
CCL5 Deficiency Enhanced Cryo–Thermal-Triggered Long-Term Anti-Tumor Immunity in 4T1 Murine Breast Cancer

Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Animal Model
2.3. The Procedures of Cryo–Thermal Therapy
2.4. Flow Cytometry (FACS) Analysis
2.5. RNA Isolation and Real-Time PCR
2.6. In Vitro Co-Culture Assay
2.7. Statistical Analysis
3. Results
3.1. CCL5 Deficiency Significantly Prolonged Long-Term Survival after Cryo–Thermal Therapy
3.2. CCL5 Deficiency Enhanced DC Activation at an Early Stage after Cryo–Thermal Therapy
3.3. CCL5 Deficiency Enhanced the Cryo–Thermal-Induced Polarization of M1 Macrophages
3.4. CCL5 Deficiency Induced the Reduction in Immunosuppressive MDSCs after Cryo–Thermal Therapy
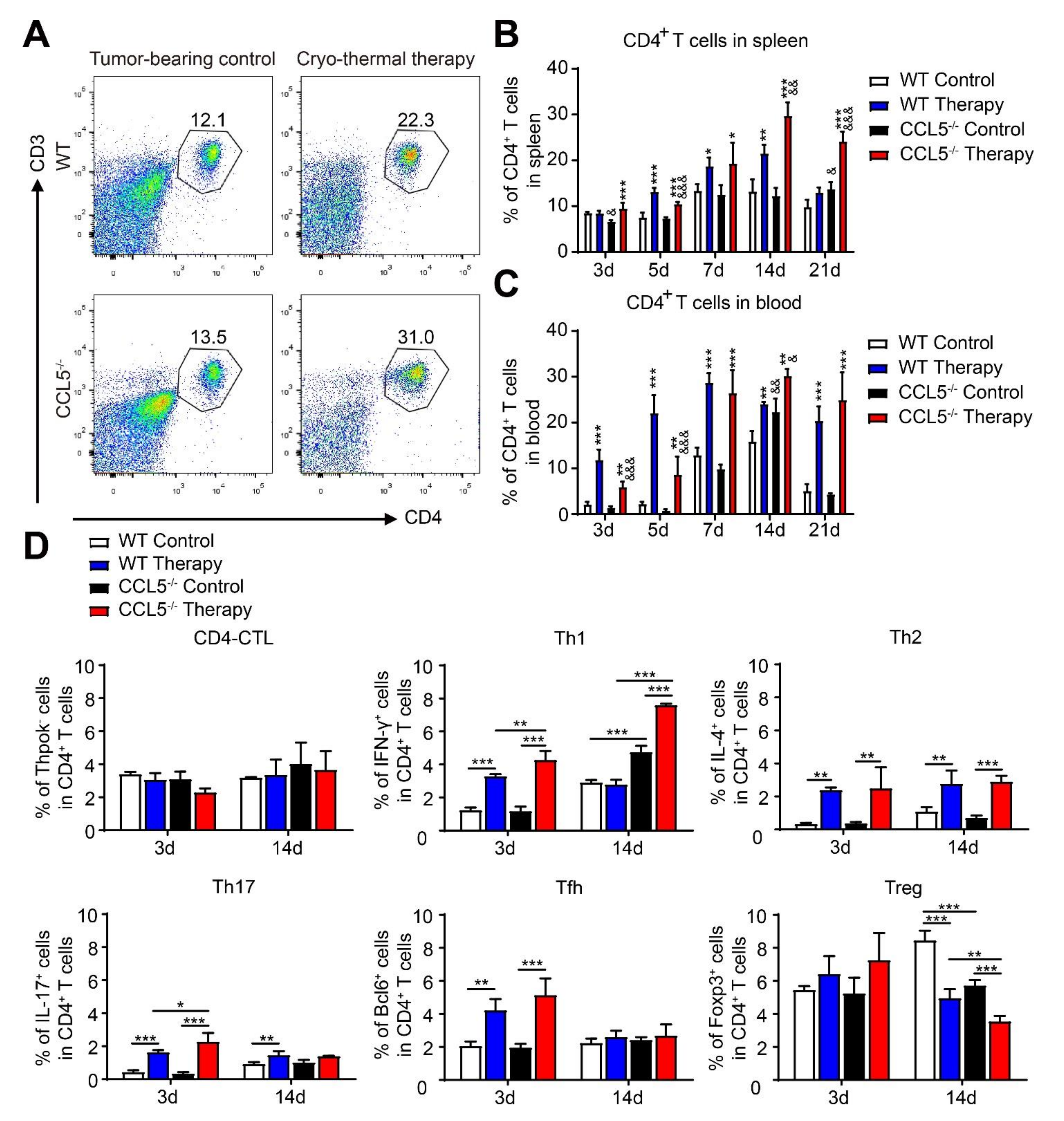
3.5. CCL5 Deficiency Modulated the Poly-Functional Differentiation of CD4+ T Cells after Cryo–Thermal Therapy
3.6. CCL5 Deficiency Promoted the Generation of Cryo–Thermal Therapy-Induced Cytotoxic CD8+ T Cells
3.7. CCL5-Deficiency-Enhancing, Cryo–Thermal-Induced M1 Macrophage Was Required for Inhibiting the Differentiation of CD4+ T Cell into Suppressive Subsets In Vitro
3.8. CCL5-Deficiency-Enhancing, Cryo–Thermal-Induced M1 Macrophage Polarization Promoted the Proliferation of CD8+ T Cells and Enhanced the Cytotoxicity of CD8+ T Cells In Vitro
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, Y.; Cao, X. Immunosuppressive cells in tumor immune escape and metastasis. J. Mol. Med. 2016, 94, 509–522. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quail, D.F.; Joyce, J.A. Microenvironmental regulation of tumor progression and metastasis. Nat. Med. 2013, 19, 1423–1437. [Google Scholar] [CrossRef]
- Kitamura, T.; Qian, B.Z.; Pollard, J.W. Immune cell promotion of metastasis. Nat. Rev. Immunol. 2015, 15, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Xu, H.; Wang, S. Immunosuppressive Role of Myeloid-Derived Suppressor Cells and Therapeutic Targeting in Lung Cancer. J. Immunol. Res. 2018, 2018, 6319649. [Google Scholar] [CrossRef]
- Rabinovich, G.A.; Gabrilovich, D.; Sotomayor, E.M. Immunosuppressive strategies that are mediated by tumor cells. Annu. Rev. Immunol. 2007, 25, 267–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becker, J.C.; Andersen, M.H.; Schrama, D.; Thor, S.P. Immune-suppressive properties of the tumor microenvironment. Cancer Immunol. Immunother. 2013, 62, 1137–1148. [Google Scholar] [CrossRef]
- Khalid, A.; Wolfram, J.; Ferrari, I.; Mu, C.; Mai, J.; Yang, Z.; Zhao, Y.; Ferrari, M.; Ma, X.; Shen, H. Recent Advances in Discovering the Role of CCL5 in Metastatic Breast Cancer. Mini Rev. Med. Chem. 2015, 15, 1063–1072. [Google Scholar] [CrossRef] [Green Version]
- Gao, D.; Rahbar, R.; Fish, E.N. CCL5 activation of CCR5 regulates cell metabolism to enhance proliferation of breast cancer cells. Open Biol. 2016, 6, 160122. [Google Scholar] [CrossRef] [Green Version]
- Aldinucci, D.; Casagrande, N. Inhibition of the CCL5/CCR5 Axis against the Progression of Gastric Cancer. Int. J. Mol. Sci. 2018, 19, 1477. [Google Scholar] [CrossRef] [Green Version]
- Sax, M.J.; Gasch, C.; Athota, V.R.; Freeman, R.; Rasighaemi, P.; Westcott, D.E.; Day, C.; Nikolic, I.; Elsworth, B.; Wei, M.; et al. Cancer cell CCL5 mediates bone marrow independent angiogenesis in breast cancer. Oncotarget 2016, 7, 85437–85449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Lang, M.; Zhao, T.; Feng, X.; Zheng, C.; Huang, C.; Hao, J.; Dong, J.; Luo, L.; Li, X.; et al. Cancer-FOXP3 directly activated CCL5 to recruit FOXP3(+)Treg cells in pancreatic ductal adenocarcinoma. Oncogene 2017, 36, 3048–3058. [Google Scholar] [CrossRef] [PubMed]
- Datar, I.; Qiu, X.; Ma, H.Z.; Yeung, M.; Aras, S.; De La Serna, I.; Al-Mulla, F.; Thiery, J.P.; Trumbly, R.; Fan, X.; et al. RKIP regulates CCL5 expression to inhibit breast cancer invasion and metastasis by controlling macrophage infiltration. Oncotarget 2015, 6, 39050–39061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, C.L.; Suri, R.M.; Rahdon, R.A.; Austyn, J.M.; Roake, J.A. Dendritic cell chemotaxis and transendothelial migration are induced by distinct chemokines and are regulated on maturation. Eur. J. Immunol. 1998, 28, 4114–4122. [Google Scholar] [CrossRef]
- Ban, Y.; Mai, J.; Li, X.; Mitchell-Flack, M.; Zhang, T.; Zhang, L.; Chouchane, L.; Ferrari, M.; Shen, H.; Ma, X. Targeting Autocrine CCL5-CCR5 Axis Reprograms Immunosuppressive Myeloid Cells and Reinvigorates Antitumor Immunity. Cancer Res. 2017, 77, 2857–2868. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Qin, J.; Zhong, L.; Gong, L.; Zhang, B.; Zhang, Y.; Gao, W.Q. CCL5-Mediated Th2 Immune Polarization Promotes Metastasis in Luminal Breast Cancer. Cancer Res. 2015, 75, 4312–4321. [Google Scholar] [CrossRef] [Green Version]
- Aldinucci, D.; Colombatti, A. The inflammatory chemokine CCL5 and cancer progression. Mediat. Inflamm. 2014, 2014, 292376. [Google Scholar] [CrossRef] [Green Version]
- Bai, J.F.; Liu, P.; Xu, L.X. Recent advances in thermal treatment techniques and thermally induced immune responses against cancer. IEEE Trans. Biomed. Eng. 2014, 61, 1497–1505. [Google Scholar] [CrossRef]
- Zhu, J.; Zhang, Y.; Zhang, A.; He, K.; Liu, P.; Xu, L.X. Cryo-thermal therapy elicits potent anti-tumor immunity by inducing extracellular Hsp70-dependent MDSC differentiation. Sci. Rep. 2016, 6, 27136. [Google Scholar] [CrossRef]
- Liu, P.; Ren, X.; Xu, L.X. Alternate cooling and heating thermal physical treatment: An effective strategy against mdscs in 4T1 mouse mammary carcinoma. In Proceedings of the Abstracts of the ASME 2012 Summer Bioengineering Conference, Fajardo, PR, USA, 20–23 June 2012; Volume 2012, pp. 937–938. [Google Scholar]
- Sun, J.; Zhang, A.; Xu, L.X. Evaluation of alternate cooling and heating for tumor treatment. Int. J. Heat Mass. Transf. 2008, 51, 5478–5485. [Google Scholar] [CrossRef]
- Xue, T.; Liu, P.; Zhou, Y.; Liu, K.; Yang, L.; Moritz, R.L.; Yan, W.; Xu, L.X. Interleukin-6 Induced “Acute” Phenotypic Microenvironment Promotes Th1 Anti-Tumor Immunity in Cryo-Thermal Therapy Revealed By Shotgun and Parallel Reaction Monitoring Proteomics. Theranostics 2016, 6, 773–794. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Liu, P.; Xu, L.X. The cryo-thermal therapy eradicated melanoma in mice by eliciting CD4(+) T-cell-mediated antitumor memory immune response. Cell Death Dis. 2017, 8, e2703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, K.; Jia, S.; Lou, Y.; Liu, P.; Xu, L.X. Cryo-thermal therapy induces macrophage polarization for durable anti-tumor immunity. Cell Death Dis. 2019, 10, 216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aldinucci, D.; Borghese, C.; Casagrande, N. The CCL5/CCR5 Axis in Cancer Progression. Cancers 2020, 10, 1765. [Google Scholar] [CrossRef]
- Zhang, S.; Zhong, M.; Wang, C.; Xu, Y.; Gao, W.Q.; Zhang, Y. CCL5-deficiency enhances intratumoral infiltration of CD8+ T cells in colorectal cancer. Cell Death Dis. 2018, 10, 766. [Google Scholar] [CrossRef] [Green Version]
- Cougoule, C.; Lastrucci, C.; Guiet, R.; Mascarau, R.; Meunier, E.; Lugo-Villarino, G.; Neyrolles, O.; Poincloux, R.; Maridonneau-Parini, I. Podosomes, But Not the Maturation Status, Determine the Protease-Dependent 3D Migration in Human Dendritic Cells. Front. Immunol. 2018, 9, 846. [Google Scholar] [CrossRef] [Green Version]
- Qian, B.Z.; Pollard, J.W. Macrophage diversity enhances tumor progression and metastasis. Cell 2010, 141, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Aswad, M.; Assi, S.; Schif-Zuck, S.; Ariel, A. CCL5 Promotes Resolution-Phase Macrophage Reprogramming in Concert with the Atypical Chemokine Receptor D6 and Apoptotic Polymorphonuclear Cells. J. Immunol. 2017, 199, 1393–1404. [Google Scholar] [CrossRef]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage plasticity, polarization, and function in health and disease. J. Cell Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef]
- Ku, A.W.; Muhitch, J.; Powers, C.A.; Diehl, M.; Kim, M.; Fisher, D.T.; Sharda, A.P.; Clements, V.K.; O’Loughlin, K.; Minderman, H.; et al. Tumor-induced MDSC act via remote control to inhibit L-selectin-dependent adaptive immunity in lymph nodes. Elife 2016, 5, e17375. [Google Scholar] [CrossRef]
- Chang, L.Y.; Lin, Y.C.; Mahalingam, J.; Huang, C.T.; Chen, T.W.; Kang, C.W.; Peng, H.M.; Chu, Y.Y.; Chiang, J.M.; Dutta, A.; et al. Tumor-derived chemokine CCL5 enhances TGF-beta-mediated killing of CD8(+) T cells in colon cancer by T-regulatory cells. Cancer Res. 2012, 72, 1092–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simson, L.; Ellyard, J.I.; Dent, L.A.; Matthaei, K.I.; Rothenberg, M.E.; Foster, P.S.; Smyth, M.J.; Parish, C. Regulation of carcinogenesis by IL-5 and CCL11: A potential role for eosinophils in tumor immune surveillance. J. Immunol. 2007, 178, 4222–4229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pozzi, L.A.M.; Maciaszek, J.W.; Rock, K.L. Both Dendritic Cells and Macrophages Can Stimulate Naive CD8 T Cells In Vivo to Proliferate, Develop Effector Function, and Differentiate into Memory Cells. J. Immunol. 2005, 175, 2071–2081. [Google Scholar] [CrossRef] [PubMed]
- Abe, H.; Wada, H.; Baghdadi, M.; Nakanishi, S.; Usui, Y.; Tsuchikawa, T.; Shichinohe, T.; Hirano, S.; Seino, K.-I. Identification of a highly immunogenic mouse breast cancer sub cell line, 4T1-S. Hum. Cell 2016, 29, 58–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bosiljcic, M.; Cederberg, R.A.; Hamilton, M.J.; LePard, N.E.; Harbourne, B.T.; Collier, J.L.; Halvorsen, E.C.; Shi, R.; Franks, S.E.; Kim, A.Y.; et al. Targeting myeloid-derived suppressor cells in combination with primary mammary tumor resection reduces metastatic growth in the lungs. Breast Cancer Res. 2019, 21, 103. [Google Scholar] [CrossRef]
- Ravindranathan, S.; Nguyen, K.G.; Kurtz, S.L.; Frazier, H.N.; Smith, S.G.; Koppolu, B.P.; Rajaram, N.; Zaharoff, D.A. Tumor-derived granulocyte colony-stimulating factor diminishes efficacy of breast tumor cell vaccines. Breast Cancer Res. 2018, 20, 126. [Google Scholar] [CrossRef]
- Steenbrugge, J.; Vander Elst, N.; Demeyere, K.; De Wever, O.; Sanders, N.N.; Van Den Broeck, W.; Dirix, L.; Van Laere, S.; Meyer, E. Comparative Profiling of Metastatic 4T1- vs. Non-metastatic Py230-Based Mammary Tumors in an Intraductal Model for Triple-Negative Breast Cancer. Front. Immunol. 2019, 10, 2928. [Google Scholar] [CrossRef]
- Blattner, C.; Fleming, V.; Weber, R.; Himmelhan, B.; Altevogt, P.; Gebhardt, C.; Schulze, T.J.; Razon, H.; Hawila, E.; Wildbaum, G.; et al. CCR5+ Myeloid-Derived Suppressor Cells Are Enriched and Activated in Melanoma Lesions. Cancer Res. 2018, 78, 157–167. [Google Scholar] [CrossRef] [Green Version]
- Nie, Y.; Huang, H.; Guo, M.; Chen, J.; Wu, W.; Li, W.; Xu, X.; Lin, X.; Fu, W.; Yao, Y.; et al. Breast Phyllodes Tumors Recruit and Repolarize Tumor-Associated Macrophages via Secreting CCL5 to Promote Malignant Progression, Which Can Be Inhibited by CCR5 Inhibition Therapy. Clin. Cancer Res. 2019, 25, 3873–3886. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Lv, D.; Kim, H.-J.; Kurt, R.A.; Bu, W.; Li, Y.; Ma, X. A novel role of hematopoietic CCL5 in promoting triple-negative mammary tumor progression by regulating generation of myeloid-derived suppressor cells. Cell Res. 2013, 23, 394–408. [Google Scholar] [CrossRef]
- Jayasinghe, M.M.; Golden, J.M.; Nair, P.; O’Donnell, C.M.; Werner, M.T.; Kurt, R.A. Tumor-derived CCL5 does not contribute to breast cancer progression. Breast Cancer Res. Treat. 2008, 111, 511–521. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Jia, S.; Lou, Y.; He, K.; Xu, L.X. Cryo-thermal therapy inducing MI macrophage polarization created CXCL10 and IL-6-rich pro-inflammatory environment for CD4+ T cell-mediated anti-tumor immunity. Int. J. Hyperthermia 2019, 36, 408–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dogan, R.-N.E.; Long, N.; Forde, E.; Dennis, K.; Kohm, A.P.; Miller, S.D.; Karpus, W.J. CCL22 regulates experimental autoimmune encephalomyelitis by controlling inflammatory macrophage accumulation and effector function. J. Leukoc. Biol. 2011, 89, 93–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, Y.-M.; Hsu, S.-C.; Zhang, J.; Zhou, Y.; Plunkett, B.; Huang, S.-K.; Gao, P.-S. Functional interaction of cockroach allergens and mannose receptor (CD206) in human circulating fibrocytes. PLoS ONE 2013, 8, e64105. [Google Scholar] [CrossRef] [Green Version]
- Ushio, A.; Arakaki, R.; Otsuka, K.; Yamada, A.; Tsunematsu, T.; Kudo, Y.; Aota, K.; Azuma, M.; Ishimaru, N. CCL22-Producing Resident Macrophages Enhance T Cell Response in Sjögren’s Syndrome. Front. Immunol. 2018, 9, 2594. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fluorescence Labeling | Antibodies | Clone |
|---|---|---|
| FITC | CD3 | 145-2C11 |
| CD11b | M1/70 | |
| CD11c | N418 | |
| PE | Gr-1 | R86-8C5 |
| IL-17 | TC11-18H10.1 | |
| Perforin | S16001B | |
| Foxp3 | MF-14 | |
| CD86 2 | GL-1 | |
| PerCP/Cy5.5 | I-A/I-E | M5/114.15.2 |
| Bcl-6 | 7D1 | |
| PE/Cy7 | CD25 | 3C7 |
| APC | F4/80 | BM8 |
| AF647 | ThPok 1 | T43-94 |
| APC/Cy7 | CD4 | RM4.5 |
| CD8 | 53-5.8 | |
| BV421 | IL-4 | 11B11 |
| BV510 | IFN-γ | XMG1.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lou, Y.; Jia, S.; Liu, P.; Xu, L.X. CCL5 Deficiency Enhanced Cryo–Thermal-Triggered Long-Term Anti-Tumor Immunity in 4T1 Murine Breast Cancer. Biomedicines 2022, 10, 559. https://doi.org/10.3390/biomedicines10030559
Lou Y, Jia S, Liu P, Xu LX. CCL5 Deficiency Enhanced Cryo–Thermal-Triggered Long-Term Anti-Tumor Immunity in 4T1 Murine Breast Cancer. Biomedicines. 2022; 10(3):559. https://doi.org/10.3390/biomedicines10030559
Chicago/Turabian StyleLou, Yue, Shengguo Jia, Ping Liu, and Lisa X. Xu. 2022. "CCL5 Deficiency Enhanced Cryo–Thermal-Triggered Long-Term Anti-Tumor Immunity in 4T1 Murine Breast Cancer" Biomedicines 10, no. 3: 559. https://doi.org/10.3390/biomedicines10030559