

Re-Analysis of Single-Nucleus Transcriptomics Reveals Diverse Dorsal Root Ganglia Macrophage Responses Following Peripheral Nerve Injury

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
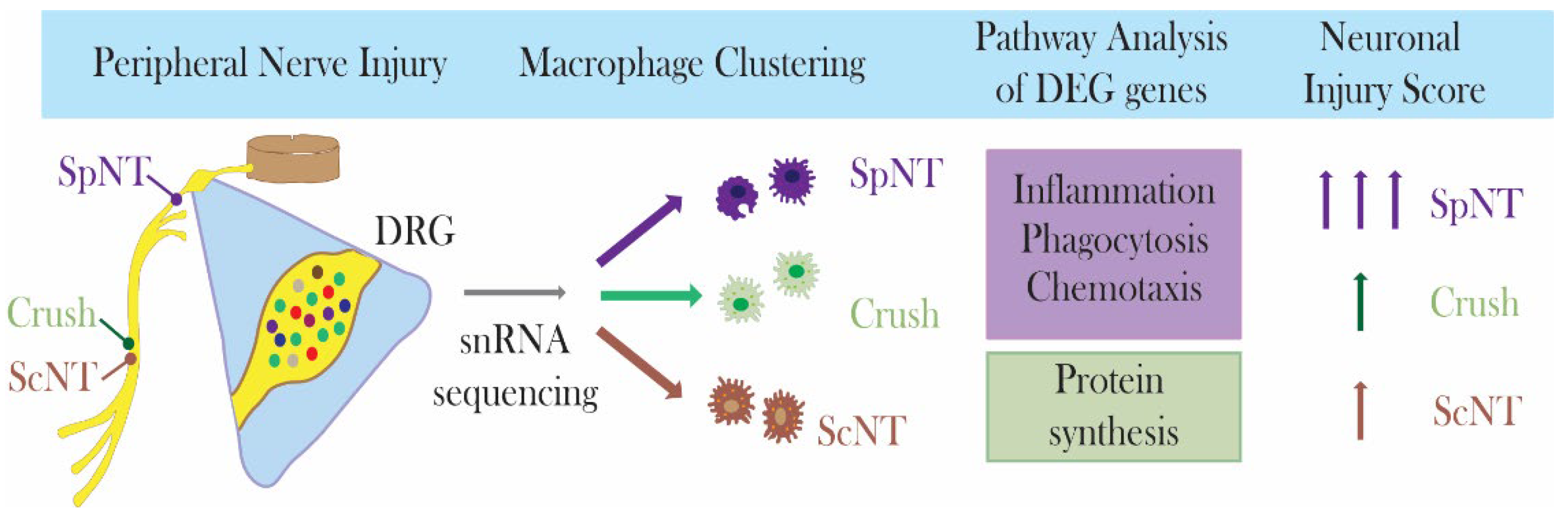
:1. Introduction
2. Materials and Methods
2.1. Single-Nucleus Transcriptomics
2.2. Preprocessing and Filtering
2.3. Clustering and Cluster Annotation
2.4. Gene Expression Scoring of Macrophage and Neuron Identities
2.5. Pseudobulk Analysis of Differentially Expressed Genes
2.6. Pathway Analysis
3. Results
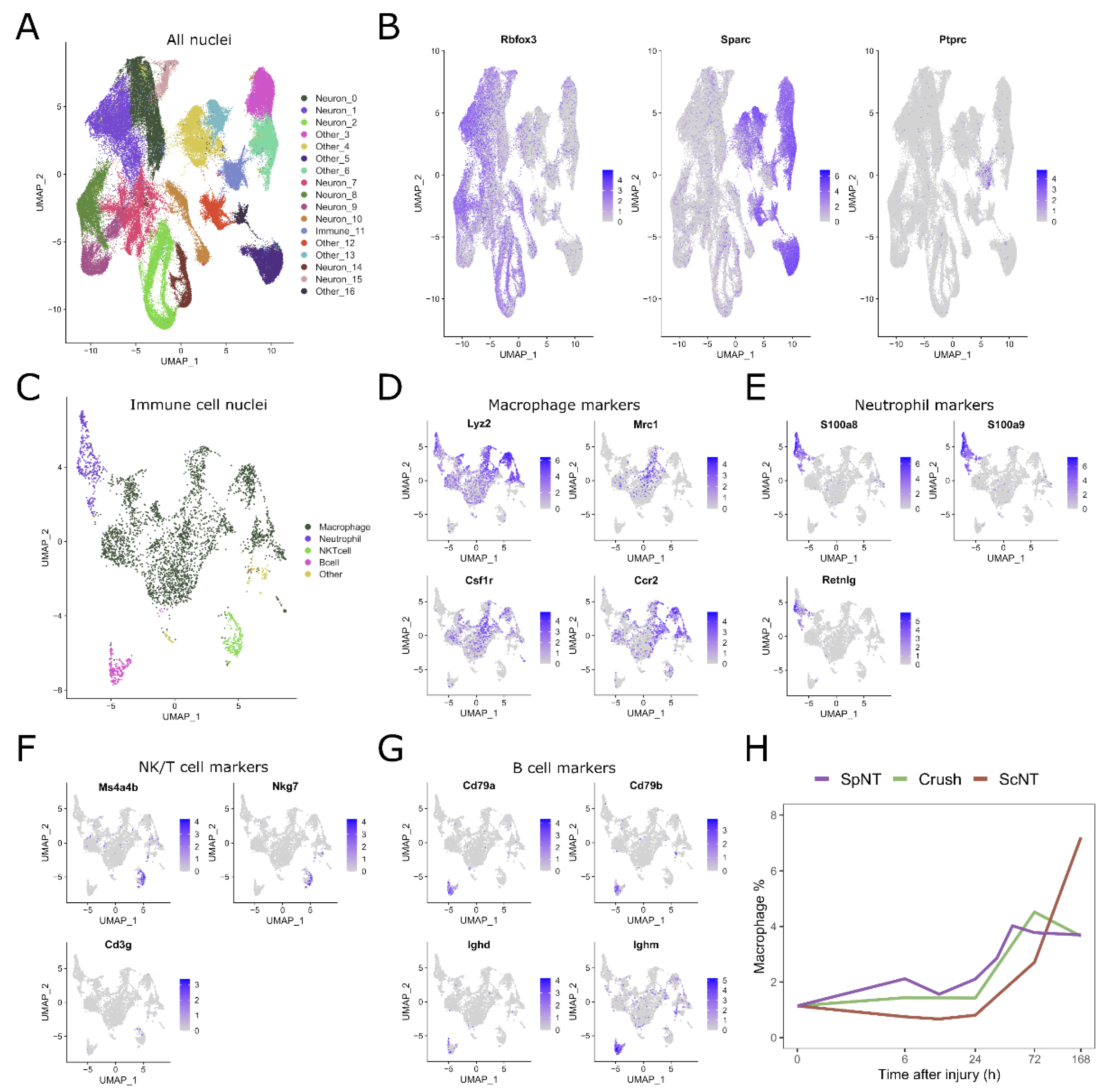
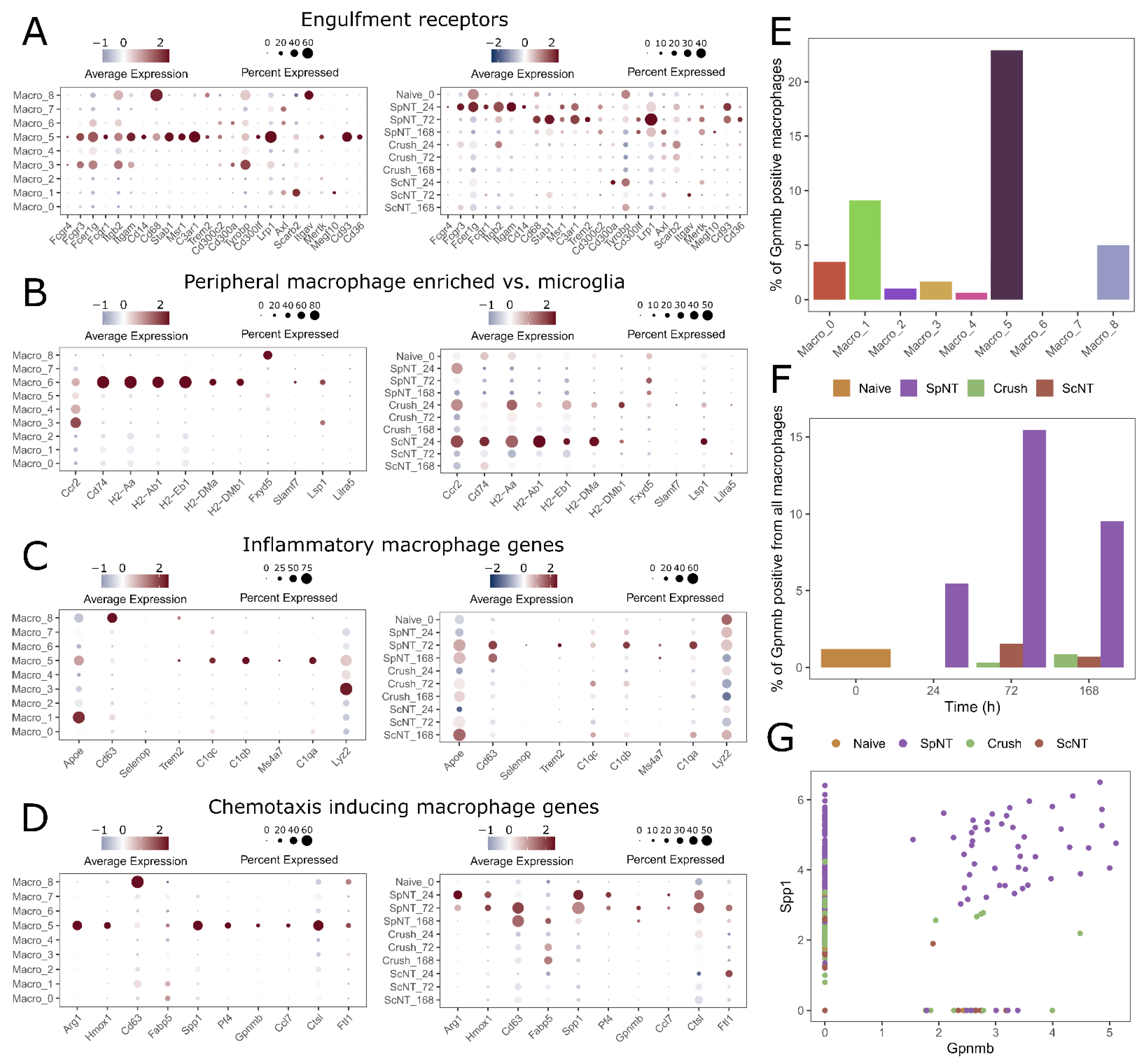
3.1. Peripheral Nerve Injuries Time-Dependently Increase the Abundance of Macrophages in the Mouse DRG
3.2. The Time Course of DRG Macrophage Responses to Peripheral Nerve Injuries Is Dependent on the Site and Type of Injury
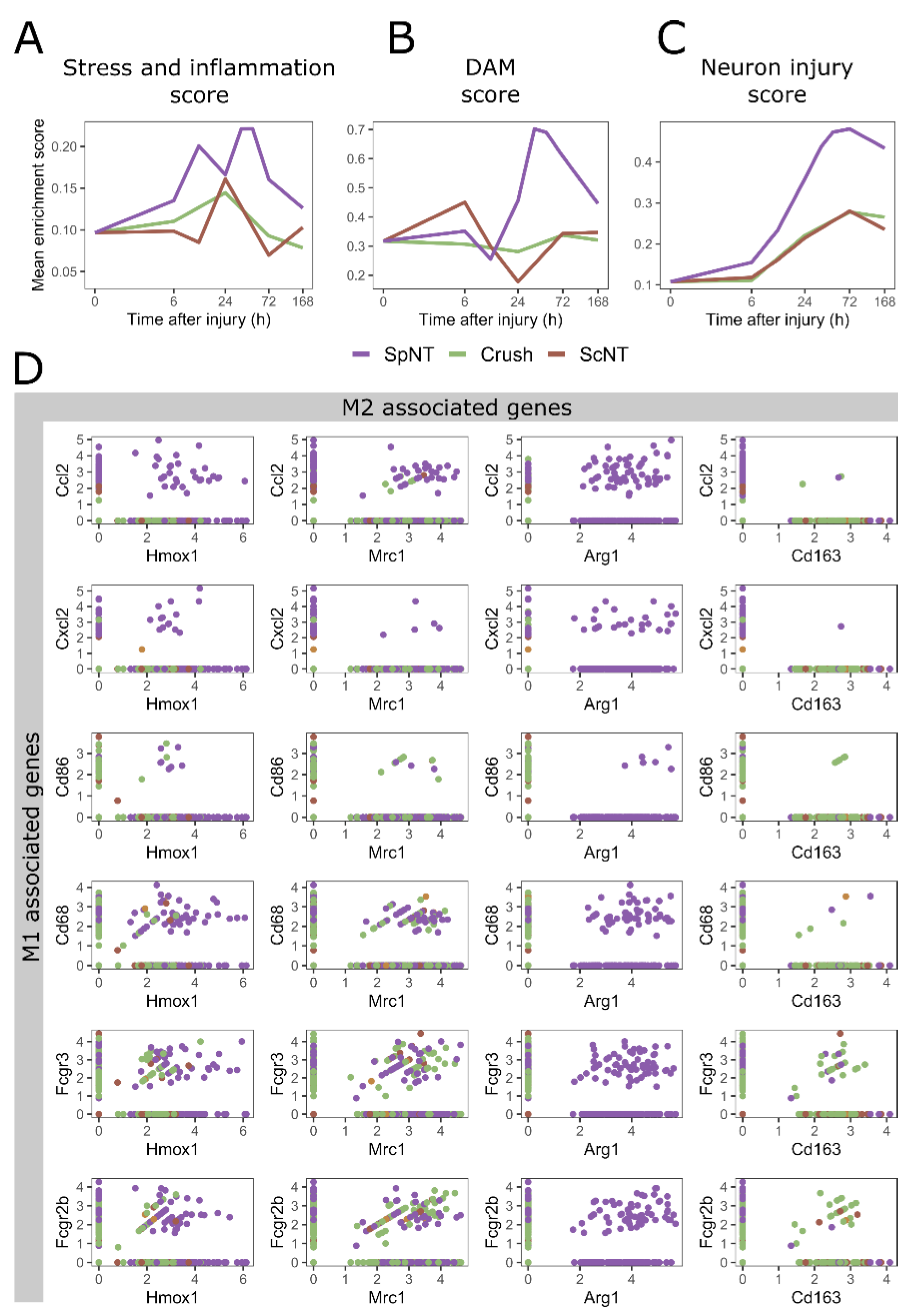
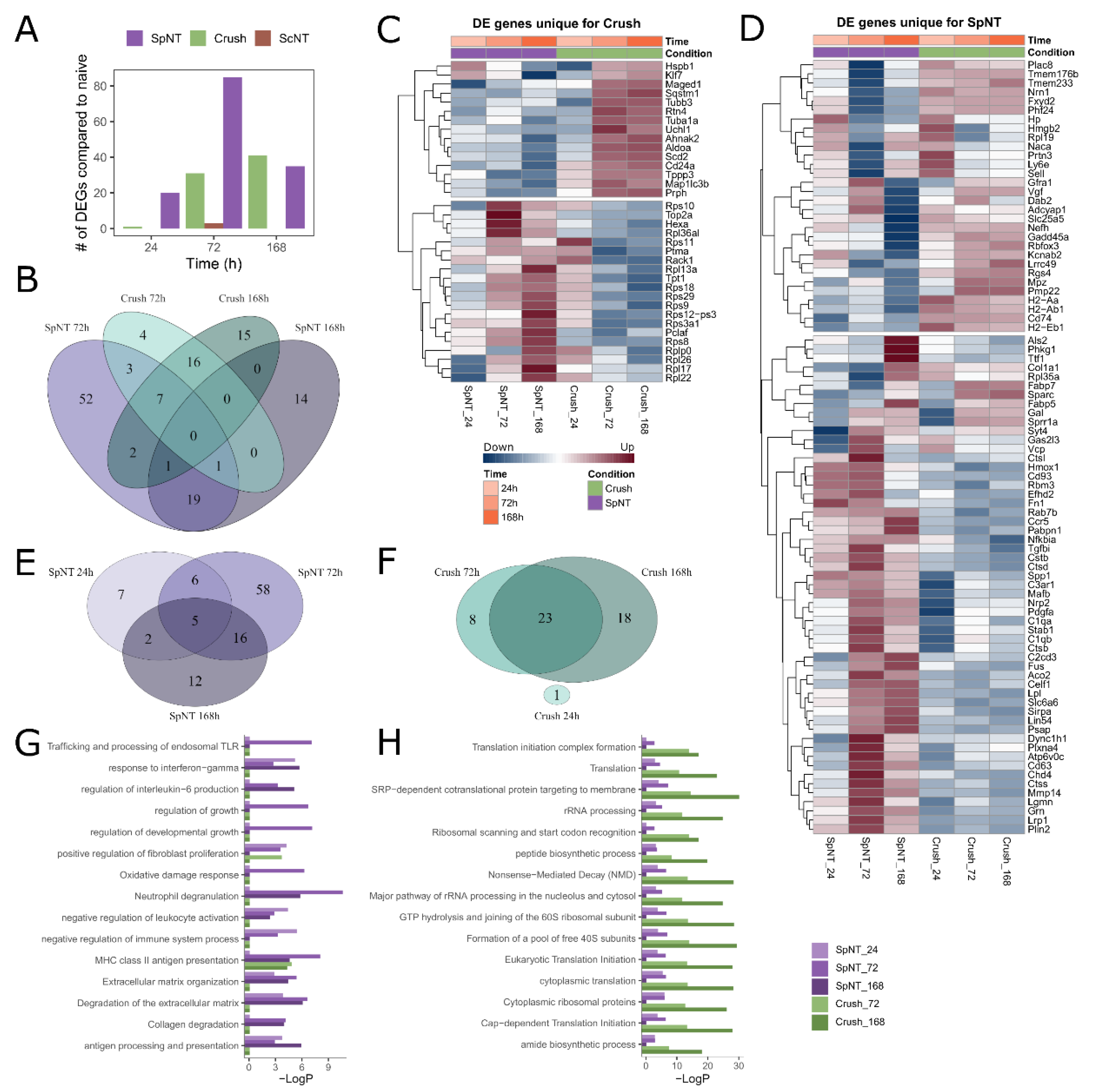
3.3. Spinal Nerve Transection Modulates Inflammatory Responses of Macrophages, While Sciatic Nerve Crush Affects Their Translational Capacity
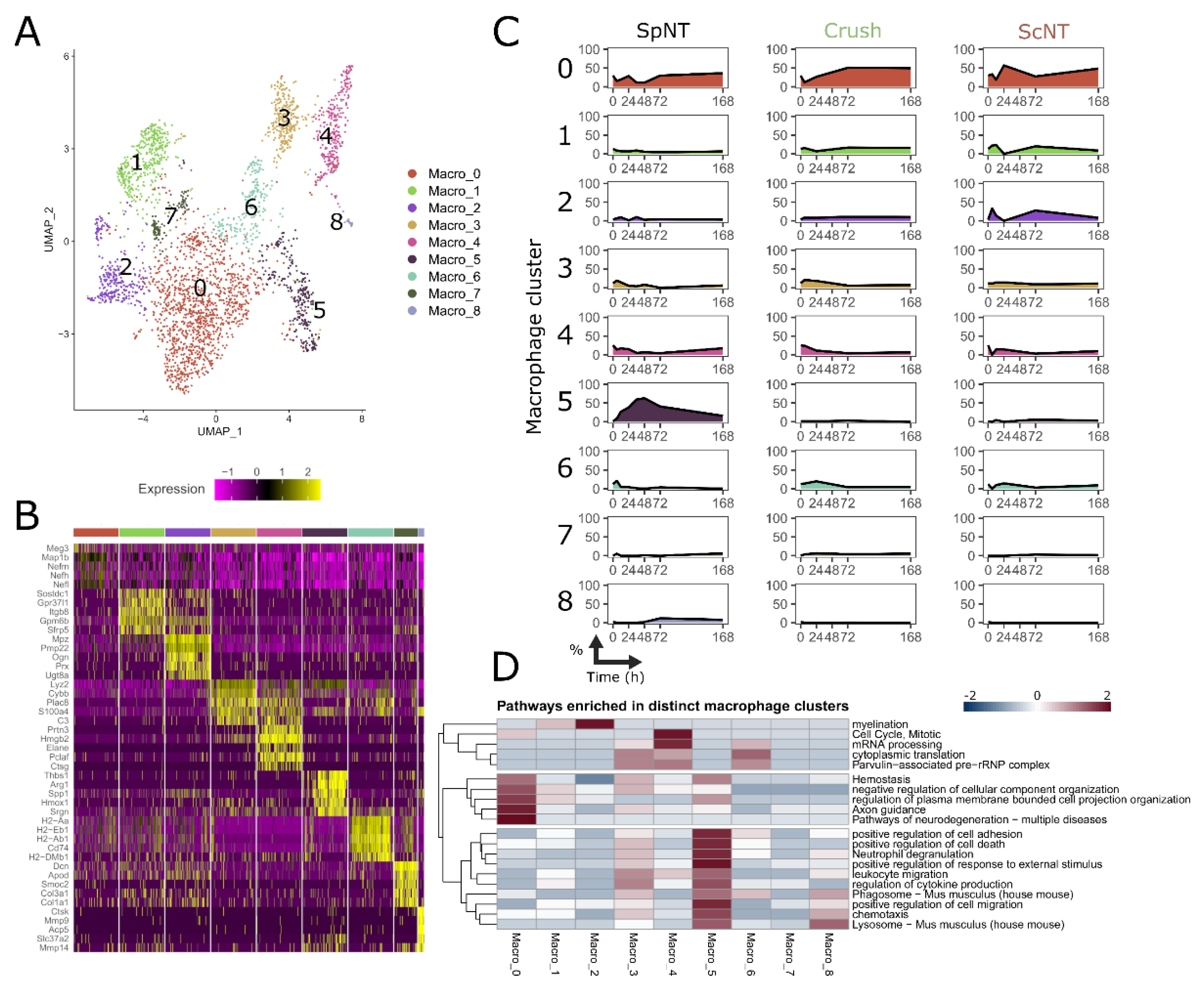
3.4. Spinal Nerve Transection Leads to the Appearance of a Particular Macrophage Cluster Associated with the Regulation of Inflammation
3.5. Nerve Injury Induces Transcriptomically Unique Macrophage Subpopulations in the DRG
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Tansley, S.; Uttam, S.; Guzmán, A.U.; Yaqubi, M.; Pacis, A.; Parisien, M.; Deamond, H.; Wong, C.; Rabau, O.; Brown, N.; et al. Single-cell RNA sequencing reveals time- and sex-specific responses of mouse spinal cord microglia to peripheral nerve injury and links ApoE to chronic pain. Nat. Commun. 2022, 13, 843. [Google Scholar] [CrossRef]
- Kalinski, A.L.; Yoon, C.; Huffman, L.D.; Duncker, P.C.; Kohen, R.; Passino, R.; Hafner, H.; Johnson, C.; Kawaguchi, R.; Carbajal, K.S.; et al. Analysis of the immune response to sciatic nerve injury identifies efferocytosis as a key mechanism of nerve debridement. Elife 2020, 9, e60223. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.; Liu, H.; Hamel, K.A.; Morvan, M.G.; Yu, S.; Leff, J.; Guan, Z.; Braz, J.M.; Basbaum, A.I. Dorsal root ganglion macrophages contribute to both the initiation and persistence of neuropathic pain. Nat. Commun. 2020, 11, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avraham, O.; Feng, R.; Ewan, E.E.; Rustenhoven, J.; Zhao, G.; Cavalli, V. Profiling sensory neuron microenvironment after peripheral and central axon injury reveals key pathways for neural repair. Elife 2021, 10, e68457. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.; Zhang, J.; Xu, J.; Fu, L.; Xu, Y.; Wang, X.; Li, Z.; Zhu, L.; Sun, H.; Zheng, H.; et al. SIRT6 inhibition delays peripheral nerve recovery by suppressing migration, phagocytosis and M2-polarization of macrophages. Cell Biosci. 2021, 11, 1–19. [Google Scholar] [CrossRef]
- Xu, J.; Wen, J.; Fu, L.; Liao, L.; Zou, Y.; Zhang, J.; Deng, J.; Zhang, H.; Liu, J.; Wang, X.; et al. Macrophage-specific RhoA knockout delays Wallerian degeneration after peripheral nerve injury in mice. J. Neuroinflamm. 2021, 18, 1–17. [Google Scholar] [CrossRef]
- Matson, K.J.E.; Russ, D.E.; Kathe, C.; Maric, D.; Hua, I.; Krynitsky, J.; Pursley, R.; Sathyamurthy, A.; Squair, J.W.; Courtine, J.; et al. A Single Cell Atlas of Spared Tissue Below a Spinal Cord Injury Reveals Cellular Mechanisms of Repair. BioRxiv 2021. [Google Scholar] [CrossRef]
- Milich, L.M.; Choi, J.S.; Ryan, C.; Cerqueira, S.R.; Benavides, S.; Yahn, S.L.; Tsoulfas, P.; Lee, J.K. Single-cell analysis of the cellular heterogeneity and interactions in the injured mouse spinal cord. J. Exp. Med. 2021, 218, e20210040. [Google Scholar] [CrossRef]
- Ydens, E.; Amann, L.; Asselbergh, B.; Scott, C.L.; Martens, L.; Sichien, D.; Mossad, O.; Blank, T.; De Prijck, S.; Low, D.; et al. Profiling peripheral nerve macrophages reveals two macrophage subsets with distinct localization, transcriptome and response to injury. Nat. Neurosci. 2020, 23, 676–689. [Google Scholar] [CrossRef]
- Chen, L.; Li, Y.; Zhu, L.; Jin, H.; Kang, X.; Feng, Z. Single-cell RNA sequencing in the context of neuropathic pain: Progress, challenges, and prospects. Transl. Res. 2022, 251, 96–103. [Google Scholar] [CrossRef]
- Renthal, W.; Tochitsky, I.; Yang, L.; Cheng, Y.-C.; Li, E.; Kawaguchi, R.; Geschwind, D.H.; Woolf, C.J. Transcriptional Reprogramming of Distinct Peripheral Sensory Neuron Subtypes after Axonal Injury. Neuron 2020, 108, 128–144.e9. [Google Scholar] [CrossRef] [PubMed]
- Hao, Y.; Hao, S.; Andersen-Nissen, E.; Mauck, W.M., 3rd; Zheng, S.; Butler, A.; Lee, M.J.; Wilk, A.J.; Darby, C.; Zager, M.; et al. Integrated analysis of multimodal single-cell data. Cell 2021, 184, 3573–3587.e29. [Google Scholar] [CrossRef] [PubMed]
- Keren-Shaul, H.; Spinrad, A.; Weiner, A.; Matcovitch-Natan, O.; Dvir-Szternfeld, R.; Ulland, T.K.; David, E.; Baruch, K.; Lara-Astaiso, D.; Toth, B.; et al. A Unique Microglia Type Associated with Restricting Development of Alzheimer’s Disease. Cell 2017, 169, 1276–1290.e17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed]
- Tsujino, H.; Kondo, E.; Fukuoka, T.; Dai, Y.; Tokunaga, A.; Miki, K.; Yonenobu, K.; Ochi, T.; Noguchi, K. Activating Transcription Factor 3 (ATF3) Induction by Axotomy in Sensory and Motoneurons: A Novel Neuronal Marker of Nerve Injury. Mol. Cell. Neurosci. 2000, 15, 170–182. [Google Scholar] [CrossRef]
- Tomlinson, J.E.; Žygelytė, E.; Grenier, J.K.; Edwards, M.G.; Cheetham, J. Temporal changes in macrophage phenotype after peripheral nerve injury. J. Neuroinflamm. 2018, 15, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Tabatabaei, N.; Hou, S.; Kim, K.-W.; Tahmasebi, S. Signaling pathways that control mRNA translation initiation in macrophages. Cell. Signal. 2020, 73, 109700. [Google Scholar] [CrossRef]
- Basu, A.; Poddar, D.; Robinet, P.; Smith, J.D.; Febbraio, M.; BaldwinIII, W.M.; Mazumder, B. Ribosomal Protein L13a Deficiency in Macrophages Promotes Atherosclerosis by Limiting Translation Control-Dependent Retardation of Inflammation. Arter. Thromb. Vasc. Biol. 2014, 34, 533–542. [Google Scholar] [CrossRef] [Green Version]
- Haidar, M.; Asselbergh, B.; Adriaenssens, E.; De Winter, V.; Timmermans, J.P.; Auer-Grumbach, M.; Juneja, M.; Timmerman, V. Neuropathy-Causing Mutations in HSPB1 Impair Autophagy by Disturbing the Formation of SQSTM1/p62 Bodies. Autophagy 2019, 15, 1051–1068. [Google Scholar] [CrossRef]
- Tang-Huau, T.-L.; Gueguen, P.; Goudot, C.; Durand, M.; Bohec, M.; Baulande, S.; Pasquier, B.; Amigorena, S.; Segura, E. Human in vivo-generated monocyte-derived dendritic cells and macrophages cross-present antigens through a vacuolar pathway. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Wu, F.; Xu, D.; Zhi, Z.; Xu, G. Inhibition of TREM1 reduces inflammation and oxidative stress after spinal cord injury (SCI) associated with HO-1 expressions. Biomed. Pharmacother. 2019, 109, 2014–2021. [Google Scholar] [CrossRef] [PubMed]
- Jiao, H.; Rutar, M.; Fernando, N.; Yednock, T.; Sankaranarayanan, S.; Aggio-Bruce, R.; Provis, J.; Natoli, R. Subretinal macrophages produce classical complement activator C1q leading to the progression of focal retinal degeneration. Mol. Neurodegener. 2018, 13, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Niehaus, J.K.; Taylor-Blake, B.; Loo, L.; Simon, J.M.; Zylka, M.J. Spinal macrophages resolve nociceptive hypersensitivity after peripheral injury. Neuron 2021, 109, 1274–1282.e6. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Korvenlaita, N.; Louhivuori, L. Re-Analysis of Single-Nucleus Transcriptomics Reveals Diverse Dorsal Root Ganglia Macrophage Responses Following Peripheral Nerve Injury. Biomedicines 2022, 10, 3295. https://doi.org/10.3390/biomedicines10123295
Korvenlaita N, Louhivuori L. Re-Analysis of Single-Nucleus Transcriptomics Reveals Diverse Dorsal Root Ganglia Macrophage Responses Following Peripheral Nerve Injury. Biomedicines. 2022; 10(12):3295. https://doi.org/10.3390/biomedicines10123295
Chicago/Turabian StyleKorvenlaita, Nea, and Lauri Louhivuori. 2022. "Re-Analysis of Single-Nucleus Transcriptomics Reveals Diverse Dorsal Root Ganglia Macrophage Responses Following Peripheral Nerve Injury" Biomedicines 10, no. 12: 3295. https://doi.org/10.3390/biomedicines10123295