
Tendon Healing Response Is Dependent on Epithelial–Mesenchymal–Tendon Transition State of Amniotic Epithelial Stem Cells

 ,
,  , ,
, ,  , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. AEC Isolation and Culture
2.3. AEC In Vitro Genotype Characterization
2.4. Phenotype Assessment of the Different AEC Experimental Groups
2.5. Comparison of In Vitro Immunomodulatory Properties of eAECs, mAECs vs. tdAECs
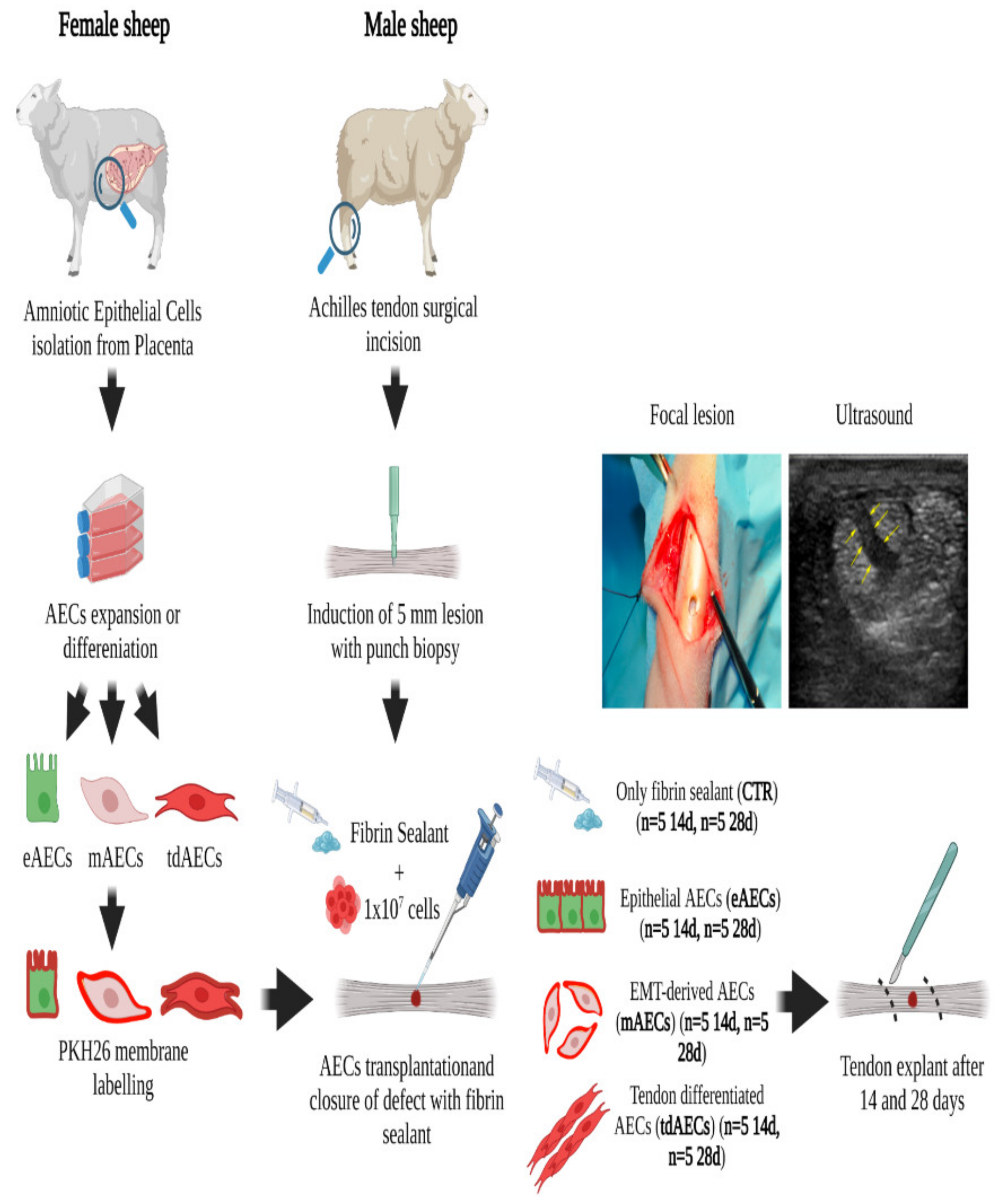
2.6. Ovine Achilles Tendon Injury Model
2.7. Gene Profiles of the Healing Tendons after Explant
2.7.1. Laser Capture Microdissection (LCM) Technique
2.7.2. Total RNA Extraction and RT-qPCR for Tendon Explants
2.8. Protein Profiles of the Healing Tendons after Explant
2.8.1. ECM Analysis of Tendon Explants: Histology and Immunohistochemistry
2.8.2. Histomorphometric Analyses on Tendon Explants
- Cellularity;
- Cell alignment;
- CD86- (M1 macrophages) and CD206-positive (M2 macrophages) cells.
2.9. Statistical Analysis
3. Results
3.1. In Vitro Production of Epithelial, Mesenchymal, and Tendon-like Committed Typologies of AECs
3.2. The Three Typologies of AECs Differed for In Vitro Immunomodulatory Properties
3.3. AEC Allotransplantation in Ovine Achilles Injured Tendon
3.4. AEC Phenotype Subsets Strongly Impact the Expression of Tendon-Related and ECM Gene Markers
3.5. AECs’ Allotransplantation Promoted the Early Tendon ECM Healing with Higher Performances in eAEC- and tdAEC-Treated Tissues
3.6. eAEC, mAEC, and tdAEC Allotransplantation Improves Cellularity and Cell Alignment in Injured Tendons
3.7. AEC Subsets’ Direct Contribution to Tendon Regeneration by Differentiating into Tenocytes
3.8. Inflammatory Phase Is Positively Modulated by eAEC, mAEC, and tdAEC Allotransplantation
3.9. Allotransplantation Modified the Response of the Injured Host Tissue: The Total Histological Score Assessment
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barboni, B.; Russo, V.; Berardinelli, P.; Mauro, A.; Valbonetti, L.; Sanyal, H.; Canciello, A.; Greco, L.; Muttini, A.; Gatta, V.; et al. Placental Stem Cells from Domestic Animals: Translational Potential and Clinical Relevance. Cell Transpl. 2018, 27, 93–116. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Lai, D. Application of Human Amniotic Epithelial Cells in Regenerative Medicine: A Systematic Review. Stem Cell Res. Ther. 2020, 11, 439. [Google Scholar] [CrossRef] [PubMed]
- Yang, P.; Yuan, W.; Liu, J.; Li, J.; Tan, B.; Qiu, C.; Zhu, X.; Qiu, C.; Lai, D.; Guo, L.; et al. Biological Characterization of Human Amniotic Epithelial Cells in a Serum-Free System and Their Safety Evaluation. Acta Pharm. Sin. 2018, 39, 1305–1316. [Google Scholar] [CrossRef] [PubMed]
- Wassmer, C.-H.; Berishvili, E. Immunomodulatory Properties of Amniotic Membrane Derivatives and Their Potential in Regenerative Medicine. Curr. Diabetes Rep. 2020, 20, 31. [Google Scholar] [CrossRef]
- Russo, V.; Mauro, A.; Martelli, A.; Di Giacinto, O.; Di Marcantonio, L.; Nardinocchi, D.; Berardinelli, P.; Barboni, B. Cellular and Molecular Maturation in Fetal and Adult Ovine Calcaneal Tendons. J. Anat. 2015, 226, 126–142. [Google Scholar] [CrossRef] [Green Version]
- Barboni, B.; Curini, V.; Russo, V.; Mauro, A.; Di Giacinto, O.; Marchisio, M.; Alfonsi, M.; Mattioli, M. Indirect Co-Culture with Tendons or Tenocytes Can Program Amniotic Epithelial Cells towards Stepwise Tenogenic Differentiation. PLoS ONE 2012, 7, e30974. [Google Scholar] [CrossRef] [Green Version]
- Citeroni, M.R.; Mauro, A.; Ciardulli, M.C.; Di Mattia, M.; El Khatib, M.; Russo, V.; Turriani, M.; Santer, M.; Della Porta, G.; Maffulli, N.; et al. Amnion-Derived Teno-Inductive Secretomes: A Novel Approach to Foster Tendon Differentiation and Regeneration in an Ovine Model. Front. Bioeng. Biotechnol. 2021, 9, 649288. [Google Scholar] [CrossRef]
- Janzen, C.; Sen, S.; Lei, M.Y.Y.; de Assumpcao, M.G.; Challis, J.; Chaudhuri, G. The Role of Epithelial to Mesenchymal Transition in Human Amniotic Membrane Rupture. J. Clin. Endocrinol. Metab. 2016, 102, 1261–1269. [Google Scholar] [CrossRef] [Green Version]
- Canciello, A.; Russo, V.; Berardinelli, P.; Bernabò, N.; Muttini, A.; Mattioli, M.; Barboni, B. Progesterone Prevents Epithelial-Mesenchymal Transition of Ovine Amniotic Epithelial Cells and Enhances Their Immunomodulatory Properties. Sci. Rep. 2017, 7, 3761. [Google Scholar] [CrossRef] [Green Version]
- Alcaraz, A.; Mrowiec, A.; Insausti, C.L.; García-Vizcaíno, E.M.; Ruiz-Canada, C.; López-Martínez, M.C.; Moraleda, J.M.; Nicolás, F.J. Autocrine TGF-β Induces Epithelial to Mesenchymal Transition in Human Amniotic Epithelial Cells. Cell Transpl. 2013, 22, 1351–1367. [Google Scholar] [CrossRef]
- Millar, N.L.; Murrell, G.A.C.; Mcinnes, I.B. Inflammatory Mechanisms in Tendinopathy—Towards Translation. Nat. Rev. Rheumatol. 2017, 13, 110–122. [Google Scholar] [CrossRef] [PubMed]
- Millar, N.L.; Silbernagel, K.G.; Thorborg, K.; Kirwan, P.D.; Galatz, L.M.; Abrams, G.D.; Murrell, G.A.C.; McInnes, I.B.; Rodeo, S.A. Tendinopathy. Nat. Rev. Dis. Primers 2021, 7, 1. [Google Scholar] [CrossRef] [PubMed]
- Desai, S.; Jayasuriya, C.T. Implementation of Endogenous and Exogenous Mesenchymal Progenitor Cells for Skeletal Tissue Regeneration and Repair. Bioengineering 2020, 7, 86. [Google Scholar] [CrossRef] [PubMed]
- Sussman, W.I.; Mautner, K.; Malanga, G. The Role of Rehabilitation after Regenerative and Orthobiologic Procedures for the Treatment of Tendinopathy: A Systematic Review. Regen. Med. 2018, 13, 249–263. [Google Scholar] [CrossRef]
- Wong, R.; Alam, N.; McGrouther, A.D.; Wong, J.K.F. Tendon Grafts: Their Natural History, Biology and Future Development. J. Hand Surg. Eur. Vol. 2015, 40, 669–681. [Google Scholar] [CrossRef]
- Van den Boom, N.A.C.; Winters, M.; Haisma, H.J.; Moen, M.H. Efficacy of Stem Cell Therapy for Tendon Disorders: A Systematic Review. Orthop. J. Sports Med. 2020, 8, 232596712091585. [Google Scholar] [CrossRef]
- Barboni, B.; Russo, V.; Gatta, V.; Bernabò, N.; Berardinelli, P.; Mauro, A.; Martelli, A.; Valbonetti, L.; Muttini, A.; Di Giacinto, O.; et al. Therapeutic Potential of HAECs for Early Achilles Tendon Defect Repair through Regeneration. J. Tissue Eng. Regen. Med. 2018, 12, e1594–e1608. [Google Scholar] [CrossRef]
- Barboni, B.; Russo, V.; Curini, V.; Mauro, A.; Martelli, A.; Muttini, A.; Bernabò, N.; Valbonetti, L.; Marchisio, M.; Di Giacinto, O.; et al. Achilles Tendon Regeneration Can Be Improved by Amniotic Epithelial Cell Allotransplantation. Cell Transpl. 2012, 21, 2377–2395. [Google Scholar] [CrossRef] [Green Version]
- Mauro, A.; Russo, V.; Di Marcantonio, L.; Berardinelli, P.; Martelli, A.; Muttini, A.; Mattioli, M.; Barboni, B. M1 and M2 Macrophage Recruitment during Tendon Regeneration Induced by Amniotic Epithelial Cell Allotransplantation in Ovine. Res. Vet. Sci. 2016, 105, 92–102. [Google Scholar] [CrossRef] [Green Version]
- Muttini, A.; Valbonetti, L.; Abate, M.; Colosimo, A.; Curini, V.; Mauro, A.; Berardinelli, P.; Russo, V.; Cocciolone, D.; Marchisio, M.; et al. Ovine Amniotic Epithelial Cells: In Vitro Characterization and Transplantation into Equine Superficial Digital Flexor Tendon Spontaneous Defects. Res. Vet. Sci. 2013, 94, 158–169. [Google Scholar] [CrossRef]
- Muttini, A.; Russo, V.; Rossi, E.; Mattioli, M.; Barboni, B.; Tosi, U.; Maffulli, N.; Valbonetti, L.; Abate, M. Pilot Experimental Study on Amniotic Epithelial Mesenchymal Cell Transplantation in Natural Occurring Tendinopathy in Horses. Ultrasonographic and Histological Comparison. Muscles Ligaments Tendons J. 2015, 5, 5–11. [Google Scholar] [CrossRef] [PubMed]
- Bruns, J.; Kampen, J.; Kahrs, J.; Plitz, W. Achilles Tendon Rupture: Experimental Results on Spontaneous Repair in a Sheep-Model. Knee Surg. Sports Traumatol. Art 2000, 8, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Barone, R. Anatomia Comparata dei Mammiferi Domestici; Edagricole: Bologna, Italy, 1983; Volume 4, ISBN 978-88-506-1455-4. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- El Khatib, M.; Mauro, A.; Di Mattia, M.; Wyrwa, R.; Schweder, M.; Ancora, M.; Lazzaro, F.; Berardinelli, P.; Valbonetti, L.; Di Giacinto, O.; et al. Electrospun PLGA Fiber Diameter and Alignment of Tendon Biomimetic Fleece Potentiate Tenogenic Differentiation and Immunomodulatory Function of Amniotic Epithelial Stem Cells. Cells 2020, 9, 1207. [Google Scholar] [CrossRef]
- Barboni, B.; Russo, V.; Curini, V.; Martelli, A.; Berardinelli, P.; Mauro, A.; Mattioli, M.; Marchisio, M.; Bonassi Signoroni, P.; Parolini, O.; et al. Gestational Stage Affects Amniotic Epithelial Cells Phenotype, Methylation Status, Immunomodulatory and Stemness Properties. Stem Cell Rev. Rep. 2014, 10, 725–741. [Google Scholar] [CrossRef] [Green Version]
- Caruso, M.; Bonassi Signoroni, P.; Zanini, R.; Ressel, L.; Vertua, E.; Bonelli, P.; Dattena, M.; Varoni, M.V.; Wengler, G.; Parolini, O. Feasibility and Potential of in Utero Foetal Membrane-Derived Cell Transplantation. Cell Tissue Bank 2014, 15, 241–249. [Google Scholar] [CrossRef]
- Hoffmann, A. Neotendon Formation Induced by Manipulation of the Smad8 Signalling Pathway in Mesenchymal Stem Cells. J. Clin. Investig. 2006, 116, 940–952. [Google Scholar] [CrossRef] [Green Version]
- Russo, V.; El Khatib, M.; di Marcantonio, L.; Ancora, M.; Wyrwa, R.; Mauro, A.; Walter, T.; Weisser, J.; Citeroni, M.R.; Lazzaro, F.; et al. Tendon Biomimetic Electrospun PLGA Fleeces Induce an Early Epithelial-Mesenchymal Transition and Tenogenic Differentiation on Amniotic Epithelial Stem Cells. Cells 2020, 9, 303. [Google Scholar] [CrossRef] [Green Version]
- El Khatib, M.; Mauro, A.; Wyrwa, R.; Di Mattia, M.; Turriani, M.; Di Giacinto, O.; Kretzschmar, B.; Seemann, T.; Valbonetti, L.; Berardinelli, P.; et al. Fabrication and Plasma Surface Activation of Aligned Electrospun PLGA Fiber Fleeces with Improved Adhesion and Infiltration of Amniotic Epithelial Stem Cells Maintaining Their Teno-Inductive Potential. Molecules 2020, 25, 3176. [Google Scholar] [CrossRef]
- Chen, X.; Yin, Z.; Chen, J.; Shen, W.; Liu, H.; Tang, Q.; Fang, Z.; Lu, L.; Ji, J.; Ouyang, H. Force and Scleraxis Synergistically Promote the Commitment of Human ES Cells Derived MSCs to Tenocytes. Sci. Rep. 2012, 2, 977. [Google Scholar] [CrossRef] [Green Version]
- Docheva, D.; Hunziker, E.B.; Fässler, R.; Brandau, O. Tenomodulin Is Necessary for Tenocyte Proliferation and Tendon Maturation. Mol. Cell. Biol. 2005, 25, 699–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murchison, N.D.; Price, B.A.; Conner, D.A.; Keene, D.R.; Olson, E.N.; Tabin, C.J.; Schweitzer, R. Regulation of Tendon Differentiation by Scleraxis Distinguishes Force-Transmitting Tendons from Muscle-Anchoring Tendons. Development 2007, 134, 2697–2708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shukunami, C.; Takimoto, A.; Nishizaki, Y.; Yoshimoto, Y.; Tanaka, S.; Miura, S.; Watanabe, H.; Sakuma, T.; Yamamoto, T.; Kondoh, G.; et al. Scleraxis Is a Transcriptional Activator That Regulates the Expression of Tenomodulin, a Marker of Mature Tenocytes and Ligamentocytes. Sci. Rep. 2018, 8, 3155. [Google Scholar] [CrossRef] [Green Version]
- Pryce, B.A.; Watson, S.S.; Murchison, N.D.; Staverosky, J.A.; Dünker, N.; Schweitzer, R. Recruitment and Maintenance of Tendon Progenitors by TGFβ Signaling Are Essential for Tendon Formation. Development 2009, 136, 1351–1361. [Google Scholar] [CrossRef] [Green Version]
- Marui, T.; Niyibizi, C.; Georgescu, H.I.; Cao, M.; Kavalkovich, K.W.; Levine, R.E.; Woo, S.L.-Y. Effect of Growth Factors on Matrix Synthesis by Ligament Fibroblasts. J. Orthop. Res. 1997, 15, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Docheva, D.; Majewski, M.; Evans, C.H. Biologics of Tendon Repair. Adv. Drug Deliv. Rev. 2015, 84, 222–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Citeroni, M.R.; Ciardulli, M.C.; Russo, V.; Della Porta, G.; Mauro, A.; El Khatib, M.; Di Mattia, M.; Galesso, D.; Barbera, C.; Forsyth, N.R.; et al. In Vitro Innovation of Tendon Tissue Engineering Strategies. Int. J. Mol. Sci. 2020, 21, 6726. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.L.; Nödl, M.; Galloway, J.L. Bringing Tendon Biology to Heel: Leveraging Mechanisms of Tendon Development, Healing, and Regeneration to Advance Therapeutic Strategies. Dev. Dyn. 2020, 250, 393–413. [Google Scholar] [CrossRef]
- Lin, D.; Alberton, P.; Caceres, M.D.; Volkmer, E.; Schieker, M.; Docheva, D. Tenomodulin Is Essential for Prevention of Adipocyte Accumulation and Fibrovascular Scar Formation during Early Tendon Healing. Cell Death Dis. 2017, 8, e3116. [Google Scholar] [CrossRef] [Green Version]
- Natsuume, T.; Nakamura, N.; Shino, K.; Toritsuka, Y.; Horibe, S.; Ochi, T. Temporal and Spatial Expression of Transforming Growth Factor-? In the Healing Patellar Ligament of the Rat. J. Orthop. Res. 1997, 15, 837–843. [Google Scholar] [CrossRef]
- Jorgensen, H.; Mclellan, S.; Crossan, J.; Curtis, A. Neutralisation of TGFβ or Binding of VLA-4 to Fibronectin Prevents Rat Tendon Adhesion Following Transection. Cytokine 2005, 30, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Kaji, D.A.; Howell, K.L.; Balic, Z.; Hubmacher, D.; Huang, A.H. Tgfβ Signaling Is Required for Tenocyte Recruitment and Functional Neonatal Tendon Regeneration. eLife 2020, 9, e51779. [Google Scholar] [CrossRef] [PubMed]
- Maffulli, N.; Ewen, S.W.B.; Waterston, S.W.; Reaper, J.; Barrass, V. Tenocytes from Ruptured and Tendinopathic Achilles Tendons Produce Greater Quantities of Type III Collagen than Tenocytes from Normal Achilles Tendons: An in Vitro Model of Human Tendon Healing. Am. J. Sports Med. 2000, 28, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.W.; Cardenas, L.; Soslowsky, L.J. Biomechanics of Tendon Injury and Repair. J. Biomech. 2004, 37, 865–877. [Google Scholar] [CrossRef]
- Bian, X.; Liu, T.; Yang, M.; Gu, C.; He, G.; Zhou, M.; Tang, H.; Lu, K.; Lai, F.; Wang, F.; et al. The Absence of Oestrogen Receptor Beta Disturbs Collagen I Type Deposition during Achilles Tendon Healing by Regulating the IRF5-CCL3 Axis. J. Cell. Mol. Med. 2020, 24, 9925–9935. [Google Scholar] [CrossRef] [PubMed]
- Woo, S.L.-Y.; Hildebrand, K.; Watanabe, N.; Fenwick, J.A.; Papageorgiou, C.D.; Wang, J.H.-C. Tissue Engineering of Ligament and Tendon Healing. Clin. Orthop. Relat. Res. 1999, 367, S312–S323. [Google Scholar] [CrossRef]
- Juneja, S.C.; Veillette, C. Defects in Tendon, Ligament, and Enthesis in Response to Genetic Alterations in Key Proteoglycans and Glycoproteins: A Review. Arthritis 2013, 2013, 154812. [Google Scholar] [CrossRef] [Green Version]
- Rigozzi, S.; Müller, R.; Stemmer, A.; Snedeker, J.G. Tendon Glycosaminoglycan Proteoglycan Sidechains Promote Collagen Fibril Sliding—AFM Observations at the Nanoscale. J. Biomech. 2013, 46, 813–818. [Google Scholar] [CrossRef]
- Sharma, P.; Maffulli, N. Biology of Tendon Injury: Healing, Modeling and Remodeling. J. Musculoskelet. Neuronal Interact. 2006, 6, 181–190. [Google Scholar]
- Nourissat, G.; Diop, A.; Maurel, N.; Salvat, C.; Dumont, S.; Pigenet, A.; Gosset, M.; Houard, X.; Berenbaum, F. Mesenchymal Stem Cell Therapy Regenerates the Native Bone-Tendon Junction after Surgical Repair in a Degenerative Rat Model. PLoS ONE 2010, 5, e12248. [Google Scholar] [CrossRef]
- Hou, Y.; Mao, Z.; Wei, X.; Lin, L.; Chen, L.; Wang, H.; Fu, X.; Zhang, J.; Yu, C. Effects of Transforming Growth Factor-Β1 and Vascular Endothelial Growth Factor 165 Gene Transfer on Achilles Tendon Healing. Matrix Biol. 2009, 28, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Manuelpillai, U.; Lourensz, D.; Vaghjiani, V.; Tchongue, J.; Lacey, D.; Tee, J.-Y.; Murthi, P.; Chan, J.; Hodge, A.; Sievert, W. Human Amniotic Epithelial Cell Transplantation Induces Markers of Alternative Macrophage Activation and Reduces Established Hepatic Fibrosis. PLoS ONE 2012, 7, e38631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiorentino, D.F.; Zlotnik, A.; Mosmann, T.R.; Howard, M.; O’Garra, A. IL-10 Inhibits Cytokine Production by Activated Macrophages. J. Immunol. 1991, 147, 3815–3822. [Google Scholar] [PubMed]
- D’Andrea, A.; Aste-Amezaga, M.; Valiante, N.M.; Ma, X.; Kubin, M.; Trinchieri, G. Interleukin 10 (IL-10) Inhibits Human Lymphocyte Interferon 3,-Production by Suppressing Natural Killer Cell Stimulatory Factor/IL-12 Synthesis in Accessory Cells. J. Exp. Med. 1993, 178, 1041–1048. [Google Scholar] [CrossRef]
- Hedrich, C.M.; Bream, J.H. Cell Type-Specific Regulation of IL-10 Expression in Inflammation and Disease. Immunol. Res. 2010, 47, 185–206. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Koike-Soko, C.; Sugimoto, J.; Yoshida, T.; Okabe, M.; Nikaido, T. Human Amnion-Derived Stem Cells Have Immunosuppressive Properties on NK Cells and Monocytes. Cell Transpl. 2015, 24, 2065–2076. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward | Reverse | |
|---|---|---|---|
| EMT | VIM a | 5′-GACCAGCTCACCAACGACA-3′ | 5′-CTCCTCCTGCAACTTCTCCC-3′ |
| CYTO8 b | 5′-CTCAAAGGCCAGAGGGCTTC-3′ | 5′-CTTGGCCTGAGCATCCTTGA-3 | |
| SNAIL a | 5′-GTCGTGGGTGGAGAGCTTTG-3′ | 5′-TGCTGGAAAGTGAGCTCTGG-3′ | |
| TWIST a | 5′-GCCGGAGACCTAGATGTCATTG-3′ | 5′-CCACGCCCTGTTTCTTTGAAT-3′ | |
| TENOGENIC | TNMD a | 5′-TGGTGAAGACCTTCACTTTCC-3′ | 5′-TTAAACCCTCCCCAGCATGC-3′ |
| SCXB a | 5′-AACAGCGTGAACACGGCTTTC-3′ | 5′-TTTCTCTGGTTGCTGAGGCAG-3′ | |
| COL1 a | 5′-CGTGATCTGCGACGAACTTAA-3′ | 5′-GTCCAGGAAGTCCAGGTTGT-3′ | |
| COL3 c,d | 5′-AAGGGCAGGGAACAACTTGAT-3′ | 5′-GTGGGCAAACTGCACAACATT-3′ | |
| TBSH4 c,d | 5′-CCGCAGGTCTTTGACCTTCT-3′ | 5′-CAGGTAACGGAGGATGGCTTT-3′ | |
| IMMUNO | CD86 c,d | 5′-AGAAGGTCCCAAGGACTGGT-3′ | 5′-GCTTGGCACAGGTGACTTTG-3′ |
| CD206 c,d | 5′-GTAGAAGCAGGCTGCCAGAA-3′ | 5′-CTTCTGCCCAGTGTTTGCAC-3′ | |
| IL10 c,d | 5′-CTGTGCCTCTCCCCTAGAGT-3′ | 5′-GCAGCTAGCTCCACAAGGAA-3′ | |
| 1L12 c,d | 5′-ACAAAGGAGGCGAGGTTCTG-3′ | 5′-CTGTGGTCCATGCTGACCTT-3′ | |
| H.K. | GAPDH a | 5′-CCTGCACCACCAACTGCTTG-3′ | 5′-TTGAGCTCAGGGATGACCTTG-3′ |
| Primary Antibody | Dilution | Secondary Antibody | Dilution | |
|---|---|---|---|---|
| EMT | CYTO8 (Abcam, Cambridge, UK) | 1:200 | Anti-mouse Alexa Fluor (Sigma-Aldrich, St. Louis, MO, USA) | 1:500 |
| VIM (Agilent Technologies, Santa Clara, CA, USA) | 1:200 | Anti-mouse Cy3 (Sigma-Aldrich, St. Louis, MO, USA) | 1:750 | |
| TENDON MARKERS | COL1 (Chemicon Int., Billrerica, MA, USA) | 1:200 | Anti-Mouse Alexa Fluor 488 (Invitrogen Ltd., Paisley, UK) | 1:400 |
| COL3 (Chemicon Int., Billrerica, MA, USA) | 1:500 | Anti-Mouse Alexa Fluor 488 (Invitrogen Ltd., Paisley, UK) | 1:400 | |
| TNMD (Abacm, Cambridge, UK) | 1:100 | Anti-Rabbit Alexa Fluor 488 (Invitrogen Ltd., Paisley, UK) | 1:400 | |
| DCN (Invitrogen, Waltham, MA, USA) | 1:100 | Anti-Rabbit Alexa Fluor 488 (Invitrogen Ltd., Paisley, UK) | 1:400 | |
| IMMUNE MARKERS | CD86 (AbD serotec, a Bio-Rad Company, Hercules, CA, USA) | 1:50 | Anti-Mouse Alexa Fluor 488 (Invitrogen Ltd., Paisley, UK) | 1:200 |
| CD206 (RδD Systems, a Bio-techne brand, 614 McKinley Place NE Minneapolis, MN 55,413 USA) | 1:25 | Anti-Goat Alexa Fluor 488 (Molecular Probes, Eugene, OR, USA) | 1:200 | |
| vWF (Dako Cytomation, Denmark) | 1:400 | Alexa Fluor 488 Anti-Rabbit (Invitrogen Ltd., Paisley, UK) | 1:400 |
| Parameter | Score 0 | Score 1 | Score 2 | Score 3 |
|---|---|---|---|---|
| Cellularity expressed as average of cell N° in CTR and treated samples within all analyzed areas/average of cell N° in healthy tendons | Fold change > 4.5 | 4.5 < fold change > 3.1 | 3 < fold change > 1.5 | Fold change < 1.5 |
| Cell nuclei alignment expressed as the ratio between angle distribution of CTR and treated groups vs. healthy tendons along the longitudinal axis of the tendon within area 4 | Ratio > 3 corresponding to an irregular cell distribution and alignment along the longitudinal axis of the tendon | 3 < Ratio > 2.1 corresponding to cells that start to acquire a parallel orientation to the longitudinal axis of the tendon with high variability | 2 < Ratio > 1 corresponding to cells that start to acquire a parallel orientation to the longitudinal axis of the tendon with low variability | Ratio < 1 corresponding to cells that acquire a parallel orientation to the longitudinal axis of the tendon |
| Vascularity | Vascular plexus | Presence of blood vessels that are either scattered or aligned to the longitudinal axis of the tendon for an extension corresponding to max 1 area within the injured tendon | Blood vessels aligned to the longitudinal axis of the tendon for an extension corresponding to 1 up to 4 areas within the injured tendon | Blood vessels oriented along the longitudinal axis of the tendon for an extension corresponding to up to 5 areas within the injured tendon |
| Fiber organization of COL1 within ECM | No COL1 expression and no fibers’ organization within the injured tendon | COL1 expression with fiber formation with an irregular distribution within the injured tendon | COL1 expression with scattered aligned fibers noted only for a small extension within the injured tendon | COL1 expression with aligned fibers noted for a wide extension within the injured tendon |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Russo, V.; Mauro, A.; Peserico, A.; Di Giacinto, O.; Khatib, M.E.; Citeroni, M.R.; Rossi, E.; Canciello, A.; Mazzotti, E.; Barboni, B. Tendon Healing Response Is Dependent on Epithelial–Mesenchymal–Tendon Transition State of Amniotic Epithelial Stem Cells. Biomedicines 2022, 10, 1177. https://doi.org/10.3390/biomedicines10051177
Russo V, Mauro A, Peserico A, Di Giacinto O, Khatib ME, Citeroni MR, Rossi E, Canciello A, Mazzotti E, Barboni B. Tendon Healing Response Is Dependent on Epithelial–Mesenchymal–Tendon Transition State of Amniotic Epithelial Stem Cells. Biomedicines. 2022; 10(5):1177. https://doi.org/10.3390/biomedicines10051177
Chicago/Turabian StyleRusso, Valentina, Annunziata Mauro, Alessia Peserico, Oriana Di Giacinto, Mohammad El Khatib, Maria Rita Citeroni, Emanuela Rossi, Angelo Canciello, Eleonora Mazzotti, and Barbara Barboni. 2022. "Tendon Healing Response Is Dependent on Epithelial–Mesenchymal–Tendon Transition State of Amniotic Epithelial Stem Cells" Biomedicines 10, no. 5: 1177. https://doi.org/10.3390/biomedicines10051177