
Proteasome-Mediated Regulation of GATA2 Expression and Androgen Receptor Transcription in Benign Prostate Epithelial Cells

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Cell Cultures
2.2. Vectors, Lentiviral Particles, Transfection and Transduction
2.3. RNA Interference
2.4. Western Blotting
2.5. Real-Time (RT) Reverse Transcription Quantitative PCR (qPCR)
2.6. Agilent Microarrays
2.7. RNA Sequencing (RNA-seq)
2.8. Statistical Analysis
3. Results
3.1. Expression of GATA Factors in Prostate Cell Lines
3.2. Exogenous GATA2 Was Degraded in TA Cells
3.3. Proteasome-Mediated Degradation of GATA2 in TA Cells
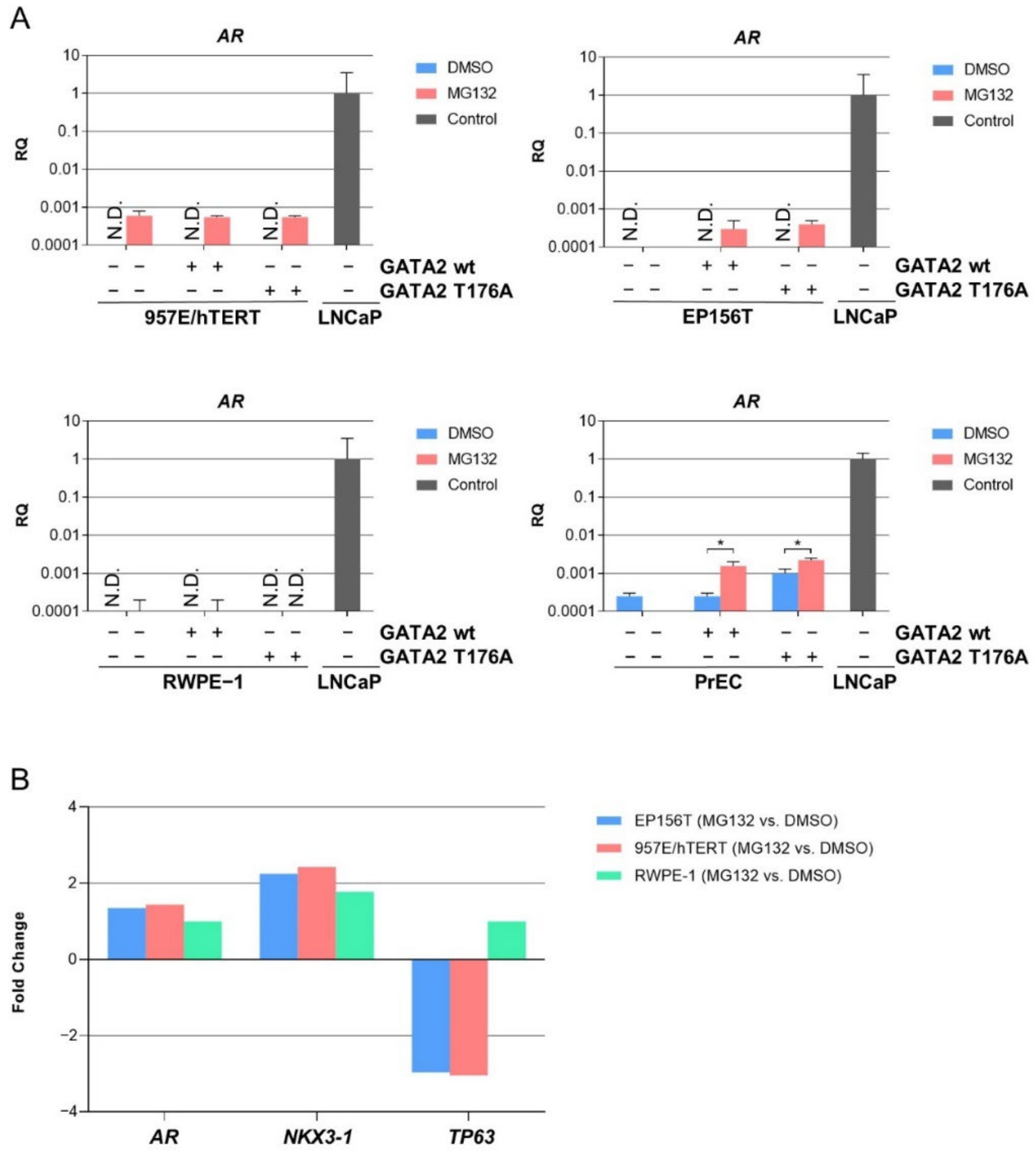
3.4. Proteasome-Mediated Expression of AR mRNA in TA Cells
3.5. Exogenous GATA2 Induces Luminal Marker Genes in PrEC Cells
3.6. Reduction in Endogenous FBXW7 Stabilized Exogenous GATA2 Protein
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Packer, J.R.; Maitland, N.J. The molecular and cellular origin of human prostate cancer. Biochim. Biophys. Acta 2016, 1863, 1238–1260. [Google Scholar] [CrossRef]
- Ke, X.S.; Li, W.C.; Hovland, R.; Qu, Y.; Liu, R.H.; McCormack, E.; Thorsen, F.; Olsen, J.R.; Molven, A.; Kogan-Sakin, I.; et al. Reprogramming of cell junction modules during stepwise epithelial to mesenchymal transition and accumulation of malignant features in vitro in a prostate cell model. Exp. Cell Res. 2011, 317, 234–247. [Google Scholar] [CrossRef] [PubMed]
- Ke, X.S.; Qu, Y.; Goldfinger, N.; Rostad, K.; Hovland, R.; Akslen, L.A.; Rotter, V.; Øyan, A.M.; Kalland, K.H. Epithelial to mesenchymal transition of a primary prostate cell line with switches of cell adhesion modules but without malignant transformation. PLoS ONE 2008, 3, e3368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, X.S.; Qu, Y.; Rostad, K.; Li, W.C.; Lin, B.; Halvorsen, O.J.; Haukaas, S.A.; Jonassen, I.; Petersen, K.; Goldfinger, N.; et al. Genome-wide profiling of histone h3 lysine 4 and lysine 27 trimethylation reveals an epigenetic signature in prostate carcinogenesis. PLoS ONE 2009, 4, e4687. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, Y.; Oyan, A.M.; Liu, R.; Hua, Y.; Zhang, J.; Hovland, R.; Popa, M.; Liu, X.; Brokstad, K.A.; Simon, R.; et al. Generation of prostate tumor-initiating cells is associated with elevation of reactive oxygen species and IL-6/STAT3 signaling. Cancer Res. 2013, 73, 7090–7100. [Google Scholar] [CrossRef] [Green Version]
- Kogan, I.; Goldfinger, N.; Milyavsky, M.; Cohen, M.; Shats, I.; Dobler, G.; Klocker, H.; Wasylyk, B.; Voller, M.; Aalders, T.; et al. hTERT-immortalized prostate epithelial and stromal-derived cells: An authentic in vitro model for differentiation and carcinogenesis. Cancer Res. 2006, 66, 3531–3540. [Google Scholar] [CrossRef] [Green Version]
- Azeem, W.; Hellem, M.R.; Olsen, J.R.; Hua, Y.; Marvyin, K.; Qu, Y.; Lin, B.; Ke, X.; Oyan, A.M.; Kalland, K.H. An androgen response element driven reporter assay for the detection of androgen receptor activity in prostate cells. PLoS ONE 2017, 12, e0177861. [Google Scholar] [CrossRef] [Green Version]
- Olsen, J.R.; Azeem, W.; Hellem, M.R.; Marvyin, K.; Hua, Y.; Qu, Y.; Li, L.; Lin, B.; Ke, X.; Oyan, A.M.; et al. Context dependent regulatory patterns of the androgen receptor and androgen receptor target genes. BMC Cancer 2016, 16, 377. [Google Scholar] [CrossRef] [Green Version]
- Antony, L.; van der Schoor, F.; Dalrymple, S.L.; Isaacs, J.T. Androgen receptor (AR) suppresses normal human prostate epithelial cell proliferation via AR/beta-catenin/TCF-4 complex inhibition of c-MYC transcription. Prostate 2014, 74, 1118–1131. [Google Scholar] [CrossRef] [Green Version]
- Vander Griend, D.J.; Litvinov, I.V.; Isaacs, J.T. Conversion of Androgen Receptor Signaling From a Growth Suppressor in Normal Prostate Epithelial Cells to an Oncogene in Prostate Cancer Cells Involves a Gain of Function in c-Myc Regulation. Int. J. Biol. Sci. 2014, 10, 627–642. [Google Scholar] [CrossRef] [Green Version]
- Mills, I.G. Maintaining and reprogramming genomic androgen receptor activity in prostate cancer. Nat. Rev. Cancer 2014, 14, 187–198. [Google Scholar] [CrossRef]
- Frank, S.B.; Miranti, C.K. Disruption of prostate epithelial differentiation pathways and prostate cancer development. Front. Oncol. 2013, 3, 273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bello, D.; Webber, M.M.; Kleinman, H.K.; Wartinger, D.D.; Rhim, J.S. Androgen responsive adult human prostatic epithelial cell lines immortalized by human papillomavirus 18. Carcinogenesis 1997, 18, 1215–1223. [Google Scholar] [CrossRef] [PubMed]
- Litvinov, I.V.; Vander Griend, D.J.; Xu, Y.; Antony, L.; Dalrymple, S.L.; Isaacs, J.T. Low-calcium serum-free defined medium selects for growth of normal prostatic epithelial stem cells. Cancer Res. 2006, 66, 8598–8607. [Google Scholar] [CrossRef] [Green Version]
- Yasunaga, Y.; Nakamura, K.; Ewing, C.M.; Isaacs, W.B.; Hukku, B.; Rhim, J.S. A novel human cell culture model for the study of familial prostate cancer. Cancer Res. 2001, 61, 5969–5973. [Google Scholar]
- Archer, L.K.; Frame, F.M.; Maitland, N.J. Stem cells and the role of ETS transcription factors in the differentiation hierarchy of normal and malignant prostate epithelium. J. Steroid Biochem. Mol. Biol. 2017, 166, 68–83. [Google Scholar] [CrossRef]
- Buhler, P.; Wolf, P.; Katzenwadel, A.; Schultze-Seemann, W.; Wetterauer, U.; Freudenberg, N.; Elsasser-Beile, U. Primary prostate cancer cultures are models for androgen-independent transit amplifying cells. Oncol. Rep. 2010, 23, 465–470. [Google Scholar]
- Petersen, K.; Oyan, A.M.; Rostad, K.; Olsen, S.; Bo, T.H.; Salvesen, H.B.; Gjertsen, B.T.; Bruserud, O.; Halvorsen, O.J.; Akslen, L.A.; et al. Comparison of nucleic acid targets prepared from total RNA or poly(A) RNA for DNA oligonucleotide microarray hybridization. Anal. Biochem. 2007, 366, 46–58. [Google Scholar] [CrossRef] [PubMed]
- Dysvik, B.; Jonassen, I. J-Express: Exploring gene expression data using Java. Bioinformatics 2001, 17, 369–370. [Google Scholar] [CrossRef]
- Dalrymple, S.; Antony, L.; Xu, Y.; Uzgare, A.R.; Arnold, J.T.; Savaugeot, J.; Sokoll, L.J.; De Marzo, A.M.; Isaacs, J.T. Role of notch-1 and E-cadherin in the differential response to calcium in culturing normal versus malignant prostate cells. Cancer Res. 2005, 65, 9269–9279. [Google Scholar] [CrossRef] [Green Version]
- Koga, S.; Yamaguchi, N.; Abe, T.; Minegishi, M.; Tsuchiya, S.; Yamamoto, M.; Minegishi, N. Cell-cycle-dependent oscillation of GATA2 expression in hematopoietic cells. Blood 2007, 109, 4200–4208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, B.; Lanz, R.B.; Fiskus, W.; Geng, C.; Yi, P.; Hartig, S.M.; Rajapakshe, K.; Shou, J.; Wei, L.; Shah, S.S.; et al. GATA2 facilitates steroid receptor coactivator recruitment to the androgen receptor complex. Proc. Natl. Acad. Sci. USA 2014, 111, 18261–18266. [Google Scholar] [CrossRef] [Green Version]
- Minegishi, N.; Suzuki, N.; Kawatani, Y.; Shimizu, R.; Yamamoto, M. Rapid turnover of GATA-2 via ubiquitin-proteasome protein degradation pathway. Genes Cells 2005, 10, 693–704. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, T.; Kitagawa, K.; Ohhata, T.; Sakai, S.; Uchida, C.; Shibata, K.; Minegishi, N.; Yumimoto, K.; Nakayama, K.I.; Masumoto, K.; et al. Regulation of GATA-binding protein 2 levels via ubiquitin-dependent degradation by Fbw7: Involvement of cyclin B-cyclin-dependent kinase 1-mediated phosphorylation of THR176 in GATA-binding protein 2. J. Biol. Chem. 2015, 290, 10368–10381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, J.; Wang, C.; Deng, Y.; Yu, L.; Huang, H. Destruction of full-length androgen receptor by wild-type SPOP, but not prostate-cancer-associated mutants. Cell Rep. 2014, 6, 657–669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heer, R.; Robson, C.N.; Shenton, B.K.; Leung, H.Y. The role of androgen in determining differentiation and regulation of androgen receptor expression in the human prostatic epithelium transient amplifying population. J. Cell. Physiol. 2007, 212, 572–578. [Google Scholar] [CrossRef] [PubMed]
- Rane, J.K.; Droop, A.P.; Maitland, N.J. A Detailed Analysis of Gene Expression in Human Basal, Luminal, and Stromal Cell Populations from Benign Prostatic Hyperplasia Tissues and Comparisons with Cultured Basal Cells. Eur. Urol. 2017, 72, 157–159. [Google Scholar] [CrossRef] [Green Version]
- Rane, J.K.; Droop, A.P.; Pellacani, D.; Polson, E.S.; Simms, M.S.; Collins, A.T.; Caves, L.S.; Maitland, N.J. Conserved two-step regulatory mechanism of human epithelial differentiation. Stem Cell Rep. 2014, 2, 180–188. [Google Scholar] [CrossRef] [Green Version]
- Gao, N.; Zhang, J.; Rao, M.A.; Case, T.C.; Mirosevich, J.; Wang, Y.; Jin, R.; Gupta, A.; Rennie, P.S.; Matusik, R.J. The role of hepatocyte nuclear factor-3 alpha (Forkhead Box A1) and androgen receptor in transcriptional regulation of prostatic genes. Mol. Endocrinol. 2003, 17, 1484–1507. [Google Scholar] [CrossRef] [Green Version]
- Perez-Stable, C.M.; Pozas, A.; Roos, B.A. A role for GATA transcription factors in the androgen regulation of the prostate-specific antigen gene enhancer. Mol. Cell. Endocrinol. 2000, 167, 43–53. [Google Scholar] [CrossRef]
- Dutta, A.; Le Magnen, C.; Mitrofanova, A.; Ouyang, X.; Califano, A.; Abate-Shen, C. Identification of an NKX3.1-G9a-UTY transcriptional regulatory network that controls prostate differentiation. Science 2016, 352, 1576–1580. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| EP156T | EPT3-M1 | LNCaP | 22Rv1 | VCaP | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Control | R1881 (10 nM) | Control | R1881 (10 nM) | Control | R1881 (10 nM) | Control | R1881 (1 nM) | Control | R1881 (1 nM) | |
| GATA1 | 0 | 0 | 0 | 0 | 0.1 | 0 | 0 | 0 | 0 | 0 |
| GATA2 isoform 1, transcript variant 1 | 0.0 | 0.2 | 0.4 | 0.5 | 6.4 | 3.6 | 0.9 | 0.2 | 4.5 | 11.7 |
| GATA2 isoform 1, transcript variant 2 | 0.2 | 0.0 | 0.1 | 0.0 | 14.5 | 3.6 | 2.5 | 1.6 | 20.2 | 10.0 |
| GATA2 isoform 2 | 0.0 | 0.0 | 0 | 0.0 | 2.1 | 0.3 | 0.7 | 0.1 | 3.9 | 5.5 |
| GATA3, transcript variant 1 | 2.6 | 1.3 | 0.3 | 0.3 | 0.0 | 0.0 | 1.9 | 0.2 | 0.0 | 0.0 |
| GATA3, transcript variant 2 | 1 | 1 | 0 | 0 | 0 | 0 | 0.3 | 0.5 | 0 | 0 |
| GATA4 | 0.0 | 0.0 | 0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 |
| GATA5 | 0.0 | 0.0 | 0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.2 | 0.1 |
| GATA6 | 1.4 | 1.1 | 2.7 | 2.9 | 0.6 | 0.6 | 0.8 | 1.2 | 1.4 | 2.1 |
| KLK3 | 0 | 0 | 0 | 0 | 35 | 799 | 4 | 9 | 5 | 52 |
| NKX3-1 | 0 | 0 | 4 | 4 | 40 | 139 | 34 | 40 | 171 | 319 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Azeem, W.; Olsen, J.R.; Hellem, M.R.; Hua, Y.; Marvyin, K.; Ke, X.; Øyan, A.M.; Kalland, K.-H. Proteasome-Mediated Regulation of GATA2 Expression and Androgen Receptor Transcription in Benign Prostate Epithelial Cells. Biomedicines 2022, 10, 473. https://doi.org/10.3390/biomedicines10020473
Azeem W, Olsen JR, Hellem MR, Hua Y, Marvyin K, Ke X, Øyan AM, Kalland K-H. Proteasome-Mediated Regulation of GATA2 Expression and Androgen Receptor Transcription in Benign Prostate Epithelial Cells. Biomedicines. 2022; 10(2):473. https://doi.org/10.3390/biomedicines10020473
Chicago/Turabian StyleAzeem, Waqas, Jan Roger Olsen, Margrete Reime Hellem, Yaping Hua, Kristo Marvyin, Xisong Ke, Anne Margrete Øyan, and Karl-Henning Kalland. 2022. "Proteasome-Mediated Regulation of GATA2 Expression and Androgen Receptor Transcription in Benign Prostate Epithelial Cells" Biomedicines 10, no. 2: 473. https://doi.org/10.3390/biomedicines10020473