
Restricted Activation of the NF-κB Pathway in Individuals with Latent Tuberculosis Infection after HIF-1α Blockade

 , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statements
2.2. Chemicals and Reagents
2.3. Study Population
2.4. Tuberculin Skin Test
2.5. IFN-γ Release Assay
2.6. Isolation of Peripheral Blood Mononuclear Cells
2.7. Gene Expression Analysis
- GAPDH:
- Forward: 5′-ACCCACTCCTCCACCTTTGA-3′,
- Reverse: 5′-CTTCTACTGGTTCAGCAGCCATCT-3′;
- A20:
- Forward: 5′-CGTCCAGGTTCCAGAACACCATTC-3′,
- Reverse: 5′-TGCGCTGGCTCGATCTCAGTTG-3′;
- IL15:
- Forward: 5′-GGA ATGTAACAGAATCTGGATG-3’,
- Reverse: 5′-GTT ATGTCTAAGCAGCAGAG-3′;
- IL18:
- Forward: 5′-ATCGCTTCCTCTCGCAACAA-3′,
- Reverse: 5′-CTTCTACTGGTTCAGCAGCCATCT-3′;
- HIF1A:
- Forward: 5′-CATAAAGTCTGCAACATGGAAGGT-3′,
- Reverse: 5′-ATTTGATGGGTGAGGAATGGGTT-3′;
- TNFA:
- Forward: 5′-CACACTCAGATCATCTTCTCAA-3′,
- Reverse: 5′-TTGAAGAGAACCTGGGAGTAG-3′;
- IL6:
- Forward: 5′-ATTACACATGTTCTCTGGGAAG-3′,
- Reverse: 5′-TTTTACCTCTTGGTTGAAGATATG-3′.
2.8. 25-hydroxyvitamin D3 (25(OH)D3) Levels
2.9. Cell Culture and Treatment Conditions
2.10. ELISA for NF-κB and TNF-α Production
2.11. Statistical Analysis
3. Results
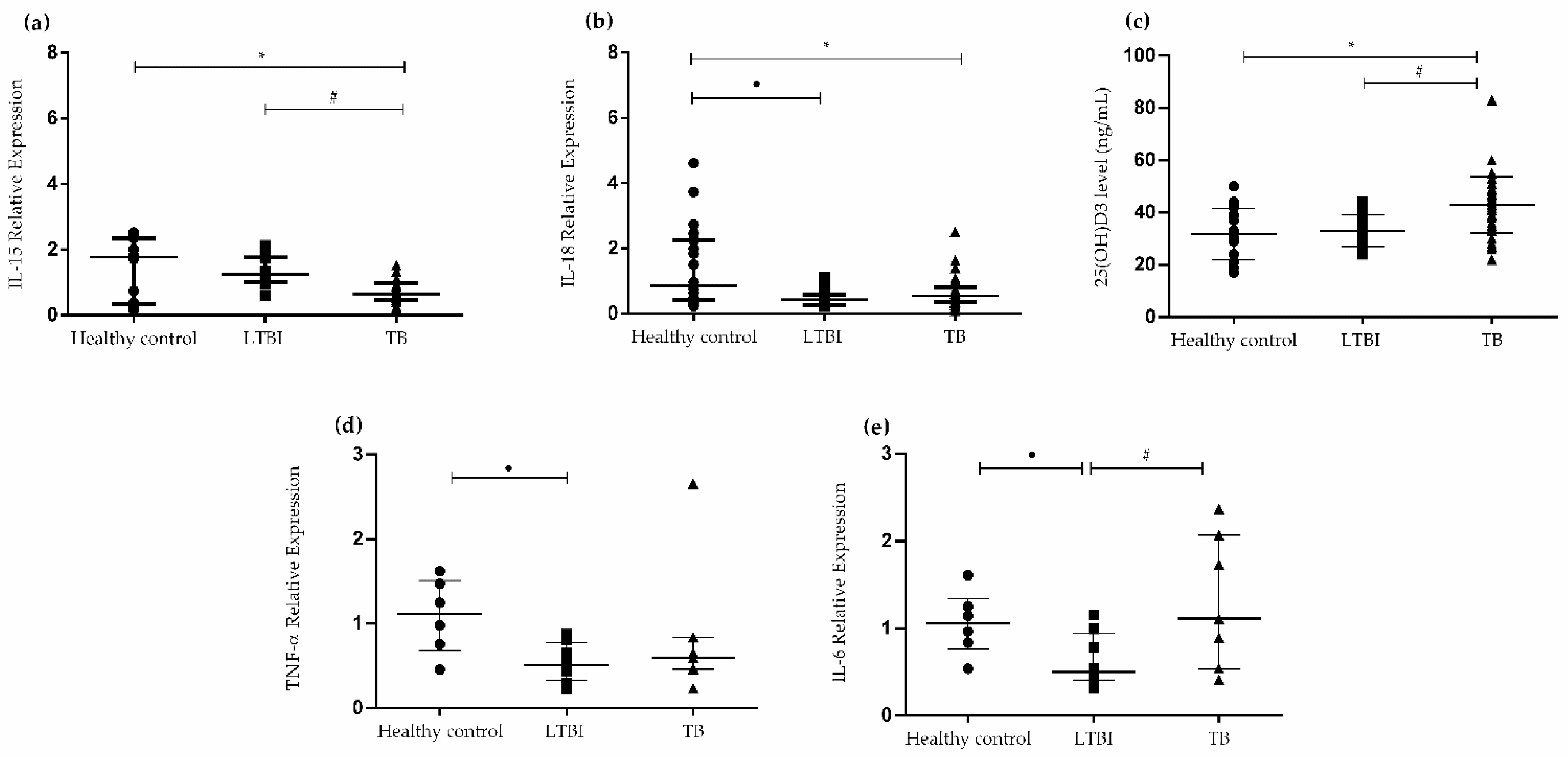
3.1. Cytokine Gene Expression Levels in PBMCs and Levels of 25(OH)D3
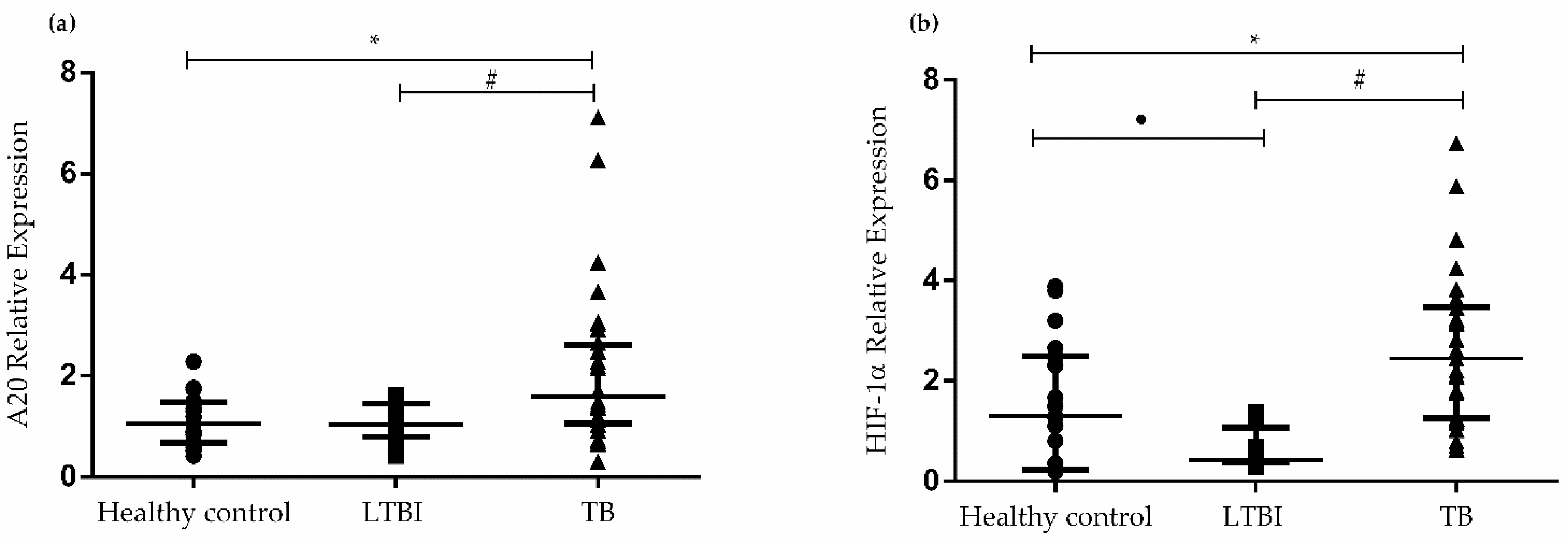
3.2. Gene Expression of A20 and HIF1A in PBMCs
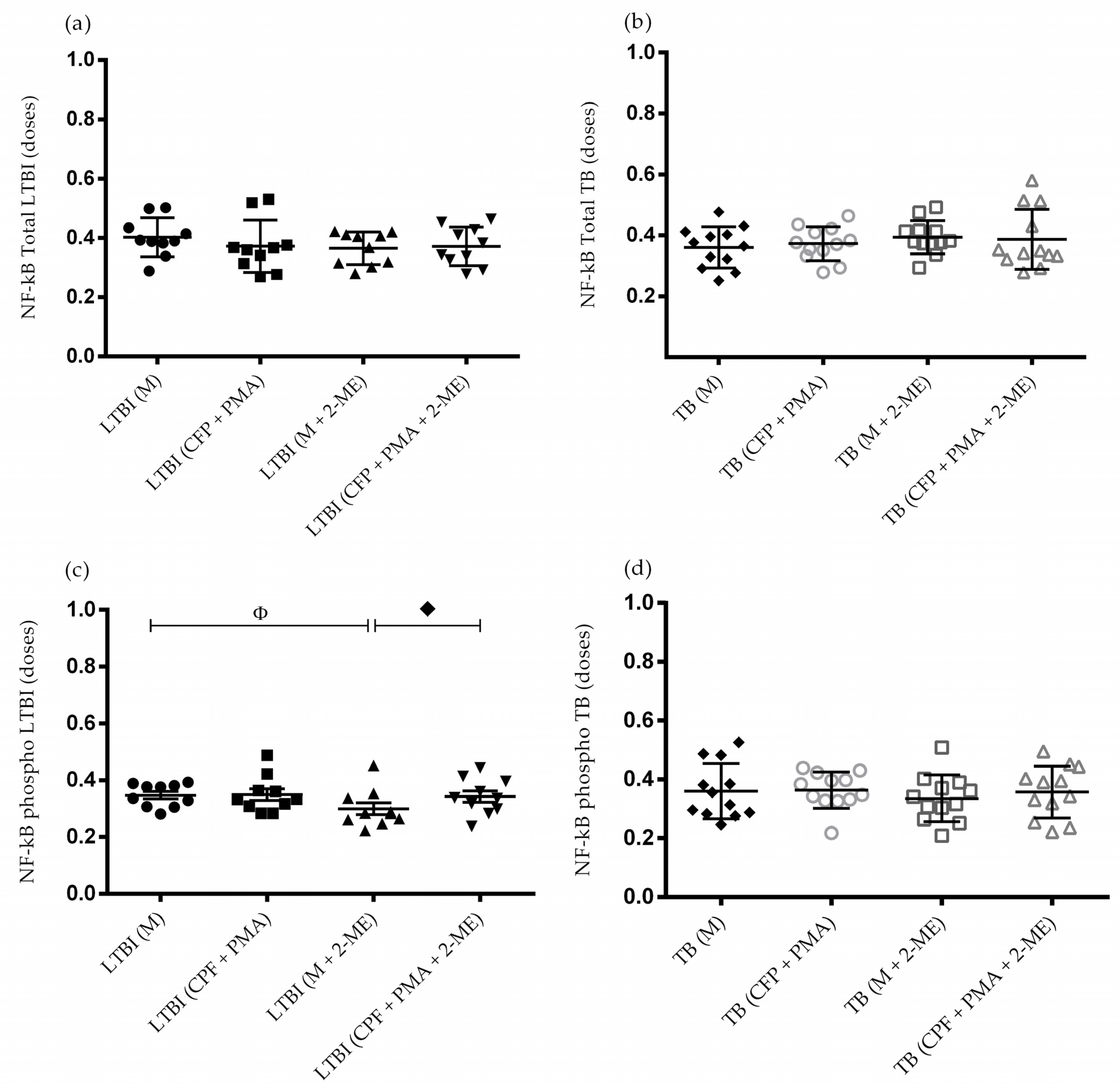
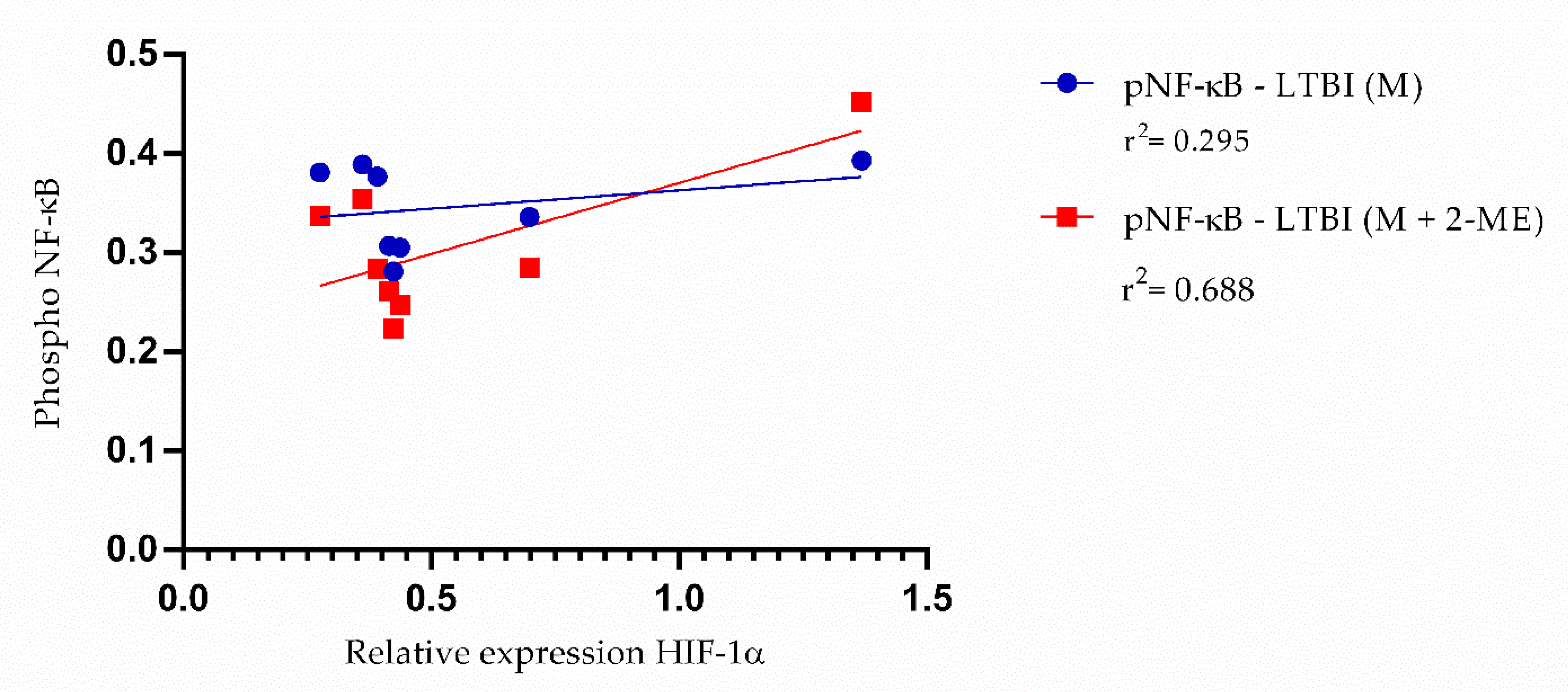
3.3. Activation of the NF-κB Pathway in the Absence of HIF-1α
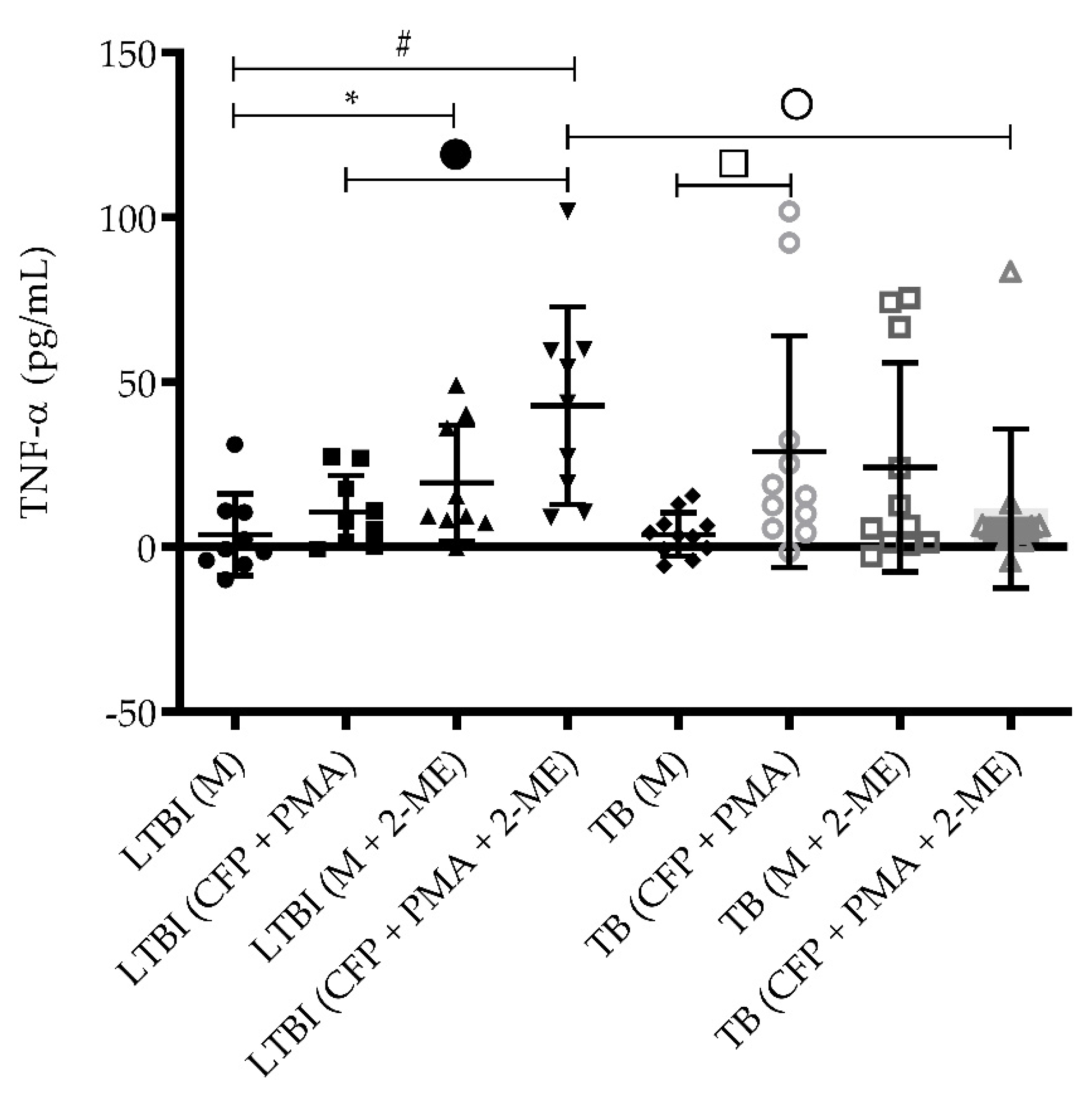
3.4. TNF-α Levels in the Absence of HIF-1α
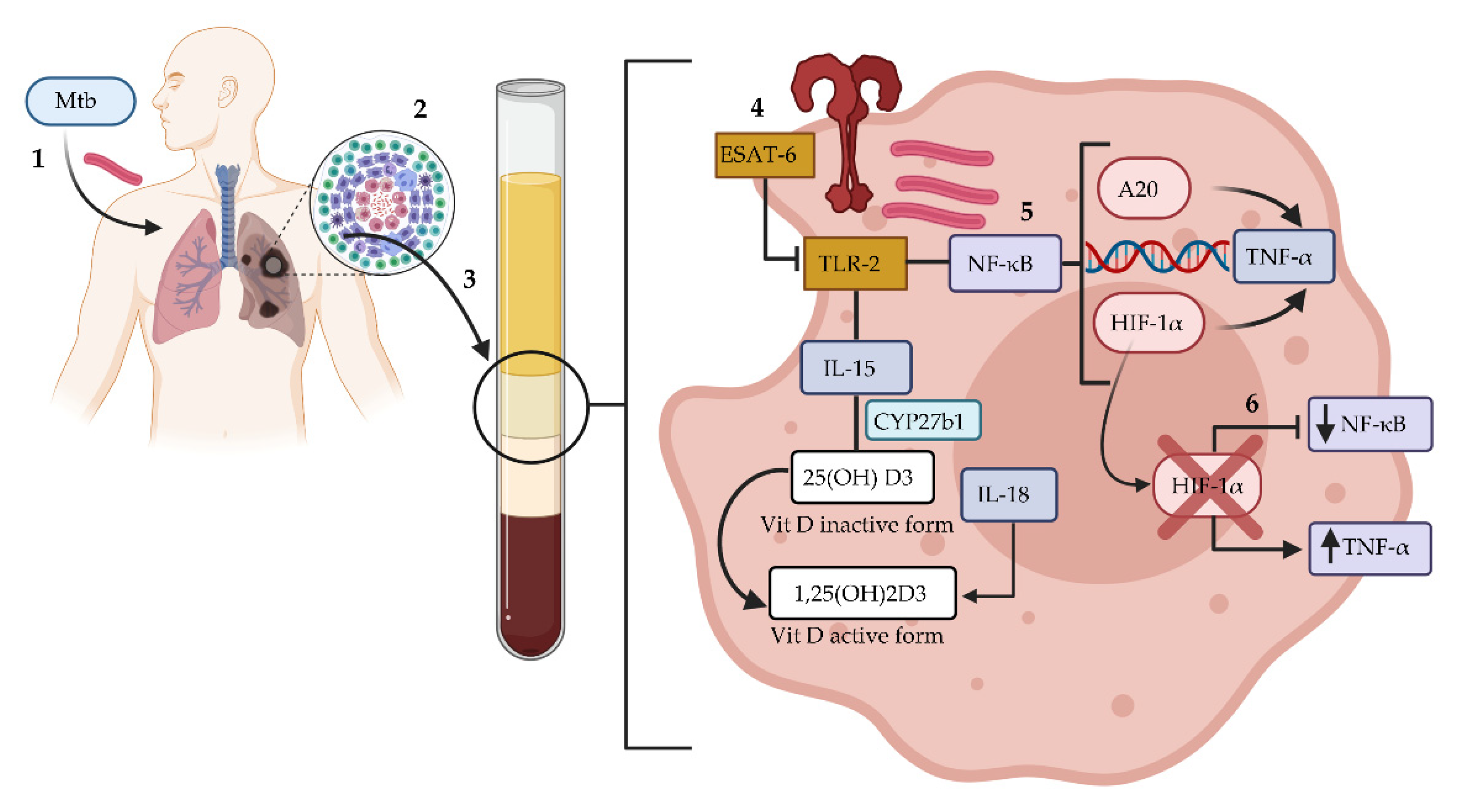
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Global Tuberculosis Report 2021. Available online: https://www.who.int/publications/digital/global-tuberculosis-report-2021/tb-diagnosis-treatment/notifications (accessed on 20 December 2021).
- Cohen, A.; Mathiasen, V.D.; Schön, T.; Wejse, C. The global prevalence of latent tuberculosis: A systematic review and meta-analysis. Eur. Respir. J. 2019, 54, 1900655. [Google Scholar] [CrossRef] [PubMed]
- Petruccioli, E.; Scriba, T.J.; Petrone, L.; Hatherill, M.; Cirillo, D.M.; Joosten, S.A.; Ottenhoff, T.H.; Denkinger, C.M.; Goletti, D. Correlates of tuberculosis risk: Predictive biomarkers for progression to active tuberculosis. Eur. Respir. J. 2016, 48, 1751–1763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, L.; Jiang, Q.; Bushkin, Y.; Subbian, S.; Tyagi, S. Biphasic Dynamics of Macrophage Immunometabolism during Mycobacterium tuberculosis Infection. mBio 2019, 10, e02550-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, X.; Sun, J. Mechanism of ESAT-6 membrane interaction and its roles in pathogenesis of Mycobacterium tuberculosis. Toxicon 2016, 116, 29–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathak, S.K.; Basu, S.; Basu, K.K.; Banerjee, A.; Pathak, S.; Bhattacharyya, A.; Kaisho, T.; Kundu, M.; Basu, J. Direct extracellular interaction between the early secreted antigen ESAT-6 of Mycobacterium tuberculosis and TLR2 inhibits TLR signaling in macrophages. Nat. Immunol. 2007, 8, 610–618. [Google Scholar] [CrossRef] [PubMed]
- Krutzik, S.R.; Hewison, M.; Liu, P.T.; Robles, J.A.; Stenger, S.; Adams, J.S.; Modlin, R.L. IL-15 Links TLR2/1-Induced Macrophage Differentiation to the Vitamin D-Dependent Antimicrobial Pathway. J. Immunol. 2008, 181, 7115–7120. [Google Scholar] [CrossRef] [PubMed]
- Selvaraj, P.; Harishankar, M.; Afsal, K. Vitamin D: Immuno-modulation and tuberculosis treatment. Can. J. Physiol. Pharmacol. 2015, 93, 377–384. [Google Scholar] [CrossRef]
- Kumar, M.; Sahu, S.K.; Kumar, R.; Subuddhi, A.; Maji, R.K.; Jana, K.; Gupta, P.; Raffetseder, J.; Lerm, M.; Ghosh, Z.; et al. MicroRNA let-7 Modulates the Immune Response to Mycobacterium tuberculosis Infection via Control of A20, an Inhibitor of the NF-κB Pathway. Cell Host Microbe 2015, 17, 345–356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, A.; Malynn, B.A. A20: Linking a complex regulator of ubiquitylation to immunity and human disease. Nat. Rev. Immunol. 2012, 12, 774–785. [Google Scholar] [CrossRef] [Green Version]
- Chu, Y.; Vahl, J.C.; Kumar, D.; Heger, K.; Bertossi, A.; Wójtowicz, E.; Soberon, V.; Schenten, D.; Mack, B.; Reutelshöfer, M.; et al. B cells lacking the tumor suppressor TNFAIP3/A20 display impaired differentiation and hyperactivation and cause inflammation and autoimmunity in aged mice. Blood 2011, 117, 2227–2236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perkins, N.D. The diverse and complex roles of NF-κB subunits in cancer. Nat. Rev. Cancer 2012, 12, 121–132. [Google Scholar] [CrossRef] [PubMed]
- Shih, V.F.-S.; Tsui, R.; Caldwell, A.; Single, A.; Hoffmann, A. A single NFκB system for both canonical and non-canonical signaling. Cell Res. 2011, 21, 86–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Dea, E.; Hoffmann, A. The Regulatory Logic of the NF-Kappa B Signaling System. Cold Spring Harb. Perspect. Biol. 2010, 2, a000216. [Google Scholar] [CrossRef] [PubMed]
- Huxford, T.; Hoffmann, A.; Ghosh, G. Understanding the Logic of IκB:NF-κB Regulation in Structural Terms. Curr. Top. Microbiol. Immunol. 2011, 349, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bandarra, D.; Biddlestone, J.; Mudie, S.; Muller, H.-A.; Rocha, S. HIF-1α restricts NF-κB dependent gene expression to control innate immunity signals. Dis. Model. Mech. 2015, 8, 169–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardoso, M.S.; Silva, T.M.; Resende, M.; Appelberg, R.; Borges, M. Lack of the Transcription Factor Hypoxia-Inducible Factor 1α (HIF-1α) in Macrophages Accelerates the Necrosis of Mycobacterium avium-Induced Granulomas. Infect. Immun. 2015, 83, 3534–3544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elks, P.M.; Brizee, S.; Van Der Vaart, M.; Walmsley, S.R.; Van Eeden, F.J.; Renshaw, S.A.; Meijer, A.H. Hypoxia Inducible Factor Signaling Modulates Susceptibility to Mycobacterial Infection via a Nitric Oxide Dependent Mechanism. PLoS Pathog. 2013, 9, e1003789. [Google Scholar] [CrossRef] [PubMed]
- Baay-Guzman, G.J.; Duran-Padilla, M.A.; Rangel-Santiago, J.; Tirado-Rodriguez, B.; Antonio-Andres, G.; Barrios-Payan, J.; Mata-Espinosa, D.; Klunder-Klunder, M.; Vega, M.I.; Hernandez-Pando, R.; et al. Dual role of hypoxia-inducible factor 1 α in experimental pulmonary tuberculosis: Its implication as a new therapeutic target. Future Microbiol. 2018, 13, 785–798. [Google Scholar] [CrossRef] [PubMed]
- Nizet, V.; Johnson, R.S. Interdependence of hypoxic and innate immune responses. Nat. Rev. Immunol. 2009, 9, 609–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egners, A.; Erdem, M.; Cramer, T. The Response of Macrophages and Neutrophils to Hypoxia in the Context of Cancer and Other Inflammatory Diseases. Mediat. Inflamm. 2016, 2016, 2053646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braverman, J.; Sogi, K.M.; Benjamin, D.; Nomura, D.K.; Stanley, S.A. HIF-1α Is an Essential Mediator of IFN-γ–Dependent Immunity to Mycobacterium tuberculosis. J. Immunol. 2016, 197, 1287–1297. [Google Scholar] [CrossRef] [Green Version]
- Won, D.; Park, J.Y.; Kim, H.-S.; Park, Y. Comparative Results of QuantiFERON-TB Gold In-Tube and QuantiFERON-TB Gold Plus Assays for Detection of Tuberculosis Infection in Clinical Samples. J. Clin. Microbiol. 2020, 58, e01854-19. [Google Scholar] [CrossRef] [PubMed]
- Riedhammer, C.; Halbritter, D.; Weissert, R. Peripheral Blood Mononuclear Cells: Isolation, Freezing, Thawing, and Culture. Methods Mol. Biol. 2016, 1304, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Owolabi, O.; Agbla, S.; Owiafe, P.K.; Donkor, S.; Togun, T.; Sillah, A.K.; Ota, M.O.C.; Sutherland, J.S. Elevated serum 25-hydroxy (OH) vitamin D levels are associated with risk of TB progression in Gambian adults. Tuberculosis 2016, 98, 86–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luc, J.G.Y.; Paulin, R.; Zhao, J.Y.; Freed, D.H.; Michelakis, E.D.; Nagendran, J. 2-Methoxyestradiol: A Hormonal Metabolite Modulates Stimulated T-Cells Function and proliferation. Transplant. Proc. 2015, 47, 2057–2066. [Google Scholar] [CrossRef] [PubMed]
- Duncan, G.S.; Brenner, D.; Tusche, M.W.; Brüstle, A.; Knobbe, C.B.; Elia, A.J.; Mock, T.; Bray, M.R.; Krammer, P.H.; Mak, T.W. 2-Methoxyestradiol inhibits experimental autoimmune encephalomyelitis through suppression of immune cell activation. Proc. Natl. Acad. Sci. USA 2012, 109, 21034–21039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaiprasongsuk, A.; Janjetovic, Z.; Kim, T.-K.; Schwartz, C.J.; Tuckey, R.C.; Tang, E.K.Y.; Raman, C.; Panich, U.; Slominski, A.T. Hydroxylumisterols, Photoproducts of Pre-Vitamin D3, Protect Human Keratinocytes against UVB-Induced Damage. Int. J. Mol. Sci. 2020, 21, 9374. [Google Scholar] [CrossRef]
- Forrellad, M.A.; Klepp, L.I.; Gioffré, A.; Sabio y García, J.; Morbidoni, H.R.; de la Paz Santangelo, M.; Cataldi, A.A.; Bigi, F. Virulence factors of the Mycobacterium tuberculosis complex. Virulence 2013, 4, 3–66. [Google Scholar] [CrossRef] [Green Version]
- Peyssonnaux, C.; Datta, V.; Cramer, T.; Doedens, A.; Theodorakis, E.A.; Gallo, R.L.; Hurtado-Ziola, N.; Nizet, V.; Johnson, R.S. HIF-1α expression regulates the bactericidal capacity of phagocytes. J. Clin. Investig. 2005, 115, 1806–1815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patidar, M.; Yadav, N.; Dalai, S.K. Interleukin 15: A key cytokine for immunotherapy. Cytokine Growth Factor Rev. 2016, 31, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekara, S.; Anupama, K.R.; Sambarey, A.; Chandra, N. High IL-6 and low IL-15 levels mark the presence of TB infection: A preliminary study. Cytokine 2016, 81, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Meraviglia, S.; Caccamo, N.R.; Salerno, A.; Sireci, G.; Dieli, F. Partial and Ineffective Activation of Vγ9Vδ2 T Cells by Mycobacterium tuberculosis-Infected Dendritic Cells. J. Immunol. 2010, 185, 1770–1776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, R.; Yang, E.; Shen, L.; Modlin, R.L.; Shen, H.; Chen, Z.W. IL-12+IL-18 Cosignaling in Human Macrophages and Lung Epithelial Cells Activates Cathelicidin and Autophagy, Inhibiting Intracellular Mycobacterial Growth. J. Immunol. 2018, 200, 2405–2417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, J.; Grando, S.A.; Li, Y.C. Regulation of IL-1 Family Cytokines IL-1α, IL-1 Receptor Antagonist, and IL-18 by 1,25-Dihydroxyvitamin D3 in Primary Keratinocytes. J. Immunol. 2006, 176, 3780–3787. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dinarello, C.A.; Novick, D.; Kim, S.; Kaplanski, G. Interleukin-18 and IL-18 Binding Protein. Front. Immunol. 2013, 4, 289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buha, I.; Škodrić-Trifunović, V.; Adžić-Vukičević, T.; Ilić, A.; Blanka-Protić, A.B.; Stjepanovic, M.; Anđelković, M.; Vreća, M.; Lazovic, J.M.; Spasovski, V.; et al. Relevance of TNF-α, IL-6 and IRAK1 gene expression for assessing disease severity and therapy effects in tuberculosis patients. J. Infect. Dev. Ctries. 2019, 13, 419–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fabri, M.; Modlin, R.L. A Vitamin for Autophagy. Cell Host Microbe 2009, 6, 201–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, J.H. Vitamin D as an inducer of cathelicidin antimicrobial peptide expression: Past, present and future. J. Steroid Biochem. Mol. Biol. 2010, 121, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.T.; Stenger, S.; Li, H.; Wenzel, L.; Tan, B.H.; Krutzik, S.R.; Ochoa, M.T.; Schauber, J.; Wu, K.; Meinken, C.; et al. Toll-Like Receptor Triggering of a Vitamin D-Mediated Human Antimicrobial Response. Science 2006, 311, 1770–1773. [Google Scholar] [CrossRef] [PubMed]
- Cavalcanti, Y.V.N.; Brelaz, M.C.A.; Neves, J.K.D.A.L.; Ferraz, J.C.; Pereira, V.R.A. Role of TNF-Alpha, IFN-Gamma, and IL-10 in the Development of Pulmonary Tuberculosis. Pulm. Med. 2012, 2012, 745483. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, D.A.A.; Da Silva, M.V.; Barros, C.C.O.; Alexandre, P.B.D.; Timóteo, R.P.; Catarino, J.S.; Sales-Campos, H.; Machado, J.R.; Rodrigues, D.B.R.; Oliveira, C.J.; et al. TNF-α blockade impairs in vitro tuberculous granuloma formation and down modulate Th1, Th17 and Treg cytokines. PLoS ONE 2018, 13, e0194430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, A.N.; Mehra, S.; Kaushal, D. Role of Interleukin 6 in Innate Immunity to Mycobacterium tuberculosis Infection. J. Infect. Dis. 2013, 207, 1253–1261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Zhang, L.; Zhang, Y.; Liu, Y.; Zhang, H.; Wei, L.; Shen, T.; Jiang, C.; Zhu, D. A20 deficiency leads to angiogenesis of pulmonary artery endothelial cells through stronger NF-κB activation under hypoxia. J. Cell. Mol. Med. 2016, 20, 1319–1328. [Google Scholar] [CrossRef] [PubMed]
- Shell, S.S.; Prestwich, E.G.; Baek, S.-H.; Shah, R.R.; Sassetti, C.M.; Dedon, P.C.; Fortune, S.M. DNA Methylation Impacts Gene Expression and Ensures Hypoxic Survival of Mycobacterium tuberculosis. PLoS Pathog. 2013, 9, e1003419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belton, M.; Brilha, S.; Manavaki, R.; Mauri, F.; Nijran, K.; Hong, Y.T.; Patel, N.H.; Dembek, M.; Tezera, L.; Green, J.; et al. Hypoxia and Tissue Destruction in Pulmonary TB. Thorax 2016, 71, 1145–1153. [Google Scholar] [CrossRef] [Green Version]
- Martens, A.; Van Loo, G. A20 at the Crossroads of Cell Death, Inflammation, and Autoimmunity. Cold Spring Harb. Perspect. Biol. 2020, 12, a036418. [Google Scholar] [CrossRef] [PubMed]
- Tannahill, G.M.; Curtis, A.M.; Adamik, J.; Palsson-McDermott, E.M.; McGettrick, A.F.; Goel, G.; Frezza, C.; Bernard, N.J.; Kelly, B.; Foley, N.H.; et al. Succinate is an inflammatory signal that induces IL-1β through HIF-1α. Nature 2013, 496, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Shalova, I.N.; Lim, J.Y.; Chittezhath, M.; Zinkernagel, A.S.; Beasley, F.; Hernández-Jiménez, E.; Toledano, V.; Cubillos-Zapata, C.; Rapisarda, A.; Chen, J.; et al. Human Monocytes Undergo Functional Re-programming during Sepsis Mediated by Hypoxia-Inducible Factor-1α. Immunity 2015, 42, 484–498. [Google Scholar] [CrossRef] [Green Version]
- Domingo-Gonzalez, R.; Das, S.; Griffiths, K.L.; Ahmed, M.; Bambouskova, M.; Gopal, R.; Gondi, S.; Muñoz-Torrico, M.; Salazar-Lezama, M.A.; Cruz-Lagunas, A.; et al. Interleukin-17 limits hypoxia-inducible factor 1α and development of hypoxic granulomas during tuberculosis. JCI Insight 2017, 2, e92973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rius, J.; Guma, M.; Schachtrup, C.; Akassoglou, K.; Zinkernagel, A.S.; Nizet, V.; Johnson, R.S.; Haddad, G.G.; Karin, M. NF-κB links innate immunity to the hypoxic response through transcriptional regulation of HIF-1α. Nature 2008, 453, 807–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anand, R.J.; Gribar, S.C.; Li, J.; Kohler, J.W.; Branca, M.F.; Dubowski, T.; Sodhi, C.P.; Hackam, D.J. Hypoxia causes an increase in phagocytosis by macrophages in a HIF-1α-dependent manner. J. Leukoc. Biol. 2007, 82, 1257–1265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prosser, G.; Brandenburg, J.; Reiling, N.; Barry, C.E.; Wilkinson, R.J.; Wilkinson, K.A. The bacillary and macrophage response to hypoxia in tuberculosis and the consequences for T cell antigen recognition. Microbes Infect. 2017, 19, 177–192. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Healthy Control | LTBI | TB | |
|---|---|---|---|
| n = 19 | n = 17 | n = 42 | |
| Gender (M/F) | 6/13 | 4/13 | 30/12 |
| Age (mean ± SD) | 31.6 ± 10.7 | 33.0 ± 13.6 | 35.1 ± 13.8 |
| TST+ (>5 mm) | 7 | 12 | – |
| IGRAs+ | 0 | 17 | – |
| BAAR+ | – | – | 42 |
| Culture+ | – | – | 12 |
| Xpert TB/RIF+ | – | – | 28 |
| X-ray+ | – | – | 39 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Oliveira Rezende, A.; Sabóia, R.S.; da Costa, A.C.; da Silva Monteiro, D.M.P.; Zagmignan, A.; Santiago, L.Â.M.; Carvalho, R.C.; Pereira, P.V.S.; Junqueira-Kipnis, A.P.; de Sousa, E.M. Restricted Activation of the NF-κB Pathway in Individuals with Latent Tuberculosis Infection after HIF-1α Blockade. Biomedicines 2022, 10, 817. https://doi.org/10.3390/biomedicines10040817
de Oliveira Rezende A, Sabóia RS, da Costa AC, da Silva Monteiro DMP, Zagmignan A, Santiago LÂM, Carvalho RC, Pereira PVS, Junqueira-Kipnis AP, de Sousa EM. Restricted Activation of the NF-κB Pathway in Individuals with Latent Tuberculosis Infection after HIF-1α Blockade. Biomedicines. 2022; 10(4):817. https://doi.org/10.3390/biomedicines10040817
Chicago/Turabian Stylede Oliveira Rezende, Aline, Rafaella Santos Sabóia, Adeliane Castro da Costa, Diana Messala Pinheiro da Silva Monteiro, Adrielle Zagmignan, Luis Ângelo Macedo Santiago, Rafael Cardoso Carvalho, Paulo Vitor Soeiro Pereira, Ana Paula Junqueira-Kipnis, and Eduardo Martins de Sousa. 2022. "Restricted Activation of the NF-κB Pathway in Individuals with Latent Tuberculosis Infection after HIF-1α Blockade" Biomedicines 10, no. 4: 817. https://doi.org/10.3390/biomedicines10040817