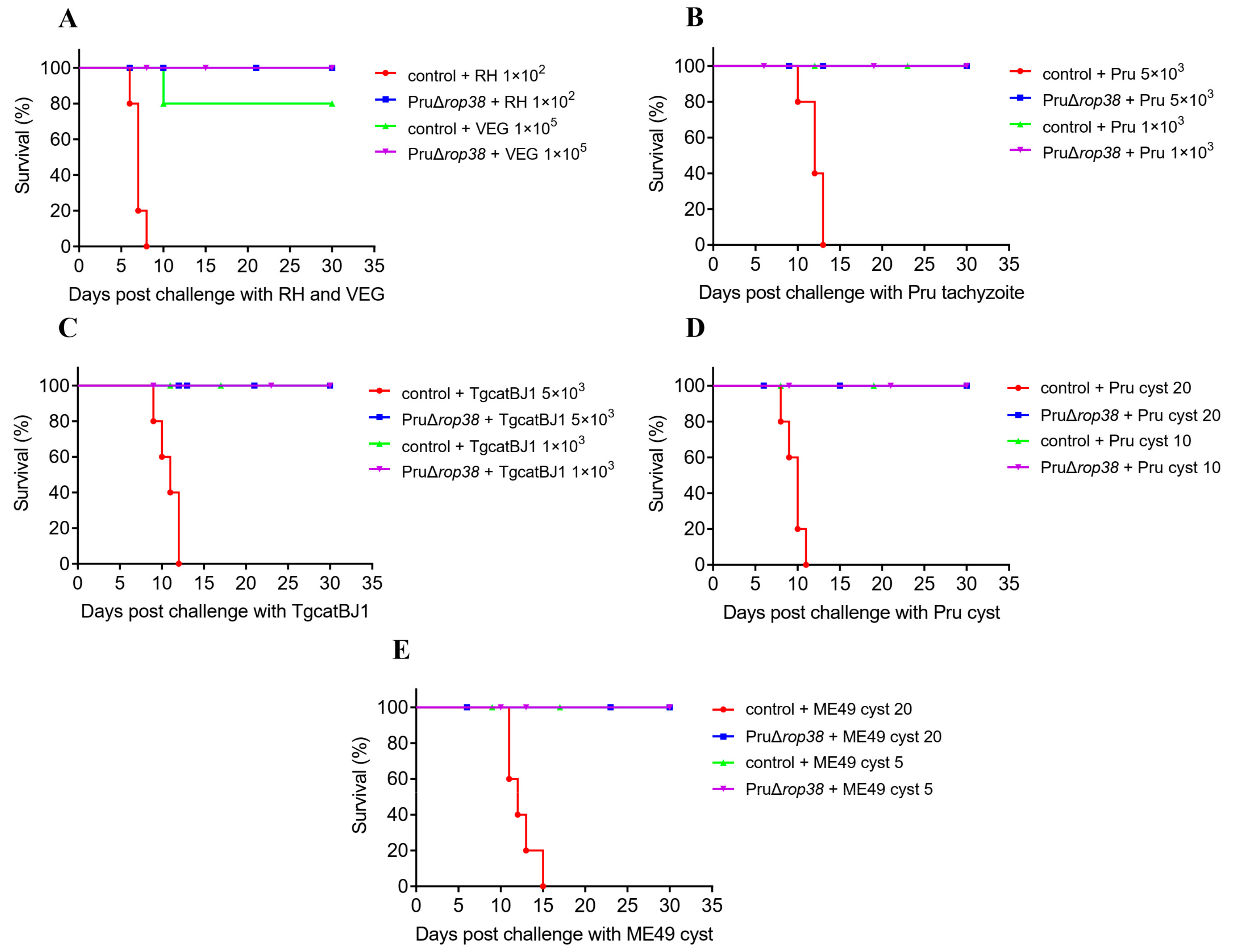
3.1. PruΔrop38 Exhibits Significantly Reduced Pathogenicity in Mice
3.2. Dynamic Distribution of PruΔrop38 in Mice
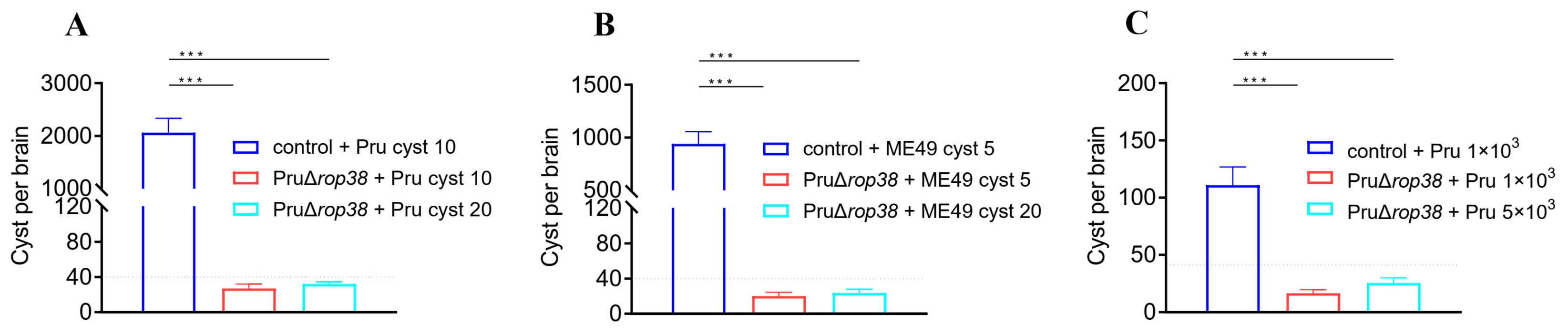
3.3. Deletion of TgROP38 Significantly Reduces Brain Parasite Burden
3.4. Humoral Immune Responses Induced by PruΔrop38
3.5. Cellular Immune Responses Induced by PruΔrop38
3.6. Protection against Varied Genotypes of Tachyzoites
3.7. Protection against Pru and ME49 Cyst
3.8. Significant Reduction in Brain Parasite Burden in Challenging Type II Strains
3.9. Vaccination with PruΔrop38 Provides Long-Time Protection in Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sanchez, S.G.; Besteiro, S. The pathogenicity and virulence of Toxoplasma gondii. Virulence 2021, 12, 3095–3114. [Google Scholar] [CrossRef]
- Yarovinsky, F. Innate immunity to Toxoplasma gondii infection. Nat. Rev. Immunol. 2014, 14, 109–121. [Google Scholar] [CrossRef] [PubMed]
- Izadi, M.; Jafari, N.J.; Poornaki, A.M.; Sadraei, J.; Rezavand, B.; Mirzaei, H.R.; Zarrinfar, H.; Khedive, A. Detection of Toxoplasma gondii from Clinical Specimens of Patients Receiving Renal Transplant Using ELISA and PCR. Nephro-Urol. Mon. 2013, 5, 983–987. [Google Scholar] [CrossRef] [Green Version]
- Matta, S.K.; Rinkenberger, N.; Dunay, I.R.; Sibley, L.D. Toxoplasma gondii infection and its implications within the central nervous system. Nat. Rev. Microbiol. 2021, 19, 467–480. [Google Scholar] [CrossRef] [PubMed]
- Tenter, A.M.; Heckeroth, A.R.; Weiss, L.M. Toxoplasma gondii: From animals to humans. Int. J. Parasitol. 2000, 30, 1217–1258. [Google Scholar] [CrossRef] [Green Version]
- Dunay, I.R.; Gajurel, K.; Dhakal, R.; Liesenfeld, O.; Montoya, J.G. Treatment of toxoplasmosis: Historical perspective, animal models, and current clinical practice. Clin. Microbiol. Rev. 2018, 31, e00057-17. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Zhang, N.; Li, T.; He, J.; Elsheikha, H.M.; Zhu, X. Advances in the development of anti-Toxoplasma gondii vaccines: Challenges, opportunities, and perspectives. Trends Parasitol. 2019, 35, 239–253. [Google Scholar] [CrossRef] [PubMed]
- Mévélec, M.-N.; Lakhrif, Z.; Dimier-Poisson, I. Key limitations and new insights into the parasite stage switching for future vaccine development in human, livestock, and cats. Front. Cell. Infect. Microbiol. 2020, 10, 607198. [Google Scholar] [CrossRef]
- Buxton, D.; Thomson, K.; Maley, S.; Wright, S.; Bos, H.J. Vaccination of sheep with a live incomplete strain (S48) of Toxoplasma gondii and their immunity to challenge when pregnant. Vet. Rec. 1991, 129, 89–93. [Google Scholar] [CrossRef]
- Shen, B.; Brown, K.M.; Lee, T.D.; Sibley, L.D. Efficient gene disruption in diverse strains of Toxoplasma gondii using CRISPR/CAS9. mBio 2014, 5, e01114-14. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Li, T.; Elsheikha, H.M.; Chen, K.; Cong, W.; Yang, W.; Bai, M.; Huang, S.; Zhu, X. Live attenuated Pru:Δcdpk2 strain of Toxoplasma gondii protects against acute, chronic, and congenital toxoplasmosis. J. Infect. Dis. 2018, 218, 768–777. [Google Scholar] [CrossRef] [Green Version]
- Lagal, V.; Dinis, M.; Cannella, D.; Bargieri, D.; Gonzalez, V.; Andenmatten, N.; Meissner, M.; Tardieux, I. AMA1-deficient Toxoplasma gondii parasites transiently colonize mice and trigger an innate immune response that leads to long-lasting protective immunity. Infect. Immun. 2015, 83, 2475–2486. [Google Scholar] [CrossRef] [Green Version]
- Xia, N.; Zhou, T.; Liang, X.; Ye, S.; Zhao, P.; Yang, J.; Zhou, Y.; Zhao, J.; Shen, B. A lactate fermentation mutant of Toxoplasma stimulates protective immunity against acute and chronic toxoplasmosis. Front. Immunol. 2018, 9, 1814. [Google Scholar] [CrossRef] [Green Version]
- Fox, B.A.; Bzik, D.J. Avirulent uracil auxotrophs based on disruption of orotidine-5′-monophosphate decarboxylase elicit protective immunity to Toxoplasma gondii. Infect. Immun. 2010, 78, 3744–3752. [Google Scholar] [CrossRef] [Green Version]
- Hunter, C.A.; Sibley, L.D. Modulation of innate immunity by Toxoplasma gondii virulence effectors. Nat. Rev. Microbiol. 2012, 10, 766–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Y.; Wang, X.; Liu, J.; Fu, Y.; Xu, J.; Liu, Q. Toxoplasma gondii rhoptry protein38 (TgROP38) affects parasite invasion, egress, and induces IL-18 secretion during early infection. Acta Biochim. Biophys. Sin. 2018, 50, 766–775. [Google Scholar] [CrossRef] [Green Version]
- Fox, B.A.; Rommereim, L.M.; Guevara, R.B.; Falla, A.; Hortua Triana, M.A.; Sun, Y.; Bzik, D.J. The Toxoplasma gondii rhoptry kinome is essential for chronic infection. mBio 2016, 7, e00193-16. [Google Scholar] [CrossRef] [Green Version]
- Qian, W.; Wang, H.; Su, C.; Shan, D.; Cui, X.; Yang, N.; Lv, C.; Liu, Q. Isolation and characterization of Toxoplasma gondii strains from stray cats revealed a single genotype in Beijing, China. Vet. Parasitol. 2012, 187, 408–413. [Google Scholar] [CrossRef]
- Mahmoud, M.E.; Ihara, F.; Fereig, R.M.; Nishimura, M.; Nishikawa, Y. Induction of depression-related behaviors by reactivation of chronic Toxoplasma gondii infection in mice. Behav. Brain Res. 2016, 298, 125–133. [Google Scholar] [CrossRef]
- Homan, W.L.; Vercammen, M.; De Braekeleer, J.; Verschueren, H. Identification of a 200- to 300-fold repetitive 529 bp DNA fragment in Toxoplasma gondii, and its use for diagnostic and quantitative PCR. Int. J. Parasitol. 2000, 30, 69–75. [Google Scholar] [CrossRef]
- Burrells, A.; Bartley, P.M.; Zimmer, I.A.; Roy, S.; Kitchener, A.C.; Meredith, A.; Wright, S.E.; Innes, E.A.; Katzer, F. Evidence of the three main clonal Toxoplasma gondii lineages from wild mammalian carnivores in the UK. Parasitology 2013, 140, 1768–1776. [Google Scholar] [CrossRef]
- Jones, C.D.; Okhravi, N.; Adamson, P.; Tasker, S.; Lightman, S. Comparison of PCR detection methods for B1, P30, and 18S rDNA genes of T. gondii in aqueous humor. Invest. Ophthalmol. Vis. 2000, 41, 634–644. [Google Scholar] [CrossRef]
- Fox, B.A.; Falla, A.; Rommereim, L.M.; Tomita, T.; Gigley, J.P.; Mercier, C.; Cesbron-Delauw, M.-F.; Weiss, L.M.; Bzik, D.J. Type II Toxoplasma gondii KU80 knockout strains enable functional analysis of genes required for cyst development and latent infection. Eukaryot. cell 2011, 10, 1193–1206. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Lin, M.; Li, M.; Yang, X.; Liu, J.; Liu, Q. Toxoplasma gondii metacaspase 2 is an important factor that influences bradyzoite formation in the Pru strain. Parasitol. Res. 2020, 119, 2287–2298. [Google Scholar] [CrossRef]
- Lourenco, E.V.; Pereira, S.R.; Faca, V.M.; Coelho-Castelo, A.A.M.; Mineo, J.R.; Roque-Barreira, M.C.; Greene, L.J.; Panunto-Castelo, A. Toxoplasma gondii micronemal protein MIC1 is a lactose-binding lectin. Glycobiology 2001, 11, 541–547. [Google Scholar] [CrossRef] [Green Version]
- Sayles, P.C.; Gibson, G.W.; Johnson, L.L. B cells are essential for vaccination-induced resistance to virulent Toxoplasma gondii. Infect. Immun. 2000, 68, 1026–1033. [Google Scholar] [CrossRef] [Green Version]
- Yarovinsky, F.; Zhang, D.; Andersen, J.F.; Bannenberg, G.L.; Serhan, C.N.; Hayden, M.S.; Hieny, S.; Sutterwala, F.S.; Flavell, R.A.; Ghosh, S.; et al. TLR11 activation of dendritic cells by a protozoan profilin-like protein. Science 2005, 308, 1626–1629. [Google Scholar] [CrossRef] [Green Version]
- Plattner, F.; Yarovinsky, F.; Romero, S.; Didry, D.; Carlier, M.F.; Sher, A.; Soldati-Favre, D. Toxoplasma profilin is essential for host cell invasion and TLR11-dependent induction of an interleukin-12 response. Cell Host Microbe 2008, 3, 77–87. [Google Scholar] [CrossRef] [Green Version]
- Mordue, D.G.; Monroy, F.; La Regina, M.; Dinarello, C.A.; Sibley, L.D. Acute toxoplasmosis leads to lethal overproduction of Th1 cytokines. J. Immunol. 2001, 167, 4574–4584. [Google Scholar] [CrossRef] [Green Version]
- Su, C.; Khan, A.; Zhou, P.; Majumdar, D.; Ajzenberg, D.; Dardé, M.-L.; Zhu, X.-Q.; Ajioka, J.W.; Rosenthal, B.M.; Dubey, J.P.; et al. Globally diverse Toxoplasma gondii isolates comprise six major clades originating from a small number of distinct ancestral lineages. Proc. Natl. Acad. Sci. USA 2012, 109, 5844–5849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galal, L.; Hamidović, A.; Dardé, M.L.; Mercier, M. Diversity of strains at the global level and its determinants. Food Waterborne Parasitol. 2019, 15, e00052. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.; Sun, L.; Elsheikha, H.M.; Cao, X.; Nie, L.; Li, T.; Li, T.; Zhu, X.; Wang, J. RHΔgra17Δnpt1 strain of Toxoplasma gondii elicits protective immunity against acute, chronic and congenital toxoplasmosis in mice. Microorganisms 2020, 8, 352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Yang, C.; Qian, J.; Li, F.; Zhao, J.; Fang, R. Toxoplasma gondii α-amylase deletion mutant is a promising vaccine against acute and chronic toxoplasmosis. Microb. Biotechnol. 2020, 13, 2057–2069. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Zhou, Z.; Ying, Z.; Xu, Y.; Liu, J.; Liu, Q. Deletion of Toxoplasma Rhoptry Protein 38 (PruΔrop38) as a Vaccine Candidate for Toxoplasmosis in a Murine Model. Biomedicines 2022, 10, 1336. https://doi.org/10.3390/biomedicines10061336
Wu Y, Zhou Z, Ying Z, Xu Y, Liu J, Liu Q. Deletion of Toxoplasma Rhoptry Protein 38 (PruΔrop38) as a Vaccine Candidate for Toxoplasmosis in a Murine Model. Biomedicines. 2022; 10(6):1336. https://doi.org/10.3390/biomedicines10061336
Chicago/Turabian StyleWu, Yayun, Zihui Zhou, Zhu Ying, Ying Xu, Jing Liu, and Qun Liu. 2022. "Deletion of Toxoplasma Rhoptry Protein 38 (PruΔrop38) as a Vaccine Candidate for Toxoplasmosis in a Murine Model" Biomedicines 10, no. 6: 1336. https://doi.org/10.3390/biomedicines10061336