
Descriptive Analysis of Circulating Antimicrobial Resistance Genes in Vancomycin-Resistant Enterococcus (VRE) during the COVID-19 Pandemic

 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yim, J.; Smith, J.R.; Rybak, M.J. Role of Combination Antimicrobial Therapy for Vancomycin-Resistant Enterococcus faecium Infections: Review of the Current Evidence. J. Hum. Pharmacol. Drug Ther. 2017, 37, 579–592. [Google Scholar] [CrossRef] [PubMed]
- Jahansepas, A.; Aghazadeh, M.; Rezaee, M.A.; Hasani, A.; Sharifi, Y.; Aghazadeh, T.; Mardaneh, J. Occurrence of Enterococcus faecalis and Enterococcus faecium in Various Clinical Infections: Detection of Their Drug Resistance and Virulence Determinants. Microb. Drug Resist. 2018, 24, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Monticelli, J.; Knezevich, A.; Luzzati, R.; di Bella, S. Clinical management of non-faecium non-faecalis vancomycin-resistant enterococci infection. Focus on Enterococcus gallinarum and Enterococcus casseliflavus/flavescens. J. Infect. Chemother. 2018, 24, 237–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorrie, C.; Higgs, C.; Carter, G.; Stinear, T.P.; Howden, B. Genomics of vancomycin-resistant Enterococcus faecium. Microb. Genom. 2019, 5, e000283. [Google Scholar] [CrossRef] [PubMed]
- Courvalin, P. Vancomycin Resistance in Gram-Positive Cocci. Clin. Infect. Dis. 2006, 42, S25–S34. Available online: http://cid.oxfordjournals.org/ (accessed on 4 February 2022). [CrossRef] [PubMed]
- Coombs, G.W.; Pearson, J.C.; Daly, D.A.; Le, T.T.; Robinson, J.O.; Gottlieb, T.; Howden, B.P.; Johnson, P.D.R.; Bennett, C.M.; Stinear, T.P.; et al. Australian Enterococcal Sepsis Outcome Programme annual report, 2013. Commun. Dis. Intell. Q. Rep. 2014, 38, E320–E326. [Google Scholar] [PubMed]
- Bender, J.K.; Kalmbach, A.; Fleige, C.; Klare, I.; Fuchs, S.; Werner, G. Population structure and acquisition of the vanB resistance determinant in German clinical isolates of Enterococcus faecium ST192. Sci. Rep. 2016, 6, 21847. [Google Scholar] [CrossRef]
- Ahmed, M.O.; Baptiste, K.E. Vancomycin-Resistant Enterococci: A Review of Antimicrobial Resistance Mechanisms and Perspectives of Human and Animal Health. Microb. Drug Resist. 2018, 24, 590–606. [Google Scholar] [CrossRef] [Green Version]
- Olearo, F.; Both, A.; Campos, C.B.; Hilgarth, H.; Klupp, E.-M.; Hansen, J.L.; Maurer, F.P.; Christner, M.; Aepfelbacher, M.; Rohde, H. Emergence of linezolid-resistance in vancomycin-resistant Enterococcus faecium ST117 associated with increased linezolid-consumption. Int. J. Med. Microbiol. 2021, 311, 151477. [Google Scholar] [CrossRef]
- Torres, C.; Alonso, C.A.; Ruiz-Ripa, L.; León-Sampedro, R.; del Campo, R.; Coque, T.M. Antimicrobial Resistance in Enterococcus spp. of animal origin. Microbiol. Spectr. 2018, 6, 24. [Google Scholar] [CrossRef]
- Bender, J.K.; Cattoir, V.; Hegstad, K.; Sadowy, E.; Coque, T.M.; Westh, H.; Hammerum, A.M.; Schaffer, K.; Burns, K.; Murchan, S.; et al. Update on prevalence and mechanisms of resistance to linezolid, tigecycline and daptomycin in enterococci in Europe: Towards a common nomenclature. Drug Resist. Updat. 2018, 40, 25–39. [Google Scholar] [CrossRef] [PubMed]
- Ayobami, O.; Willrich, N.; Reuss, A.; Eckmanns, T.; Markwart, R. The ongoing challenge of vancomycin-resistant Enterococcus faecium and Enterococcus faecalis in Europe: An epidemiological analysis of bloodstream infections. Emerg. Microbes Infect. 2020, 9, 1180–1193. [Google Scholar] [CrossRef] [PubMed]
- Yam, E.L.Y. COVID-19 will further exacerbate global antimicrobial resistance. J. Travel Med. 2020, 27, taaa098. [Google Scholar] [CrossRef]
- Crăciunaş, C.; Butiuc-Keul, A.; Flonta, M.; Brad, A.; Sigarteu, M. Applications of molecular techniques to the study of Pseudomonas aeruginosa clinical isolate in Cluj-Napoca, Romania. Ann. Oradea Univ. Biol. Fascicle 2010, 243–247. [Google Scholar]
- Butiuc-Keul, A.; Carpa, R.; Podar, D.; Szekeres, E.; Muntean, V.; Iordache, D.; Farkas, A. Antibiotic Resistance in Pseudomonas spp. Through the Urban Water Cycle. Curr. Microbiol. 2021, 78, 1227–1237. [Google Scholar] [CrossRef] [PubMed]
- Rawson, T.M.; Moore, L.S.; Zhu, N.; Ranganathan, N.; Skolimowska, K.; Gilchrist, M.; Satta, G.; Cooke, G.; Holmes, A. Bacterial and fungal co-infection in individuals with coronavirus: A rapid review to support COVID-19 antimicrobial prescribing. Clin. Infect. Dis. 2020, 71, 2459–2468. [Google Scholar]
- Rawson, T.M.; Moore, L.; Castro-Sanchez, E.; Charani, E.; Davies, F.; Satta, G.; Ellington, M.J.; Holmes, A.H. COVID-19 and the potential long-term impact on antimicrobial resistance. J. Antimicrob. Chemother. 2020, 75, 1681–1684. [Google Scholar] [CrossRef]
- Knight, G.M.; Glover, R.E.; McQuaid, C.F.; Olaru, I.D.; Gallandat, K.; Leclerc, Q.J.; Fuller, N.M.; Willcocks, S.J.; Hasan, R.; van Kleef, E.; et al. Antimicrobial resistance and COVID-19: Intersections and implications. eLife 2021, 10, e64139. [Google Scholar] [CrossRef]
- Ukuhor, H.O. The interrelationships between antimicrobial resistance, COVID-19, past, and future pandemics. J. Infect. Public Health 2020, 14, 53–60. [Google Scholar] [CrossRef]
- Hughes, S.; Troise, O.; Donaldson, H.; Mughal, N.; Moore, L.S.P. Bacterial and fungal coinfection among hospitalized patients with COVID-19: A retrospective cohort study in a UK secondary-care setting. Clin. Microbiol. Infect. 2020, 26, 1395–1399. [Google Scholar] [CrossRef]
- Toc, D.A.; Costache, C.; Botan, A.; Mihaila, R.M.; Colosi, I.A.; Buksa, S.B.; Chiorescu, R.M. Mixed Etiology COVID-19 Associated Pulmonary Aspergillosis (CAPA)—A Case Report and Brief Review of the Literature. J. Fungi 2021, 7, 877. [Google Scholar] [CrossRef] [PubMed]
- Giacobbe, D.R.; Battaglini, D.; Ball, L.; Brunetti, I.; Bruzzone, B.; Codda, G.; de Maria, A.; Dentone, C.; di Biagio, A.; Icardi, G.; et al. Bloodstream infections in critically ill patients with COVID-19. Eur. J. Clin. Investig. 2020, 10, e13319. [Google Scholar] [CrossRef] [PubMed]
- Raza, T.; Ullah, S.R.; Mehmood, K.; Andleeb, S. Vancomycin resistant Enterococci: A brief review. J. Pak. Med. Assoc. 2018, 68, 768–772. [Google Scholar] [PubMed]
- Miller, W.R.; Murray, B.E.; Rice, L.B.; Arias, C.A. Resistance in Vancomycin-Resistant Enterococci. Infect. Dis. Clin. N. Am. 2020, 34, 751–771. [Google Scholar] [CrossRef]
- Ducu, R.; Gheorghe, I.; Chifiriuc, M.C.; Mihăescu, G.; Sârbu, I. Prevalence of vancomycin resistance phenotypes among Enterococcus species isolated from clinical samples in a Romanian hospital. Biointerface Res. Appl. Chem. 2019, 9, 4699–4704. [Google Scholar] [CrossRef]
- Mirzaei, B.; Babaei, R.; Asiabar, A.P.D.; Bameri, Z. Detection of both vanA & vanB genes in vanA phenotypes of Enterococci by Taq Man RT-PCR. Braz. J. Microbiol. 2015, 46, 161–165. [Google Scholar] [CrossRef]
- Papagiannitsis, C.; Malli, E.; Florou, Z.; Medvecky, M.; Sarrou, S.; Hrabak, J.; Petinaki, E. First description in Europe of the emergence of Enterococcus faecium ST117 carrying both vanA and vanB genes, isolated in Greece. J. Glob. Antimicrob. Resist. 2017, 11, 68–70. [Google Scholar] [CrossRef]
- Laverde Gomez, J.A.; Hendrickx, A.P.A.; Willems, R.J.; Top, J.; Sava, I.; Huebner, J.; Witte, W.; Werner, G. Intra- and interspecies genomic transfer of the Enterococcus faecalis pathogenicity Island. PLoS ONE 2011, 6, e16720. [Google Scholar]
- Manson, J.M.; Hancock, L.E.; Gilmore, M.S. Mechanism of chromosomal transfer of Enterococcus faecalis pathogenicity island, capsule, antimicrobial resistance, and other traits. Proc. Natl. Acad. Sci. USA 2010, 107, 12269–12274. [Google Scholar] [CrossRef] [Green Version]
- Palmer, K.L.; Kos, V.N.; Gilmore, M.S. Horizontal gene transfer and the genomics of enterococcal antibiotic resistance. Curr. Opin. Microbiol. 2010, 13, 632–639. [Google Scholar] [CrossRef] [Green Version]
- Dos Santos, L.D.R.; Furlan, J.P.R.; Gallo, I.F.L.; Ramos, M.S.; Savazzi, E.A.; Stehling, E.G. Occurrence of multidrug-resistant Enterococcus faecium isolated from environmental samples. Lett. Appl. Microbiol. 2021, 73, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Molechan, C.; Amoako, D.G.; Abia, A.L.K.; Somboro, A.M.; Bester, L.A.; Essack, S.Y. Molecular epidemiology of antibiotic-resistant Enterococcus spp. from the farm-to-fork continuum in intensive poultry production in KwaZulu-Natal, South Africa. Sci. Total Environ. 2019, 692, 868–878. [Google Scholar] [CrossRef] [PubMed]
- García-Solache, M.; Rice, L.B. The Enterococcus: A Model of Adaptability to Its Environment. Clin. Microbiol. Rev. 2019, 32, e00058-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kresken, M.; Becker, K.; Seifert, H.; Leitner, E.; Körber-Irrgang, B.; Von Eiff, C.; Löschmann, P.-A. Study group. Resistance trends and in vitro activity of tigecycline and 17 other antimicrobial agents against Gram-positive and Gram-negative organisms, including multidrug-resistant pathogens, in Germany. Eur. J. Clin. Microbiol. 2011, 30, 1095–1103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiedler, S.; Bender, J.K.; Klare, I.; Halbedel, S.; Grohmann, E.; Szewzyk, U.; Werner, G. Tigecycline resistance in clinical isolates of Enterococcus faecium is mediated by an upregulation of plasmid-encoded tetracycline determinants tet(L) and tet(M). J. Antimicrob. Chemother. 2016, 71, 871–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, T.-W.; Hung, W.-C.; Tsai, J.-C.; Lin, Y.-T.; Lee, H.; Hsueh, P.-R.; Lee, T.-F.; Teng, L.-J. Novel Structure of Enterococcus faecium-Originated ermB -Positive Tn 1546 -Like Element in Staphylococcus aureus. Antimicrob. Agents Chemother. 2016, 60, 6108–6114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portillo, A.; Ruiz-Larrea, F.; Zarazaga, M.; Alonso, A.; Martinez, J.L.; Torres, C. Macrolide Resistance Genes in Enterococcus spp. Antimicrob. Agents Chemother. 2000, 44, 967–971. [Google Scholar] [CrossRef] [Green Version]
- Adesida, S.A.; Ezenta, C.C.; Adagbada, A.O.; Aladesokan, A.A.; Coker, A.O. Carriage of Multidrug Resistant Enterococcus Faecium and Enterococcus Faecalis Among Apparently Healthy Humans. Afr. J. Infect. Dis. 2017, 11, 83–89. [Google Scholar] [CrossRef] [Green Version]
- Li, X.Z.; Nikaido, H. Efflux-mediated drug resistance in bacteria: An update. Drugs 2009, 69, 1555–1623. [Google Scholar] [CrossRef]
- Dziri, R.; Lozano, C.; Ben Said, L.; Bellaaj, R.; Boudabous, A.; Ben Slama, K.; Torres, C.; Klibi, N. Multidrug-resistant enterococci in the hospital environment: Detection of novel vancomycin-resistant E. faecium clone ST910. J. Infect. Dev. Ctries. 2016, 10, 799–806. [Google Scholar] [CrossRef] [Green Version]
- López, M.; Sáenz, Y.; Rojo-Bezares, B.; Martínez, S.; del Campo, R.; Ruiz-Larrea, F.; Zarazaga, M.; Torres, C. Detection of vanA and vanB2-containing enterococci from food samples in Spain, including Enterococcus faecium strains of CC17 and the new singleton ST425. Int. J. Food Microbiol. 2009, 133, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Barreto, Â.; Guimarães, B.; Radhouani, H.; Araújo, C.; Gonçalves, A.; Gaspar, E.; Rodrigues, J.; Igrejas, G.; Poeta, P. Detection of antibiotic resistant E. coli and Enterococcus spp. in stool of healthy growing children in Portugal. J. Basic Microbiol. 2009, 49, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-H.; Lin, S.-Y.; Lin, Y.-T.; Tseng, S.-P.; Chang, C.-C.; Yu, S.-Y.; Hung, W.-W.; Jao, Y.-T.; Lin, C.-Y.; Chen, Y.-H. Emergence of aac(6′)-Ie-aph(2′′)-Ia-positive enterococci with non-high-level gentamicin resistance mediated by IS1216V: Adaptation to decreased aminoglycoside usage in Taiwan. J. Antimicrob. Chemother. 2021, 76, 1689–1697. [Google Scholar] [CrossRef] [PubMed]
- Ounissi, H.; Derlot, E.; Carlier, C.; Courvalin, P. Gene homogeneity for aminoglycoside-modifying enzymes in gram-positive cocci. Antimicrob. Agents Chemother. 1990, 34, 2164–2168. [Google Scholar] [CrossRef] [Green Version]
- Chow, J.W.; Zervos, M.J.; Lerner, S.A.; Thal, L.A.; Donabedian, S.M.; Jaworski, D.D.; Tsai, S.; Shaw, K.J.; Clewell, D.B. A novel gentamicin resistance gene in Enterococcus. Antimicrob. Agents Chemother. 1997, 41, 511–514. [Google Scholar] [CrossRef] [Green Version]
- Moellering, R.C.; Weinberg, A.N. Effect of various antibiotics on the uptake of 4C-labeles streptomycin by Enterococci. J. Clin. Investig. 1971, 50, 2580–2584. [Google Scholar] [CrossRef] [Green Version]
- Geraci, J.E.; Martin, W.J. Antibiotic Therapy of Bacterial Endocarditis-Pathologic and Therapeutic Consideration of 33 Cases. Circulation 1954, 10, 173–194. Available online: http://ahajournals.org (accessed on 4 February 2022). [CrossRef] [Green Version]
- Rehman, M.U.; Zhang, H.; Huang, S.; Iqbal, M.K.; Mehmood, K.; Luo, H.; Li, J. Characteristics of Integrons and Associated Gene Cassettes in Antibiotic-Resistant Escherichia coli Isolated from Free-Ranging Food Animals in China. J. Food Sci. 2017, 82, 1902–1907. [Google Scholar] [CrossRef]
- Wu, S.; Dalsgaard, A.; Hammerum, A.M.; Porsbo, L.J.; Jensen, L.B. Prevalence and characterization of plasmids carrying sulfonamide resistance genes among Escherichia coli from pigs, pig carcasses and human. Acta Veter. Scand. 2010, 52, 47. [Google Scholar] [CrossRef] [Green Version]
- Ben, W.; Wang, J.; Pan, X.; Qiang, Z. Dissemination of antibiotic resistance genes and their potential removal by on-farm treatment processes in nine swine feedlots in Shandong Province, China. Chemosphere 2017, 167, 262–268. [Google Scholar] [CrossRef]
- Changkaew, K.; Utrarachkij, F.; Siripanichgon, K.; Nakajima, C.; Suthienkul, O.; Suzuki, Y. Characterization of Antibiotic Resistance in Escherichia coli Isolated from Shrimps and Their Environment. J. Food Prot. 2014, 77, 1394–1401. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Cheng, H.; Liang, Y.; Yu, S.; Yu, T.; Fang, J.; Zhu, C. Diverse Mobile Genetic Elements and Conjugal Transferability of Sulfonamide Resistance Genes (sul1, sul2, and sul3) in Escherichia coli Isolates from Penaeus vannamei and Pork from Large Markets in Zhejiang, China. Front. Microbiol. 2019, 10, 1787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wailan, A.M.; Paterson, D.L. The spread and acquisition of NDM-1: A multifactorial problem. Expert Rev. Anti-Infect. Ther. 2014, 12, 91–115. [Google Scholar] [CrossRef]
- Yong, D.; Toleman, M.A.; Giske, C.G.; Cho, H.S.; Sundman, K.; Lee, K.; Walsh, T.R. Characterization of a new metallo-β-lactamase gene, bla NDM-1, and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob. Agents Chemother. 2009, 53, 5046–5054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, T.R.; Weeks, J.; Livermore, D.M.; Toleman, M.A. Dissemination of NDM-1 positive bacteria in the New Delhi environment and its implications for human health: An environmental point prevalence study. Lancet Infect. Dis. 2011, 11, 355–362. [Google Scholar] [CrossRef]
- Arbune, M.; Gurau, G.; Niculet, E.; Iancu, A.V.; Lupasteanu, G.; Fotea, S.; Vasile, M.C.; Tatu, A.L. Prevalence of Antibiotic Resistance of ESKAPE Pathogens Over Five Years in an Infectious Diseases Hospital from South-East of Romania. Infect. Drug Resist. 2021, 14, 2369–2378. [Google Scholar] [CrossRef]
- Lazăr, V.; Gheorghe, I.; Curutiu, C.; Savin, I.; Marinescu, F.; Cristea, V.C.; Dobre, D.; Popa, G.L.; Chifiriuc, M.C.; Popa, M.I. Antibiotic resistance profiles in cultivable microbiota isolated from some romanian natural fishery lakes included in Natura 2000 network. BMC Veter. Res. 2021, 17, 52. [Google Scholar] [CrossRef]
- Almahdawy, O.T.; Pricop, R.; Sadik, O.; Najee, H.; Pircalabioru, G.G.; Marutescu, L.; Barbu, I.C.; Banu, O.; Cristea, V.; Grigore, R.; et al. Description of vancomycin resistance genes in Enterococcus sp. clinical strains isolated from Bucharest, Romania. Rom. Biotechnol. Lett. 2019, 24, 395–399. [Google Scholar] [CrossRef]
- Toc, D.A.; Pandrea, S.L.; Botan, A.; Mihaila, R.M.; Costache, C.A.; Colosi, I.A.; Junie, L.M. Enterococcus raffinosus, Enterococcus durans and Enterococcus avium Isolated from a Tertiary Care Hospital in Romania—Retrospective Study and Brief Review. Biology 2022, 11, 598. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Pairs | Amplicon (bp) | |
|---|---|---|---|
| Forward | Reverse | ||
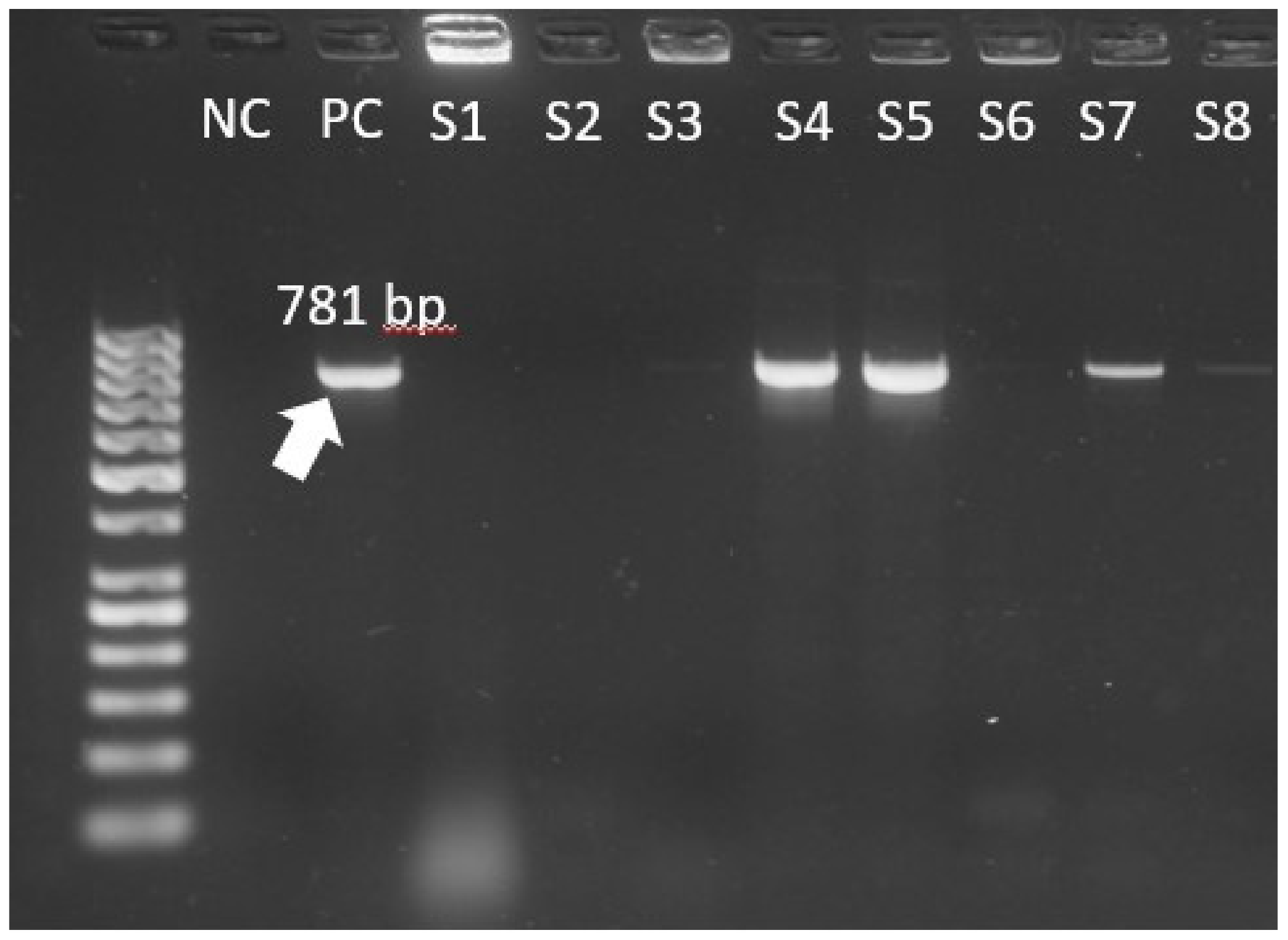
| vanA | GCTATTCAGCTGTACT | CAGCGGCCATCATACGG | 781 |
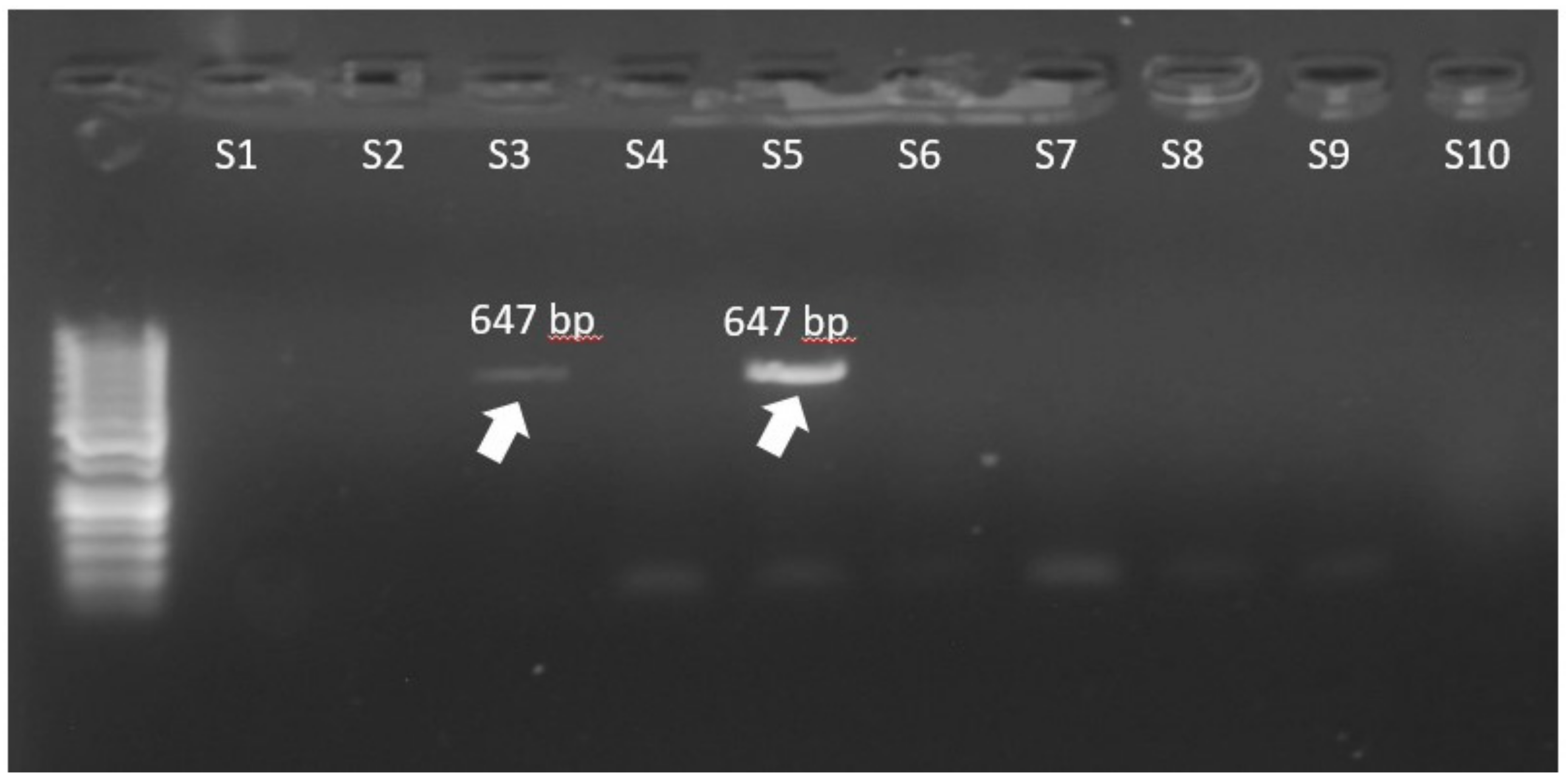
| vanB | CGCCATATTCTCCCCGGATAG | AAGCCCTCTGCATCCAAGCAC | 647 |
| tet(M) | CCGTCTGAACTTTGCGGAAA | CAACGGAAGCGGTGATACAG | 627 |
| tet(L) | TATTCAAGGGGCTGGTGCAG | CGGCAGTACTTAGCTGGTGA | 545 |
| ermB | GAAAAGGTACTCAACCAAATA | AGTAACGGTACTTAAATTGTTTAC | 639 |
| msrA | AGGGAAAGGTCATTTTACTGC | CCCTACCTATAACTAAACATT | 343 |
| mefA | CATCGACGTATTGGGTGCTG | CCGAAAGCCCCATTATTGCA | 516 |
| aac(6′)-Im | GGCTGACAGATGACCGTGTTCTTG | GTAGATATTGGCATACTACTCTGC | 482 |
| aph(2)-Ib | CTGAACACAGCAGCGACTAC | TTGTAATCGCCATGCACCAG | 646 |
| ant(4′)-Ia | GTCAAAAACTGCTAACACAAG | AATAATACTGCTAACGATAAT | 135 |
| sul1 | AGGCATGATCTAACCCTCGG | GGCCGATGAGATCAGACGTA | 665 |
| sul2 | GACAGTTATCAACCCGCGAC | GAAACAGACAGAAGCACCGG | 380 |
| sul3 | GTGGGCGTTGTGGAAGAAAT | AAAAGAAGCCCATACCCGGA | 370 |
| NDM-1 | GGTTTGGCGATCTGGTTTTC | CGGAATGGCTCATCACGATC | 621 |
| Gene | Initial Denaturation | Steps (30 Cycles) | Final Elongation |
|---|---|---|---|
| vanA, ermB | 94 °C for 4 min | Denaturation at 94 °C for 1 min Annealing at 51 °C for 45 s Elongation at 72 °C for 45 s | 72 °C for 8 min |
| ant(4′)-la, msrA | Denaturation at 94 °C for 1 min Annealing at 53 °C for 45 s Elongation at 72 °C for 45 s | ||
| aph(2)-lb, mefA, TetM, sul3 | Denaturation at 94 °C for 1 min Annealing at 55 °C for 45 s Elongation at 72 °C for 45 s | ||
| TetL, sul1, sul2 | Denaturation at 94 °C for 1 min Annealing at 57 °C for 45 s Elongation at 72 °C for 45 s | ||
| aac(6′)–Im, vanB | Denaturation at 94 °C for 1 min Annealing at 61 °C for 45 s Elongation at 72 °C for 45 s |
| Enterococcus Faecalis (n = 11) | Enterococcus Faecium (n = 68) | p | |
|---|---|---|---|
| van A | 9 (81.8%) | 57 (83.8%) | 1.000 |
| van B | 0 (0%) | 8 (11.8%) | 0.591 |
| tet(M) | 5 (45.5%) | 38 (55.9%) | 0.529 |
| tet(L) | 1 (9.1%) | 5 (7.4%) | 1.000 |
| ermB | 9 (81.8%) | 66 (97.1%) | 0.091 |
| msrA | 0 (0%) | 2 (2.9%) | 1.000 |
| mefA | 0 (0%) | 1 (1.5%) | 1.000 |
| aac(6′)-Im | / | / | N/A |
| aph(2)-Ib | / | / | N/A |
| ant(4′)-Ia | 2 (18.2%) | 20 (29.41%) | 0.718 |
| sul1 | / | / | N/A |
| sul2 | / | / | N/A |
| sul3 | / | / | N/A |
| NDM1 | / | / | N/A |
| Frequency | Percentage | |
|---|---|---|
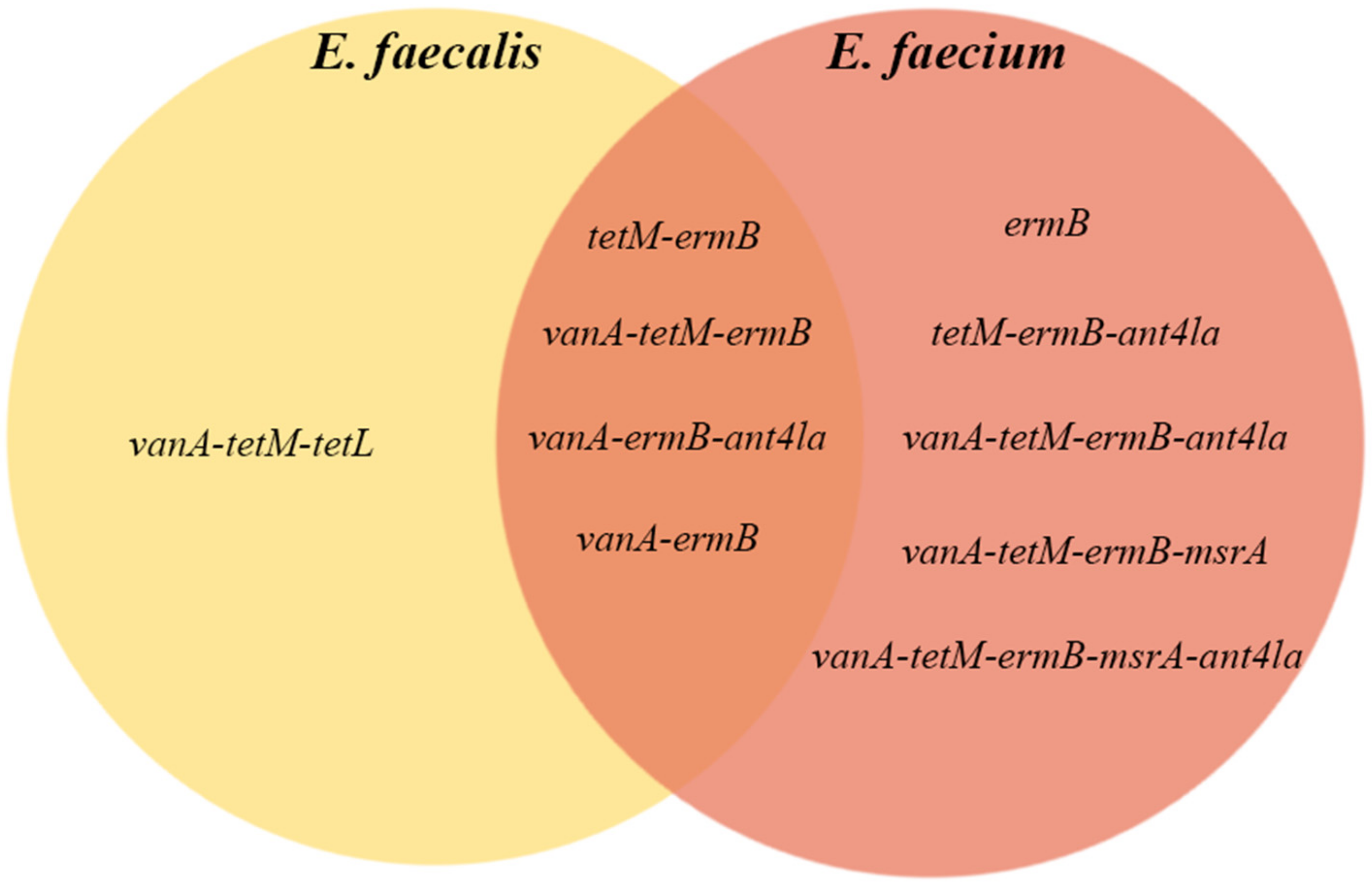
| Resistance gene | 1 | 9.1% |
| tetM-ermB | 1 | 9.1% |
| vanA-tetM-tetL | 1 | 9.1% |
| vanA-ermB-ant4la | 2 | 18.2% |
| vanA-ermB | 3 | 27.3% |
| vanA-tetM-ermB | 3 | 27.3% |
| Total | 11 | 100.0% |
| Frequency | Percentage | |
|---|---|---|
| Resistance gene | 1 | 1.5% |
| tetM-ermB | 1 | 1.5% |
| tetM-ermB-ant4la | 1 | 1.5% |
| vanA-tetM-ermB-msrA | 1 | 1.5% |
| vanA-tetM-ermB-msrA-ant4la | 1 | 1.5% |
| vanA-tetM-tetL-ermB-ant4la | 1 | 1.5% |
| vanA-tetM-tetL-ermB-mefA | 1 | 1.5% |
| vanA-vanB-ermB | 1 | 1.5% |
| vanA-vanB-tetM-ant4la | 1 | 1.5% |
| ermB | 2 | 2.9% |
| vanB-tetM-ermB | 2 | 2.9% |
| vanA-tetM-tetL-ermB | 3 | 4.4% |
| vanB-ermB | 4 | 5.9% |
| vanA-tetM-ermB-ant4la | 6 | 8.8% |
| vanA-ermB-ant4la | 10 | 14.7% |
| vanA-ermB | 12 | 17.6% |
| vanA-tetM-ermB | 20 | 29.4% |
| Total | 68 | 100.0% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Toc, D.A.; Butiuc-Keul, A.L.; Iordache, D.; Botan, A.; Mihaila, R.M.; Costache, C.A.; Colosi, I.A.; Chiorean, C.; Neagoe, D.S.; Gheorghiu, L.; et al. Descriptive Analysis of Circulating Antimicrobial Resistance Genes in Vancomycin-Resistant Enterococcus (VRE) during the COVID-19 Pandemic. Biomedicines 2022, 10, 1122. https://doi.org/10.3390/biomedicines10051122
Toc DA, Butiuc-Keul AL, Iordache D, Botan A, Mihaila RM, Costache CA, Colosi IA, Chiorean C, Neagoe DS, Gheorghiu L, et al. Descriptive Analysis of Circulating Antimicrobial Resistance Genes in Vancomycin-Resistant Enterococcus (VRE) during the COVID-19 Pandemic. Biomedicines. 2022; 10(5):1122. https://doi.org/10.3390/biomedicines10051122
Chicago/Turabian StyleToc, Dan Alexandru, Anca Livia Butiuc-Keul, Dumitrana Iordache, Alexandru Botan, Razvan Marian Mihaila, Carmen Anca Costache, Ioana Alina Colosi, Claudia Chiorean, Dan Stefan Neagoe, Liana Gheorghiu, and et al. 2022. "Descriptive Analysis of Circulating Antimicrobial Resistance Genes in Vancomycin-Resistant Enterococcus (VRE) during the COVID-19 Pandemic" Biomedicines 10, no. 5: 1122. https://doi.org/10.3390/biomedicines10051122