
Activation of Voltage-Gated Na+ Current by GV-58, a Known Activator of CaV Channels


Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals, Drugs, and Solutions
2.2. Cell Preparations
2.3. Electrophysiological Measurements
2.4. Data Analyses
2.5. Curve-Fitting Procedures and Statistical Analyses
3. Results
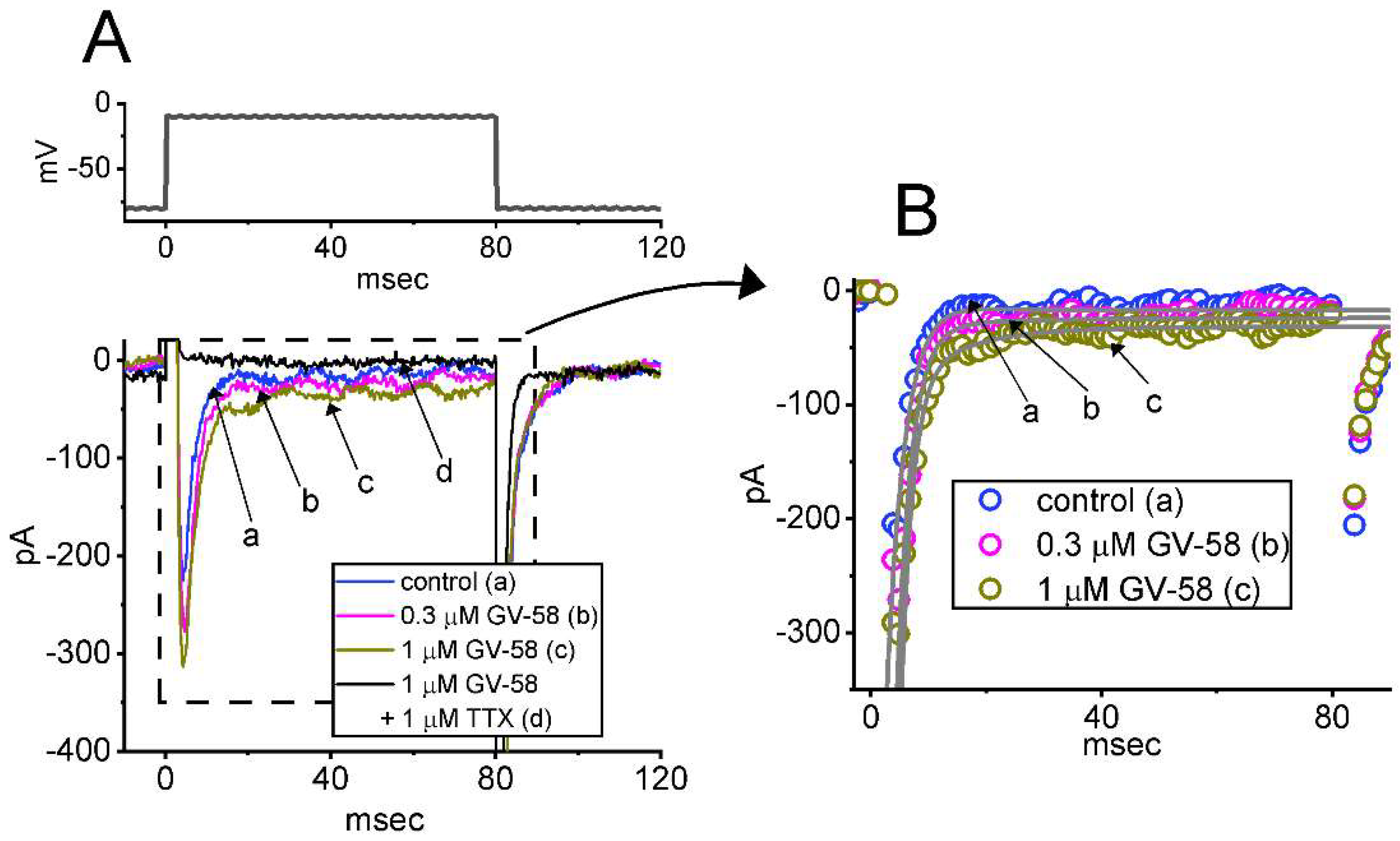
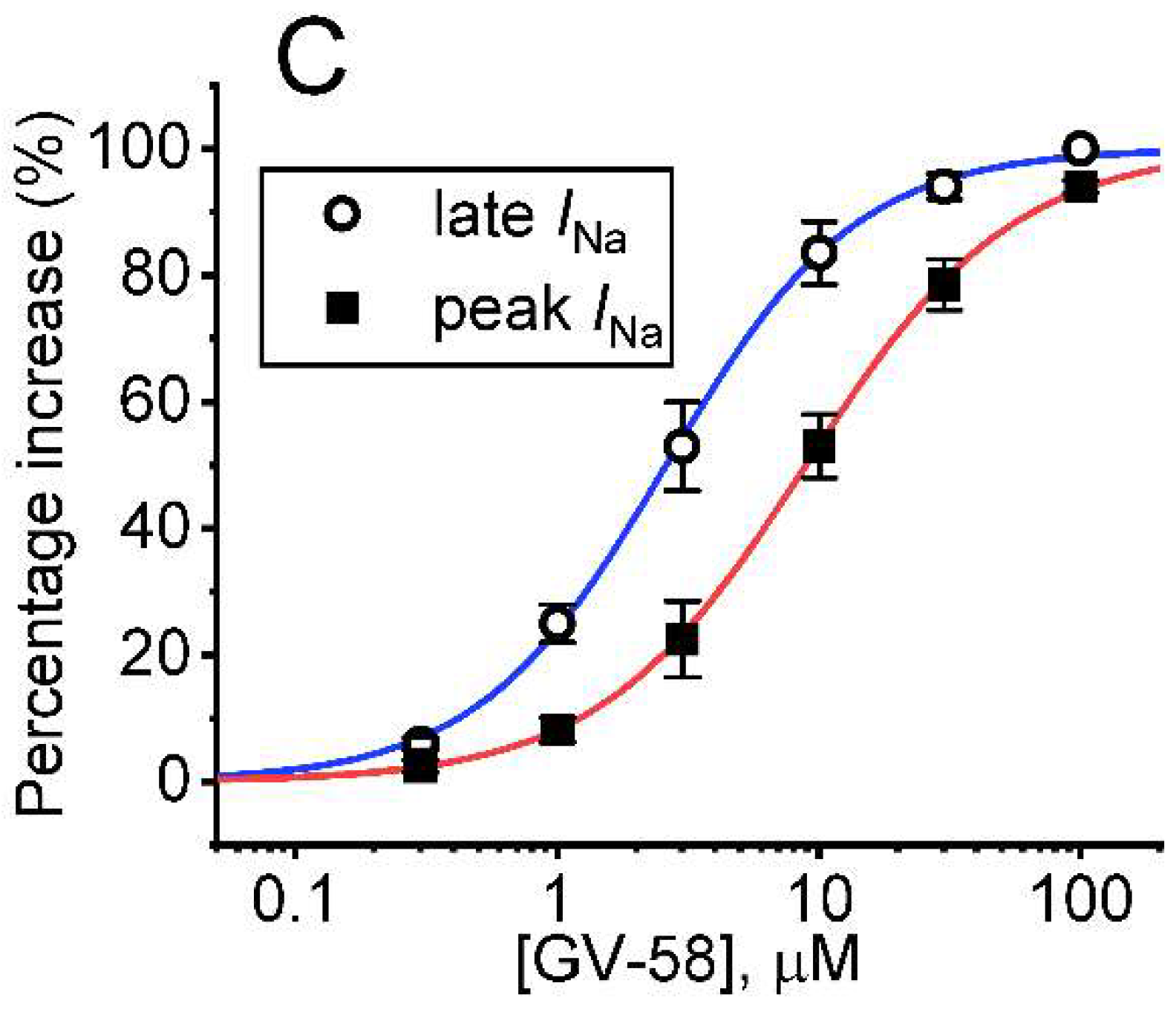
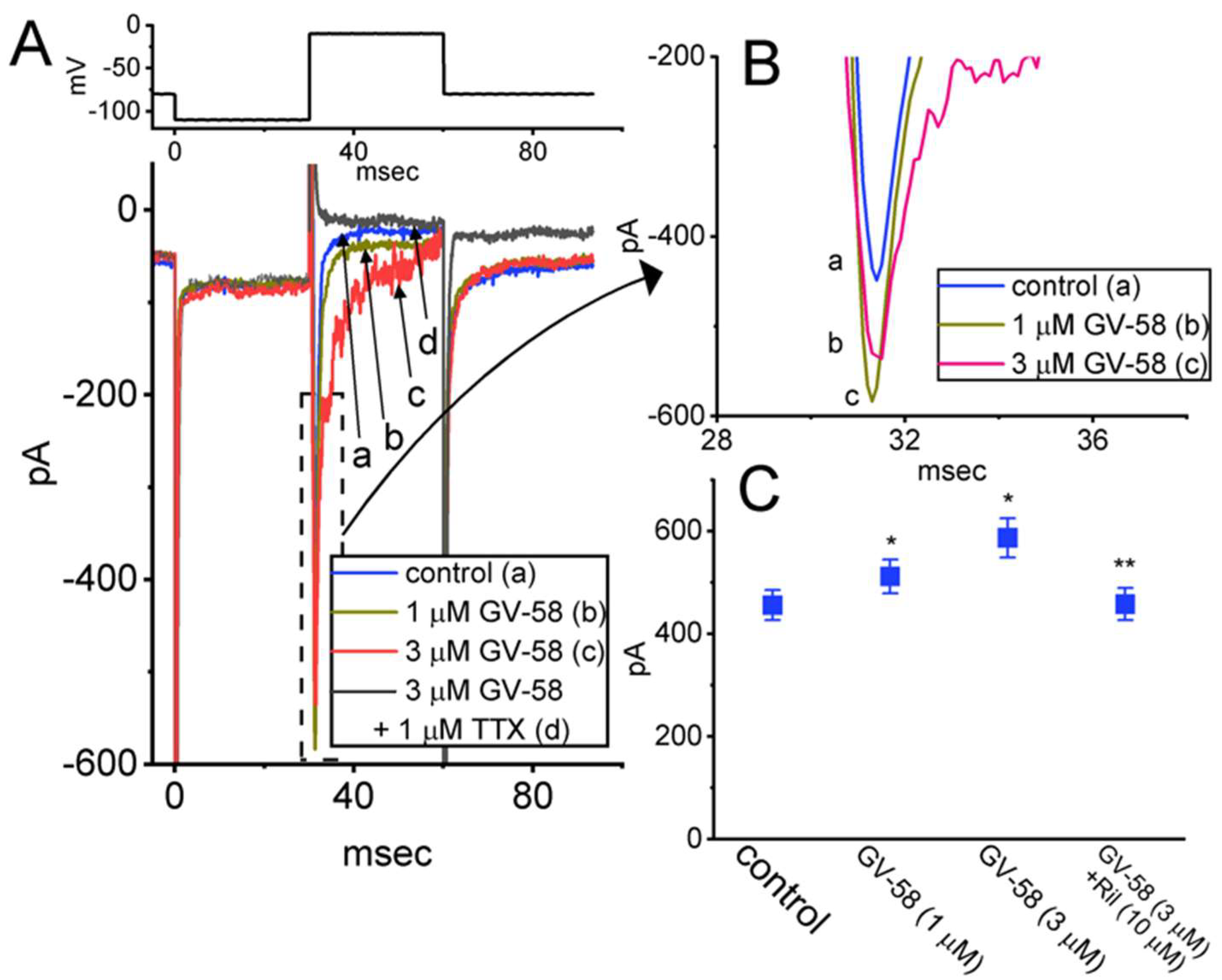
3.1. Stimulatory Effect of GV-58 on Voltage-Gated Na+ Current Measured from Pituitary GH3 Cells
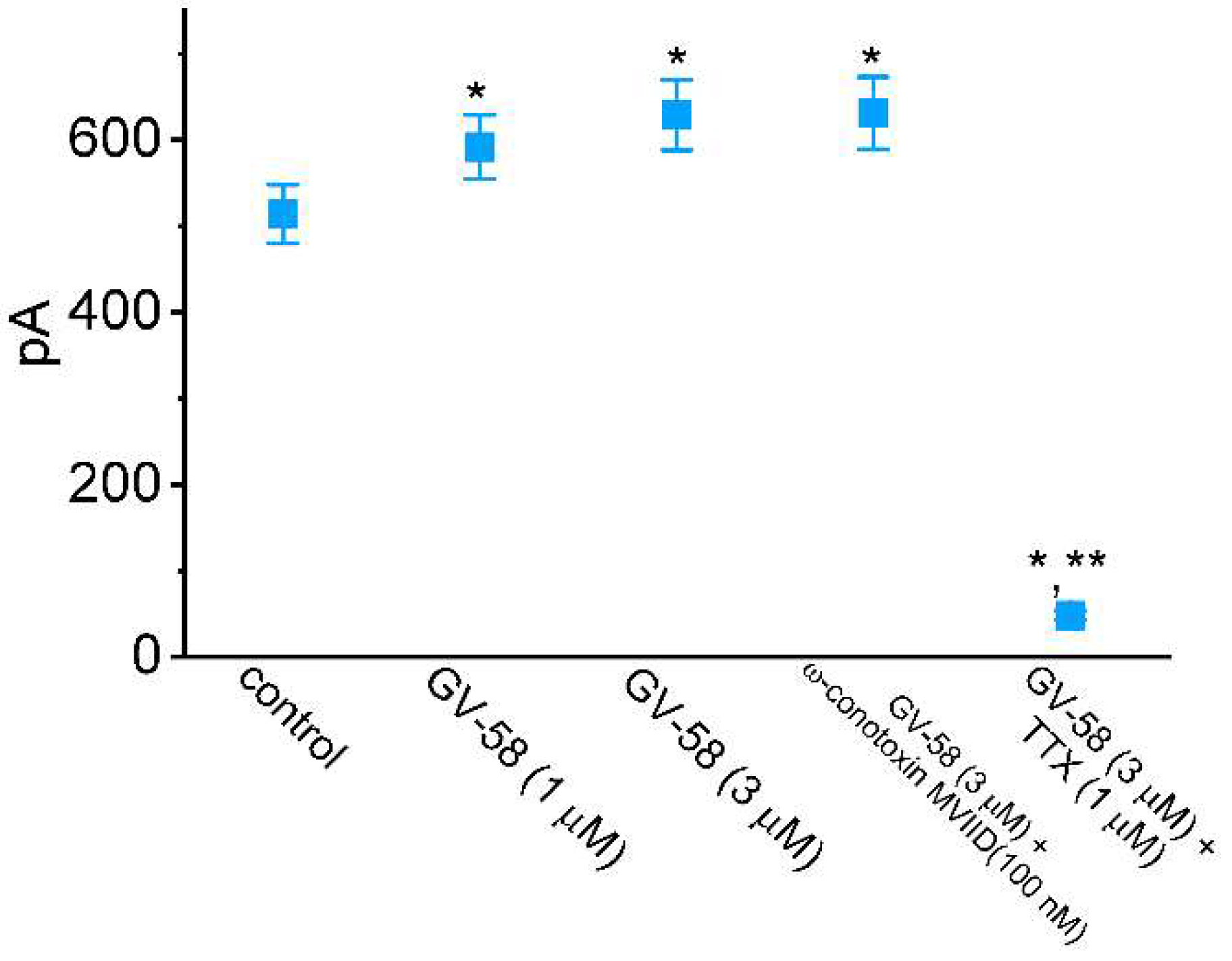
3.2. Comparisons of Effects of GV-58, ω-Conotoxin MVIID, and Tetrodotoxin (TTX) on Peak INa
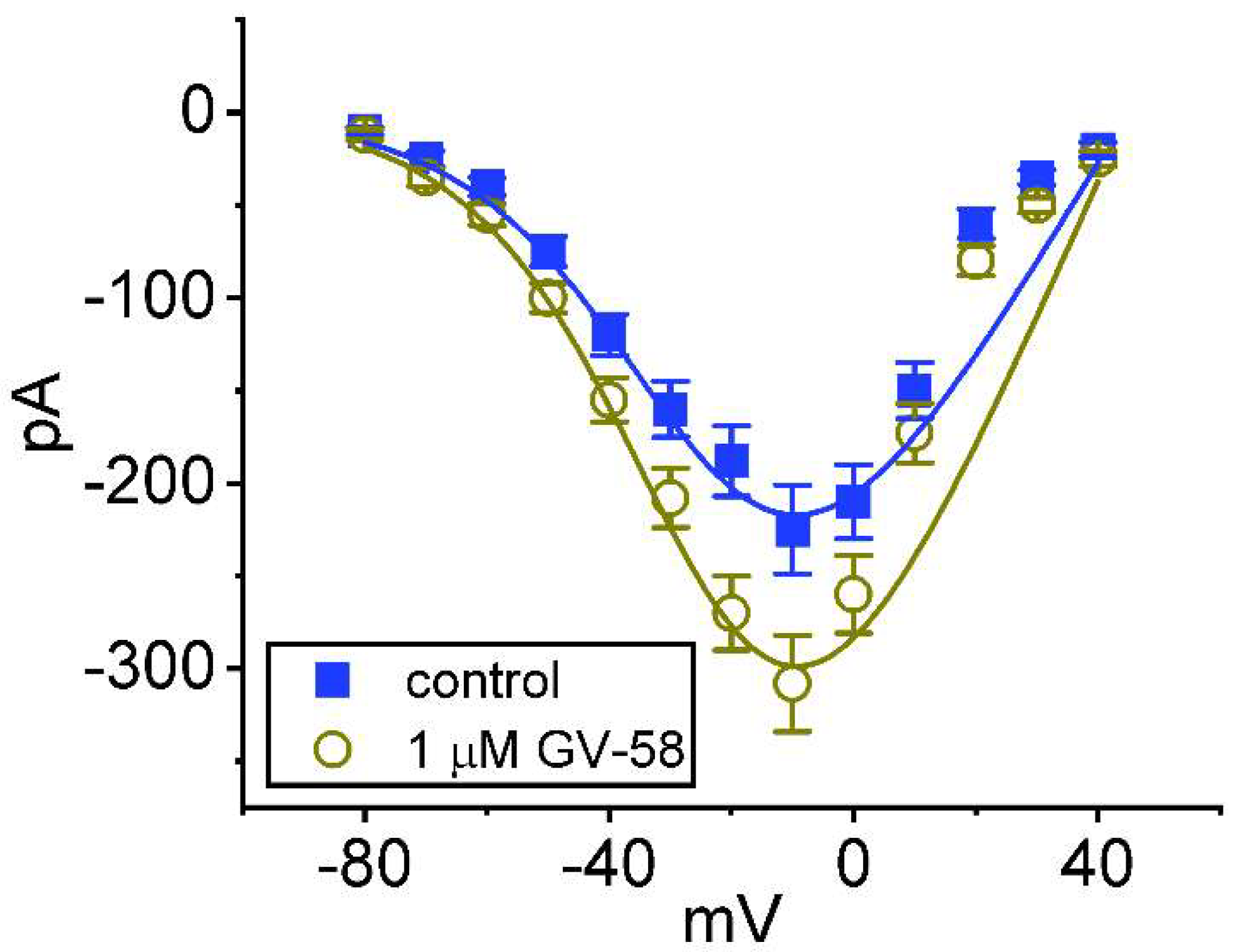
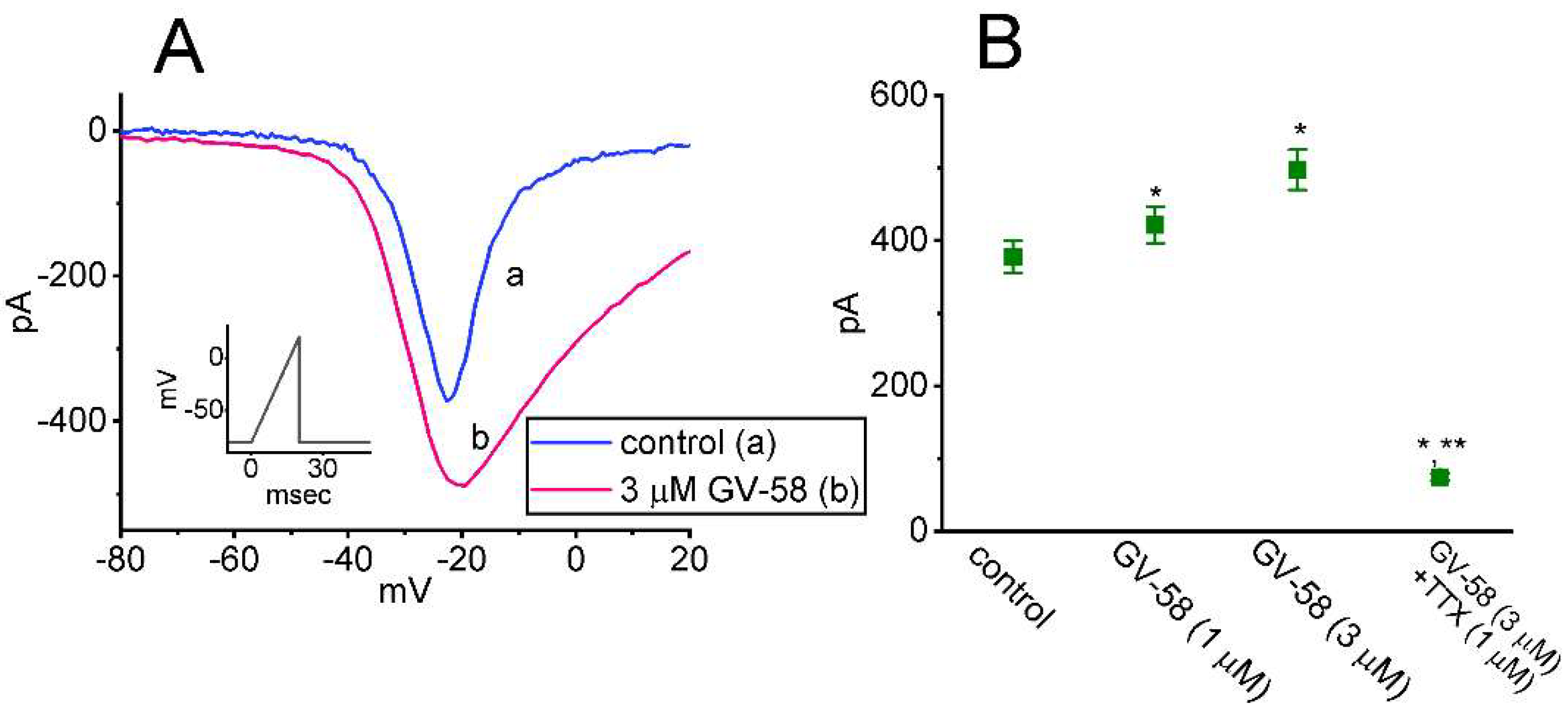
3.3. Effect of GV-58 on the Steady-State Current–Voltage (I–V) Relationship of Peak INa
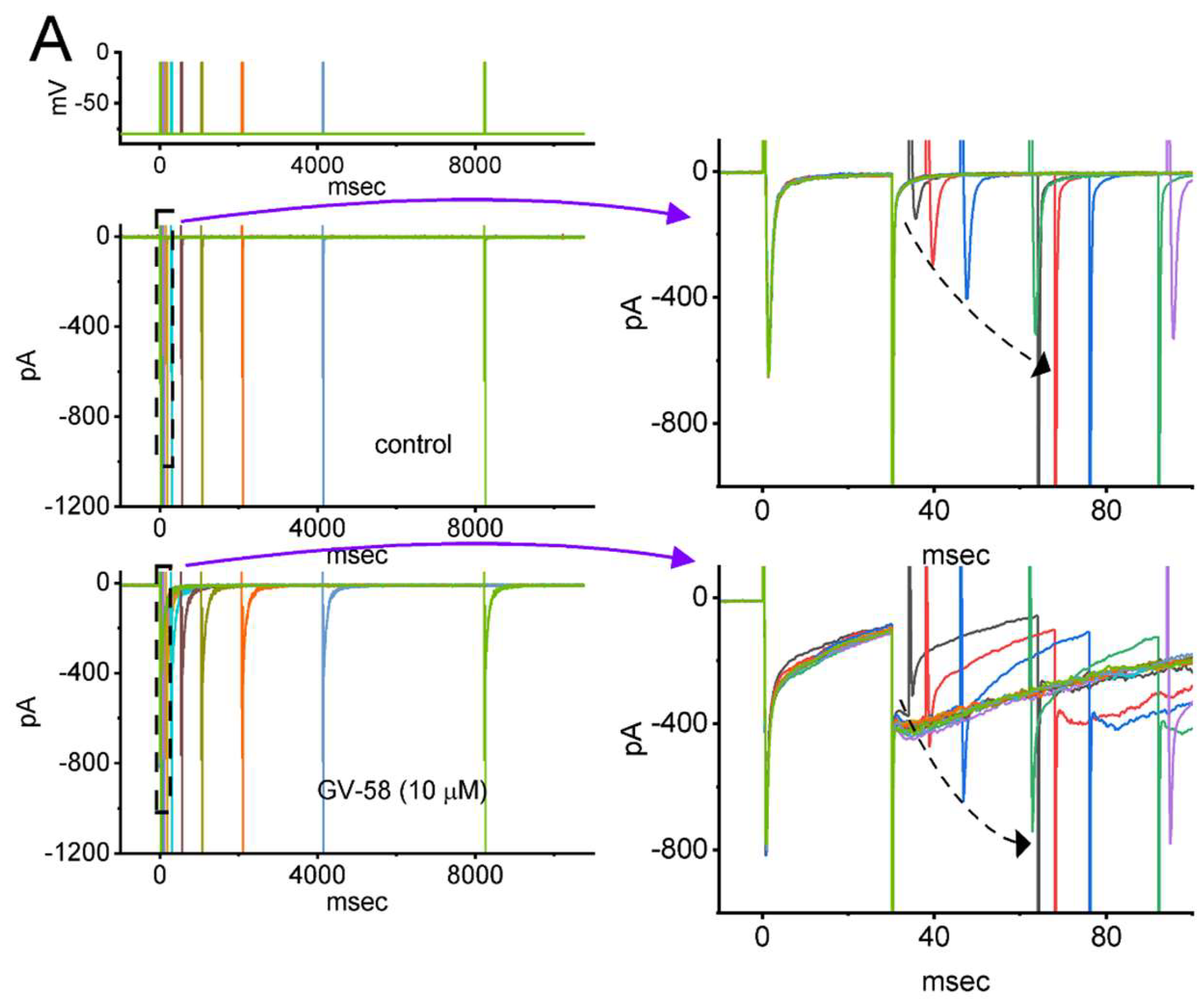
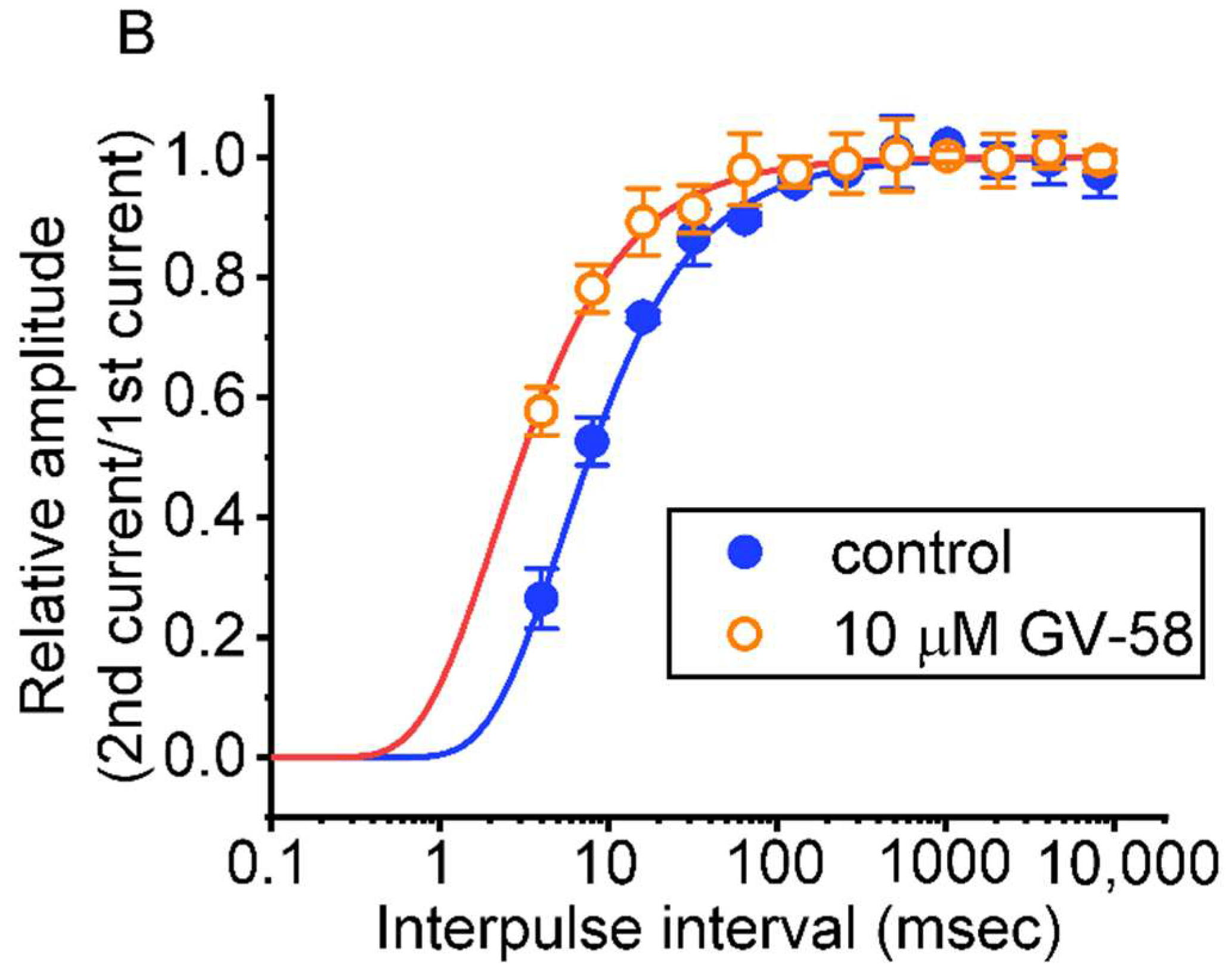
3.4. Effect of GV-58 on the Recovery from INa Inactivation Evoked during Varying Interpulse Intervals
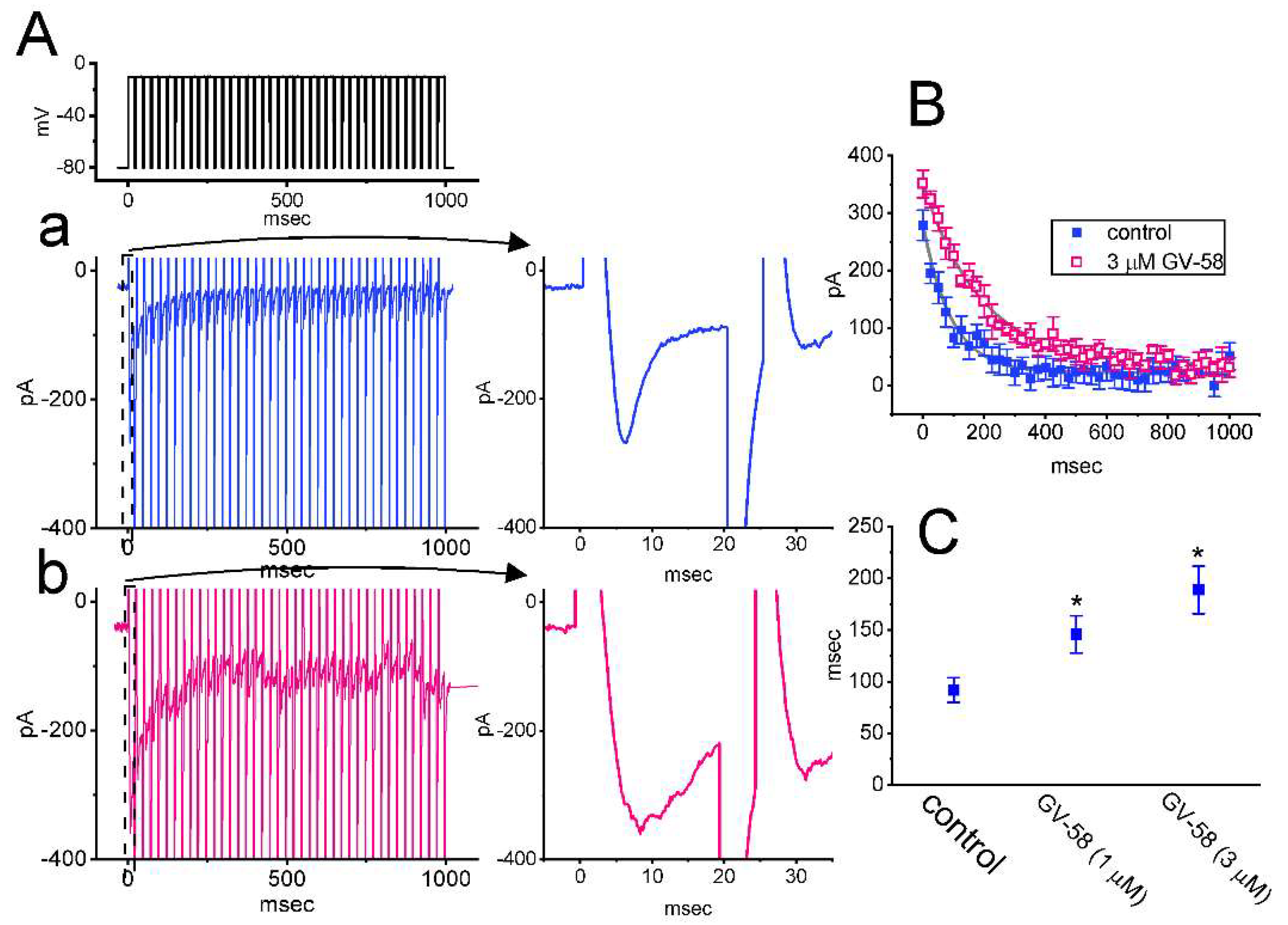
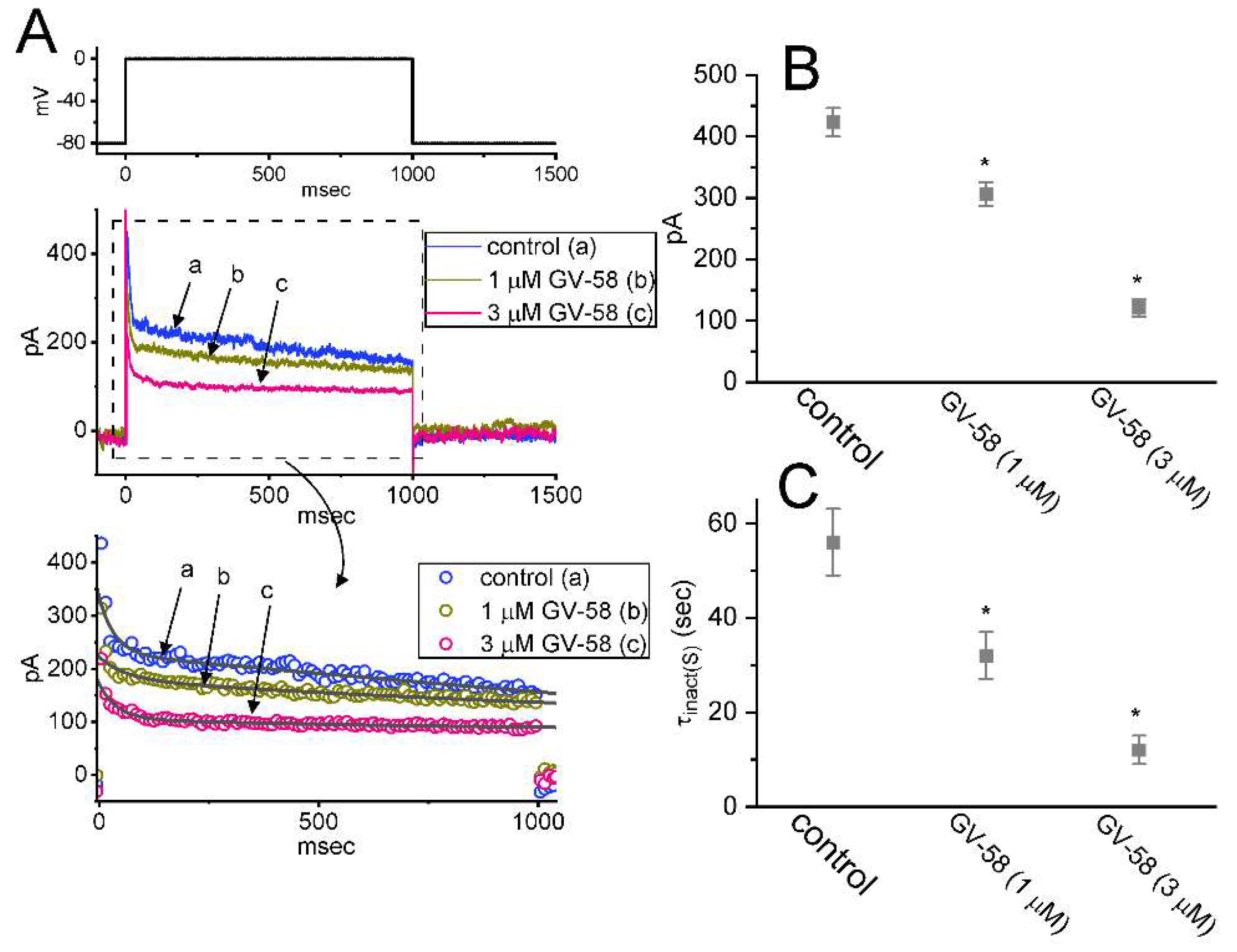
3.5. GV-58 Effect on INa Decay Evoked during a Train of Depolarizing Command Voltages in GH3 Cells
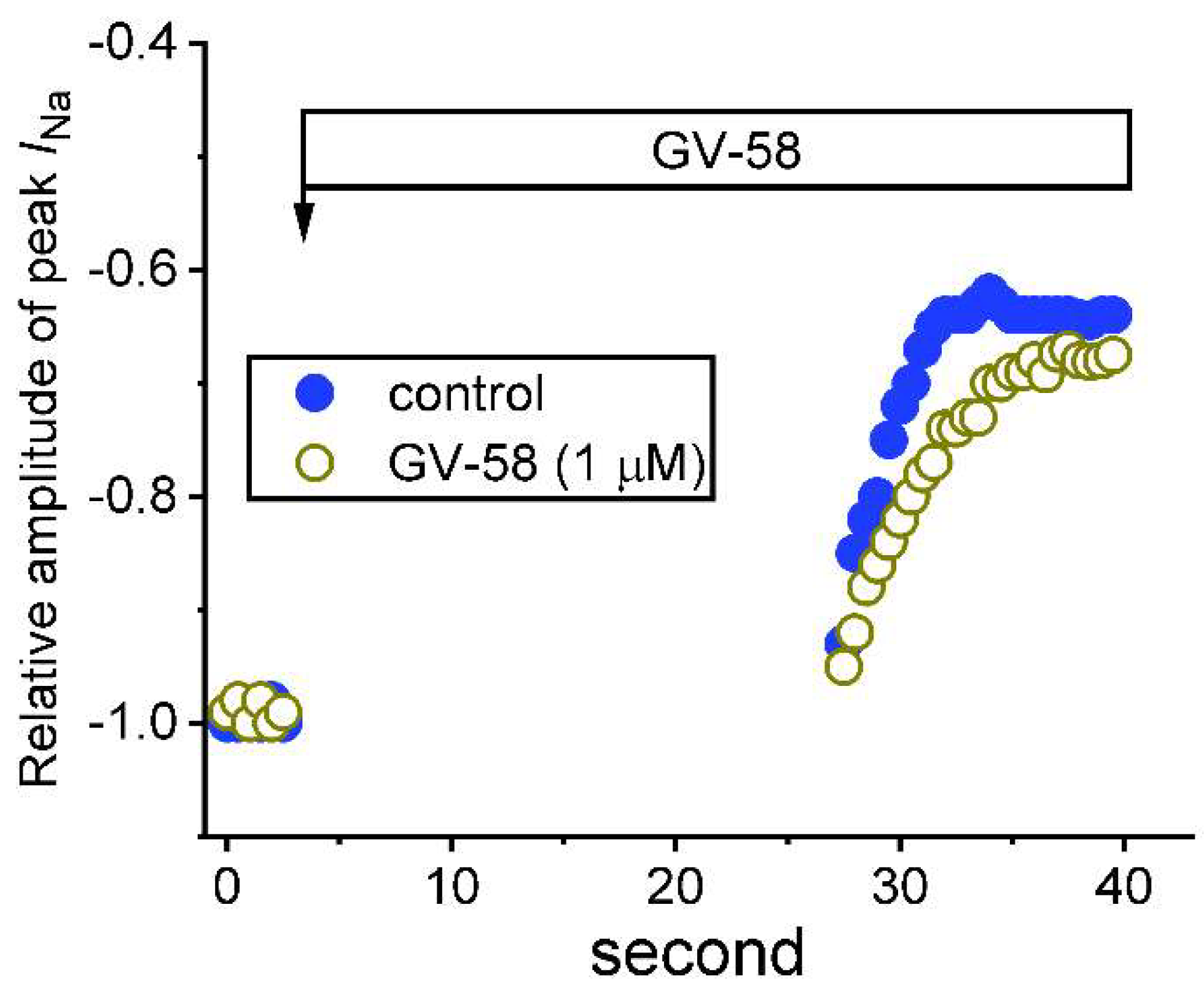
3.6. Use-Dependence of GV-58-Induced Stimulation of Peak INa
3.7. Stimulatory Effect of GV-58 on Resurgent INa (INa(R)) Seen in GH3 Cells
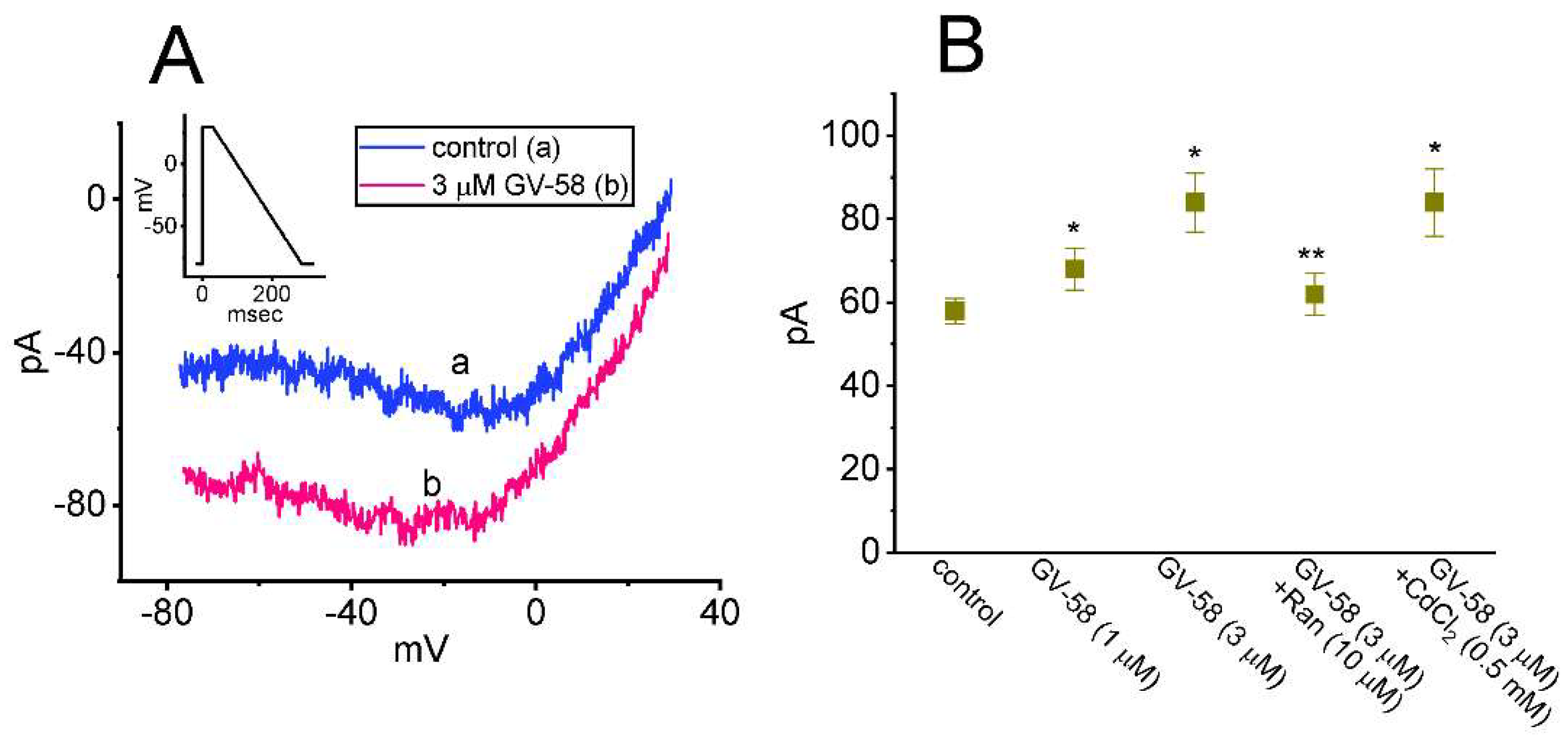
3.8. Effect of GV-58 on the Window Component of INa (INa(W)) Recorded from GH3 Cells
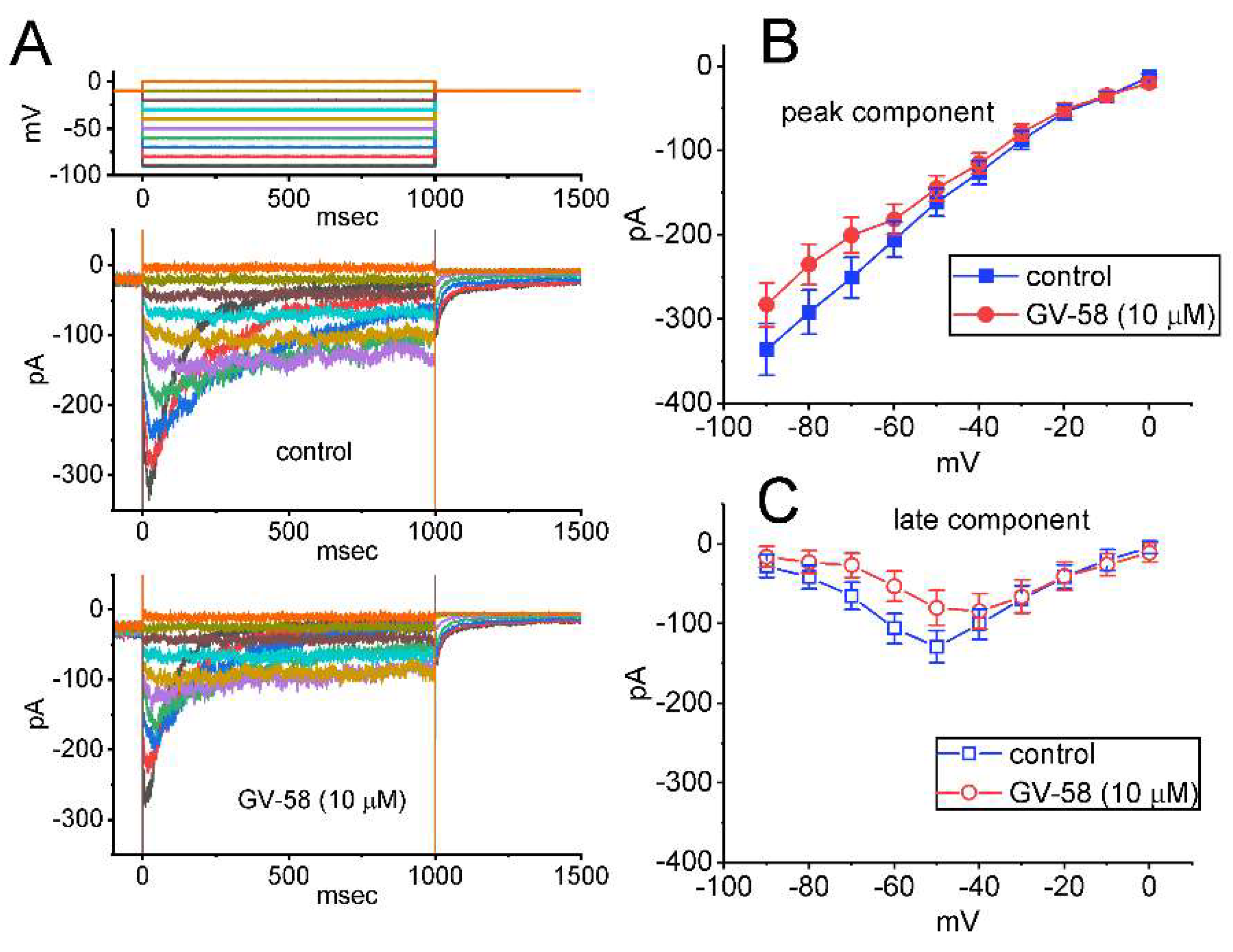
3.9. Inhibitory Effect of GV-58 on A-Type K+ Current (IK(A)) in GH3 Cells
3.10. Mild Suppression of Erg-Mediated K+ Current (IK(erg)) Produced by GV-58
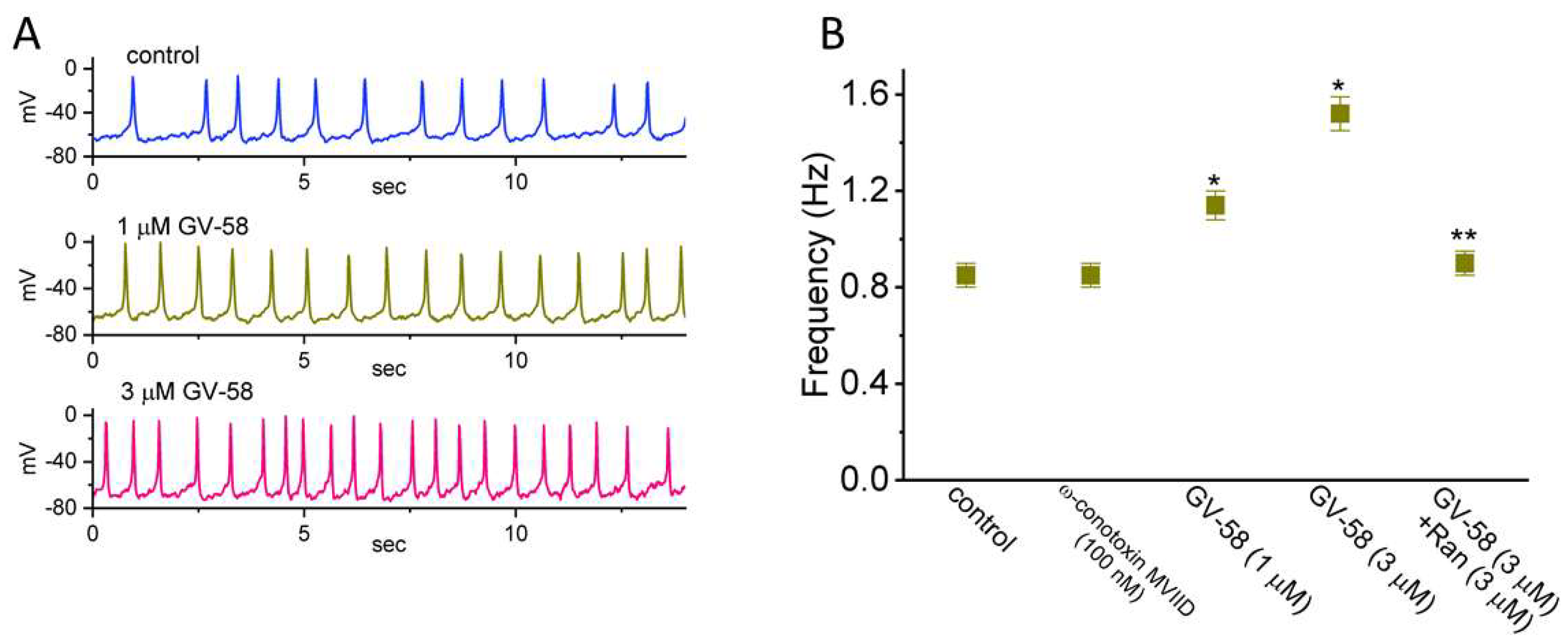
3.11. Stimulatory Effect of GV-58 on Spontaneous Action Potentials (Aps) Recorded from GH3 Cells
3.12. Effect of GV-58 on INa Inherently in NSC-34 Motor Neuron-like Cells
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ANOVA | analysis of variance |
| AP | action potential |
| CaV channel | voltage-gated Ca2+ channel |
| EC50 | concentration required for 50% stimulation |
| erg | ether-à-go-go-related gene |
| GV-58 | (2R)-2-[(6-{[(5-methylthiophen-2-yl)methyl]amino}-9-propyl-9H-purin-2-yl)amino]butan-1-ol |
| I–V | current versus voltage |
| ICa,L | L-type Ca2+ current |
| IK(A) | A-type K+ current |
| IK(erg) | erg-mediated K+ current |
| INa | voltage-gated Na+ current |
| INa(R) | resurgent INa |
| INa(W) | window INa |
| NaV channel | voltage-gated Na+ channel |
| Ran | ranolazine |
| Ril | riluzole |
| τinact(S) | slow component in the inactivation rate constant |
| TTX | tetrodotoxin |
References
- Tarr, T.B.; Valdomir, G.; Liang, M.; Wipf, P.; Meriney, S.D. New calcium channel agonists as potential therapeutics in Lambert-Eaton myasthenic syndrome and other neuromuscular diseases. Ann. N. Y. Acad. Sci. 2012, 1275, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Tarr, T.B.; Malick, W.; Liang, M.; Valdomir, G.; Frasso, M.; Lacomis, D.; Reddel, S.W.; Garcia-Ocano, A.; Wipf, P.; Meriney, S.D. Evaluation of a novel calcium channel agonist for therapeutic potential in Lambert-Eaton myasthenic syndrome. J. Neurosci. 2013, 33, 10559–10567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarr, T.B.; Lacomis, D.; Reddel, S.W.; Liang, M.; Valdomir, G.; Frasso, M.; Wipf, P.; Meriney, S.D. Complete reversal of Lambert-Eaton myasthenic syndrome synaptic impairment by the combined use of a K+ channel blocker and a Ca2+ channel agonist. J. Physiol. 2014, 592, 3687–3696. [Google Scholar] [CrossRef] [PubMed]
- Black, B.J.; Atmaramani, R.; Pancrazio, J.J. Spontaneous and Evoked Activity from Murine Ventral Horn Cultures on Microelectrode Arrays. Front. Cell Neurosci. 2017, 11, 304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tyagi, S.; Bendrick, T.R.; Filipova, D.; Papadopoulos, S.; Bannister, R.A. A mutation in Ca(V)2.1 linked to a severe neurodevelopmental disorder impairs channel gating. J. Gen. Physiol. 2019, 151, 850–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, M.; Tarr, T.B.; Bravo-Altamirano, K.; Valdomir, G.; Rensch, G.; Swanson, L.; DeStefino, N.R.; Mazzarisi, C.M.; Olszewski, R.A.; Wilson, G.M.; et al. Synthesis and biological evaluation of a selective N- and p/q-type calcium channel agonist. ACS Med. Chem. Lett. 2012, 3, 985–990. [Google Scholar] [CrossRef] [Green Version]
- Tarr, T.B.; Wipf, P.; Meriney, S.D. Synaptic Pathophysiology and Treatment of Lambert-Eaton Myasthenic Syndrome. Mol. Neurobiol. 2015, 52, 456–463. [Google Scholar] [CrossRef] [Green Version]
- Meriney, S.D.; Tarr, T.B.; Ojala, K.S.; Wu, M.; Li, Y.; Lacomis, D.; Garcia-Ocaña, A.; Liang, M.; Valdomir, G.; Wipf, P.; et al. Lambert-Eaton myasthenic syndrome: Mouse passive-transfer model illuminates disease pathology and facilitates testing therapeutic leads. Ann. N. Y. Acad. Sci. 2018, 1412, 73–81. [Google Scholar] [CrossRef]
- Wu, M.; White, H.V.; Boehm, B.; Meriney, C.J.; Kerrigan, K.; Frasso, M.; Liang, M.; Gotway, E.M.; Wilcox, M.R.; Johnson, J.W.; et al. New Cav2 calcium channel gating modifiers with agonist activity and therapeutic potential to treat neuromuscular disease. Neuropharmacology 2018, 131, 176–189. [Google Scholar] [CrossRef]
- Catterall, W.A.; Goldin, A.L.; Waxman, S.G. International Union of Pharmacology.XLVII.Nomenclature and structure-function relationships of voltage-gated sodium channels. Pharmacol. Rev. 2005, 57, 397–409. [Google Scholar] [CrossRef]
- Stojilkovic, S.S.; Tabak, J.; Bertram, R. Ion channels and signaling in the pituitary gland. Endocr. Rev. 2010, 31, 845–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.-N.; Chen, B.-S.; Hsu, T.-I.; Peng, H.; Wu, Y.-H.; Lo, Y.-C. Analytical studies of rapidly inactivating and noninactivating sodium currents in differentiated NG108-15 neuronal cells. J. Theor. Biol. 2009, 259, 828–836. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.N.; Wu, Y.H.; Chen, B.S.; Lo, Y.C.; Liu, Y.C. Underlying mechanism of actions of tefluthrin, a pyrethroid insecticide, on voltage-gated ion currents and on action currents in pituitary tumor (GH3) cells and GnRH-secreting (GT1-7) neurons. Toxicology 2009, 258, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Lai, M.C.; Wu, S.N.; Huang, C.W. Telmisartan, an Antagonist of Angiotensin II Receptors, Accentuates Voltage-Gated Na(+) Currents and Hippocampal Neuronal Excitability. Front. Neurosci. 2020, 14, 902. [Google Scholar] [CrossRef]
- Chuang, T.H.; Cho, H.Y.; Wu, S.N. Effective Accentuation of Voltage-Gated Sodium Current Caused by Apocynin (4’-Hydroxy-3’-methoxyacetophenone), a Known NADPH-Oxidase Inhibitor. Biomedicines 2021, 9, 1146. [Google Scholar] [CrossRef]
- Chen, B.S.; Lo, Y.C.; Peng, H.; Hsu, T.I.; Wu, S.N. Effects of ranolazine, a novel anti-anginal drug, on ion currents and membrane potential in pituitary tumor GH(3) cells and NG108-15 neuronal cells. J. Pharmacol. Sci. 2009, 110, 295–305. [Google Scholar] [CrossRef] [Green Version]
- Chang, W.T.; Wu, S.N. Characterization of Direct Perturbations on Voltage-Gated Sodium Current by Esaxerenone, a Nonsteroidal Mineralocorticoid Receptor Blocker. Biomedicines 2021, 9, 549. [Google Scholar] [CrossRef]
- Chen, P.C.; Ruan, J.S.; Wu, S.N. Evidence of Decreased Activity in Intermediate-Conductance Calcium-Activated Potassium Channels During Retinoic Acid-Induced Differentiation in Motor Neuron-Like NSC-34 Cells. Cell. Physiol. Biochem. 2018, 48, 2374–2388. [Google Scholar] [CrossRef]
- So, E.C.; Wu, S.-N.; Lo, Y.-C.; Su, K. Differential regulation of tefluthrin and telmisartan on the gating charges of I(Na) activation and inactivation as well as on resurgent and persistent I(Na) in a pituitary cell line (GH(3)). Toxicol. Lett. 2018, 285, 104–112. [Google Scholar] [CrossRef]
- Gandía, L.; Lara, B.; Imperial, J.S.; Villarroya, M.; Albillos, A.; Maroto, R.; Garcia, A.G.; Olivera, B.M. Analogies and differences between omega-conotoxins MVIIC and MVIID: Binding sites and functions in bovine chromaffin cells. Pflugers Arch. 1997, 435, 55–64. [Google Scholar]
- Taddese, A.; Bean, B.P. Subthreshold sodium current from rapidly inactivating sodium channels drives spontaneous firing of tuberomammillary neurons. Neuron 2002, 33, 587–600. [Google Scholar] [CrossRef] [Green Version]
- Kuo, P.-C.; Kao, Z.-H.; Lee, S.-W.; Wu, S.-N. Effects of Sesamin, the Major Furofuran Lignan of Sesame Oil, on the Amplitude and Gating of Voltage-Gated Na(+) and K(+) Currents. Molecules 2020, 25, 3062. [Google Scholar] [CrossRef] [PubMed]
- Lewis, A.H.; Raman, I.M. Resurgent current of voltage-gated Na(+) channels. J. Physiol. 2014, 592, 4825–4838. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.N. Contribution of non-inactivating Na+ current induced by oxidizing agents to the firing behavior of neuronal action potentials: Experimental and theoretical studies from NG108-15 neuronal cells. Chin. J. Physiol. 2011, 54, 19–29. [Google Scholar] [CrossRef] [Green Version]
- Morris, C.E.; Boucher, P.A.; Joós, B. Left-shifted nav channels in injured bilayer: Primary targets for neuroprotective nav antagonists? Front. Pharmacol. 2012, 3, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chuang, T.-H.; Cho, H.-Y.; Wu, S.-N. The Evidence for Sparsentan-Mediated Inhibition of INa and IK(erg): Possibly Unlinked to Its Antagonism of Angiotensin II or Endothelin Type a Receptor. Biomedicines 2022, 10, 86. [Google Scholar] [CrossRef] [PubMed]
- Sanders, D.B.; Juel, V.C.; Harati, Y.; Smith, A.G.; Peltier, A.C.; Marburger, T.; Lou, J.; Pascuzzi, R.M.; Richman, D.P.; Xie, T.; et al. 3,4-diaminopyridine base effectively treats the weakness of Lambert-Eaton myasthenia. Muscle Nerve 2018, 57, 561–568. [Google Scholar] [CrossRef] [Green Version]
- Simasko, S.M. Evidence for a delayed rectifier-like potassium current in the clonal rat pituitary cell line GH3. Am. J. Physiol. 1991, 261 Pt 1, E66–E75. [Google Scholar] [CrossRef]
- Huang, C.W.; Huang, C.C.; Liu, Y.C.; Wu, S.N. Inhibitory effect of lamotrigine on A-type potassium current in hippocampal neuron-derived H19-7 cells. Epilepsia 2004, 45, 729–736. [Google Scholar] [CrossRef]
- Liu, P.Y.; Chang, W.T.; Wu, S.N. Characterization of the Synergistic Inhibition of I(K(erg)) and I(K(DR)) by Ribociclib, a Cyclin-Dependent Kinase 4/6 Inhibitor. Int. J. Mol. Sci. 2020, 21, 8078. [Google Scholar] [CrossRef]
- Cho, H.Y.; Chuang, T.H.; Wu, S.N. Effective Perturbations on the Amplitude and Hysteresis of Erg-Mediated Potassium Current Caused by 1-Octylnonyl 8-[(2-hydroxyethyl)[6-oxo-6(undecyloxy)hexyl]amino]-octanoate (SM-102), a Cationic Lipid. Biomedicines 2021, 9, 1367. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.N.; Yeh, C.C.; Huang, H.C.; So, E.C.; Lo, Y.C. Electrophysiological characterization of sodium-activated potassium channels in NG108-15 and NSC-34 motor neuron-like cells. Acta Physiol. 2012, 206, 120–134. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-J.; Lin, M.-W.; Lin, A.-A.; Wu, S.-N. Riluzole-induced block of voltage-gated Na+ current and activation of BKCa channels in cultured differentiated human skeletal muscle cells. Life Sci. 2008, 82, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Chiò, A.; Mazzini, L.; Mora, G. Disease-modifying therapies in amyotrophic lateral sclerosis. Neuropharmacology 2020, 167, 107986. [Google Scholar] [CrossRef]
- Fehlings, M.G.; Badhiwala, J.H.; Ahn, H.; Farhadi, H.F.; I Shaffrey, C.; Nassr, A.; Mummaneni, P.; Arnold, P.M.; Jacobs, W.B.; Riew, K.D.; et al. Safety and efficacy of riluzole in patients undergoing decompressive surgery for degenerative cervical myelopathy (CSM-Protect): A multicentre, double-blind, placebo-controlled, randomised, phase 3 trial. Lancet Neurol. 2021, 20, 98–106. [Google Scholar] [CrossRef]
- Simasko, S.M. A background sodium conductance is necessary for spontaneous depolarizations in rat pituitary cell line GH3. Am. J. Physiol. 1994, 266 Pt 1, C709–C719. [Google Scholar] [CrossRef]
- Wu, N.; Hsiao, C.F.; Chandler, S.H. Membrane resonance and subthreshold membrane oscillations in mesencephalic V neurons: Participants in burst generation. J. Neurosci. 2001, 21, 3729–3739. [Google Scholar] [CrossRef]
- Herrington, J.; Stern, R.C.; Evers, A.; Lingle, C. Halothane inhibits two components of calcium current in clonal (GH3) pituitary cells. J. Neurosci. 1991, 11, 2226–2240. [Google Scholar] [CrossRef]
- Lo, Y.-K.; Wu, S.-N.; Lee, C.-T.; Li, H.-F.; Chiang, H.-T. Characterization of action potential waveform-evoked L-type calcium currents in pituitary GH3 cells. Pflugers Arch. 2001, 442, 547–557. [Google Scholar] [CrossRef]
- Nicole, S.; Lory, P. New Challenges Resulting From the Loss of Function of Na(v)1.4 in Neuromuscular Diseases. Front. Pharmacol. 2021, 12, 751095. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bathing Solution | Purpose or Name | Composition |
| Bathing solution | Normal Tyrode’s solution | 136 mM NaCl, 5.4 mM KCl, 1.8 mM CaCl2, 0.53 mM MgCl2, 5.5 mM glucose, and 5.5 mM HEPES-NaOH buffer, pH 7.4 |
| Bathing solution | High-K+, Ca2+-free solution | 130 mM KCl, 10 mM NaCl, 3 mM MgCl2, 6 mM glucose, 5 mM HEPES-KOH buffer, pH 7.4 |
| Pipette solution | For recordings of K+ current or membrane potential | 130 mM K-aspartate, 20 mM KCl, 1 mM KH2PO4, 1 mM MgCl2, 3 mM Na2ATP, 0.1 mM Na2GTP, 0.1 mM EGTA, and 5 mM HEPES-KOH buffer, pH 7.2 |
| Pipette solution | For recordings of Na+ or Ca2+ current | 130 mM Cs-aspartate, 20 mM CsCl, 1 mM KH2PO4, 1 mM MgCl2, 3 mM Na2ATP, 0.1 mM Na2GTP, 0.1 mM EGTA, and 5 mM HEPES-CsOH buffer, pH 7.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, H.-Y.; Chen, P.-C.; Chuang, T.-H.; Yu, M.-C.; Wu, S.-N. Activation of Voltage-Gated Na+ Current by GV-58, a Known Activator of CaV Channels. Biomedicines 2022, 10, 721. https://doi.org/10.3390/biomedicines10030721
Cho H-Y, Chen P-C, Chuang T-H, Yu M-C, Wu S-N. Activation of Voltage-Gated Na+ Current by GV-58, a Known Activator of CaV Channels. Biomedicines. 2022; 10(3):721. https://doi.org/10.3390/biomedicines10030721
Chicago/Turabian StyleCho, Hsin-Yen, Pei-Chun Chen, Tzu-Hsien Chuang, Meng-Cheng Yu, and Sheng-Nan Wu. 2022. "Activation of Voltage-Gated Na+ Current by GV-58, a Known Activator of CaV Channels" Biomedicines 10, no. 3: 721. https://doi.org/10.3390/biomedicines10030721