
Local and Systemic Injections of Human Cord Blood Myeloid-Derived Suppressor Cells to Prevent Graft Rejection in Corneal Transplantation

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Murine Model of Corneal Transplantation
2.3. MDSCs Administration
2.4. Immunofluorescent Staining
2.5. Corneal Whole Mount Staining by Immunofluorescence
2.6. Real-Time Polymerase Chain Reaction (PCR)
2.7. Flow Cytometry Assay
2.8. Statistical Analysis
3. Results
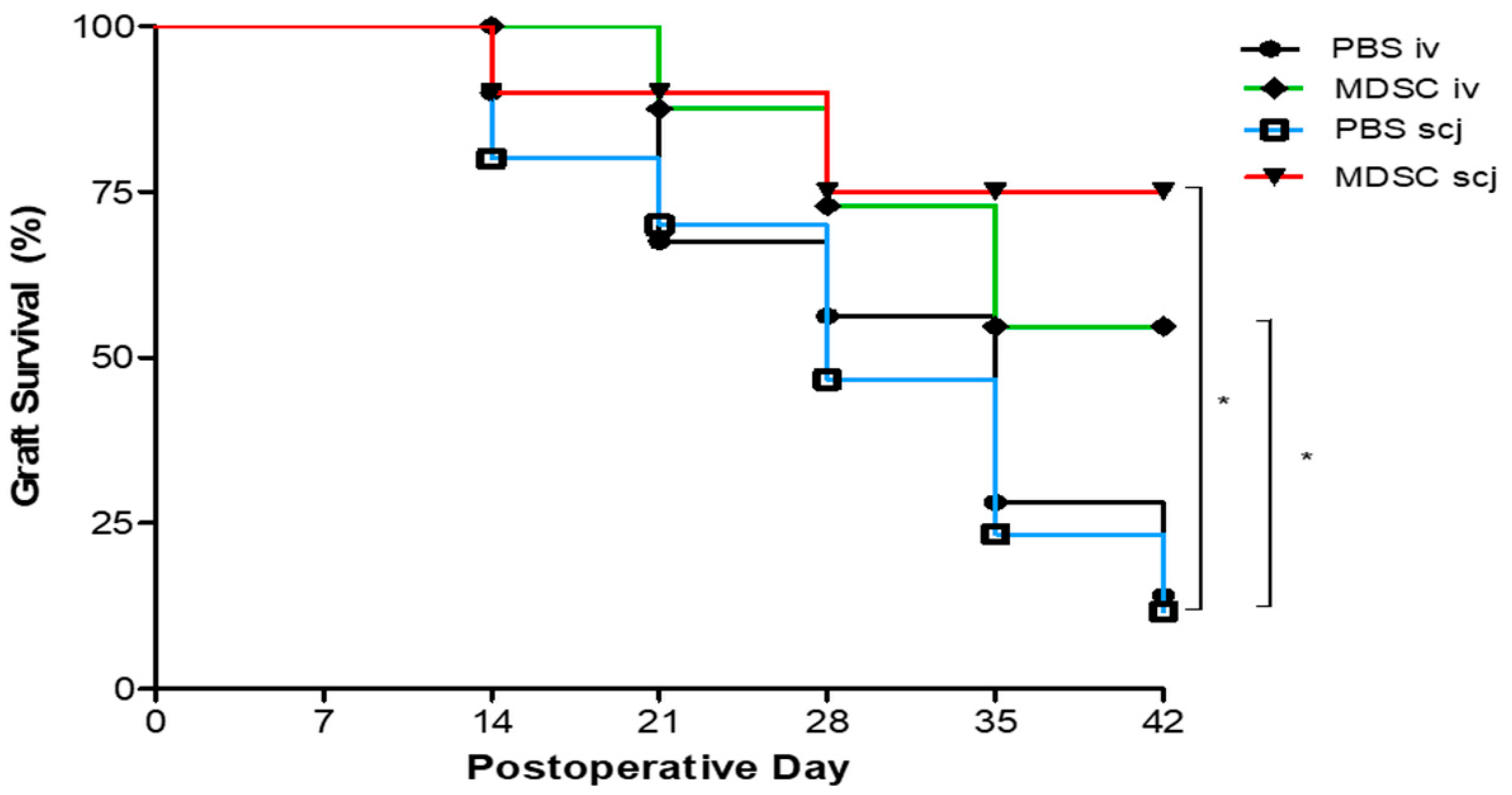
3.1. Local and Systemic MDSCs Administration Enhanced the Corneal Graft Survival after Corneal Transplantation
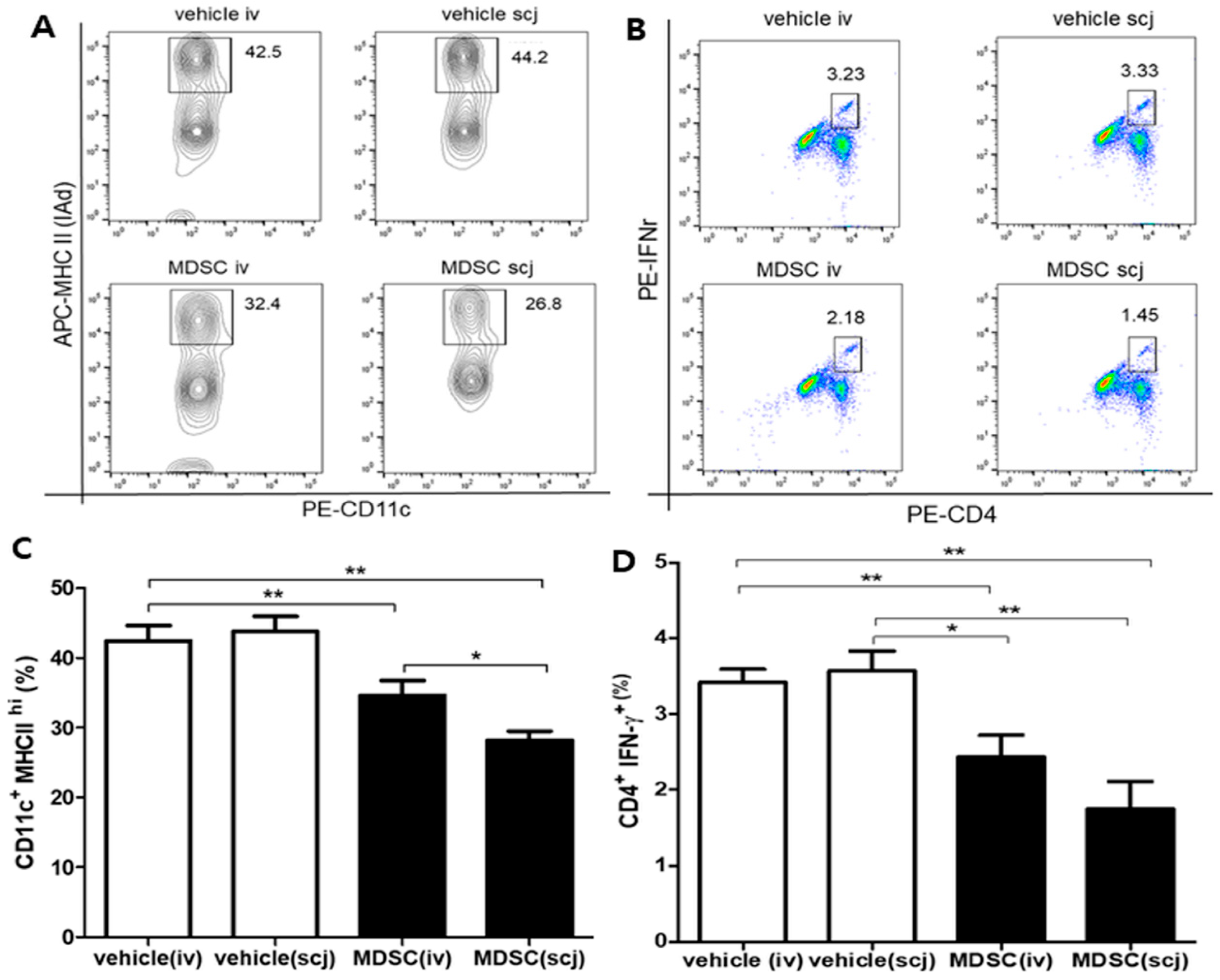
3.2. Local and Systemic MDSCs Suppressed DC Maturation and IFNγ-Expressing Effector CD4+ T Cell Generation in Draining LNs in the Corneal Transplantation Model
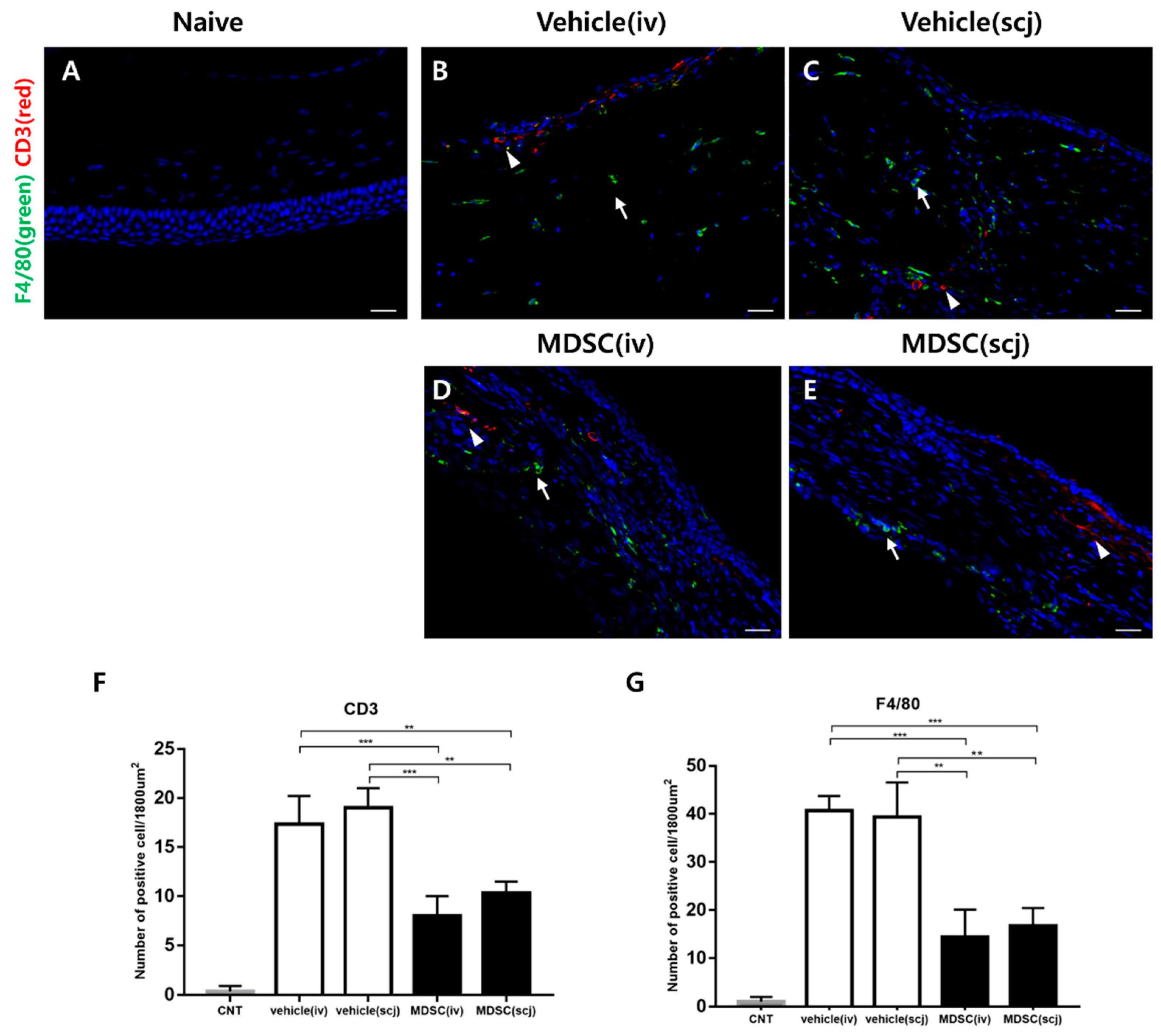
3.3. MDSCs Reduced Recruiting Macrophages and CD3+ T Cells in Grafted Corneas
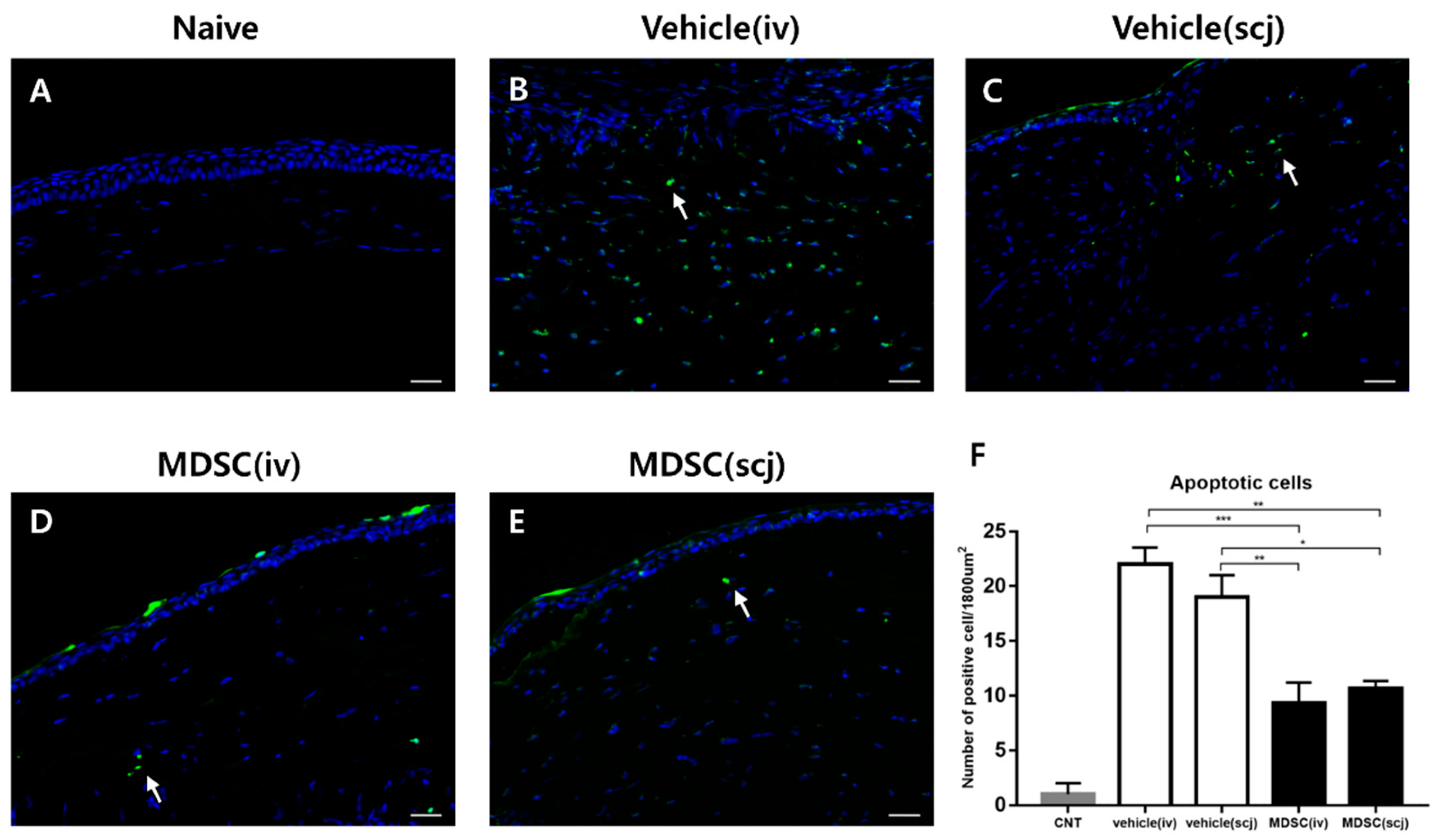
3.4. MDSCs Prevented Cellular Apoptosis in Grafted Corneas
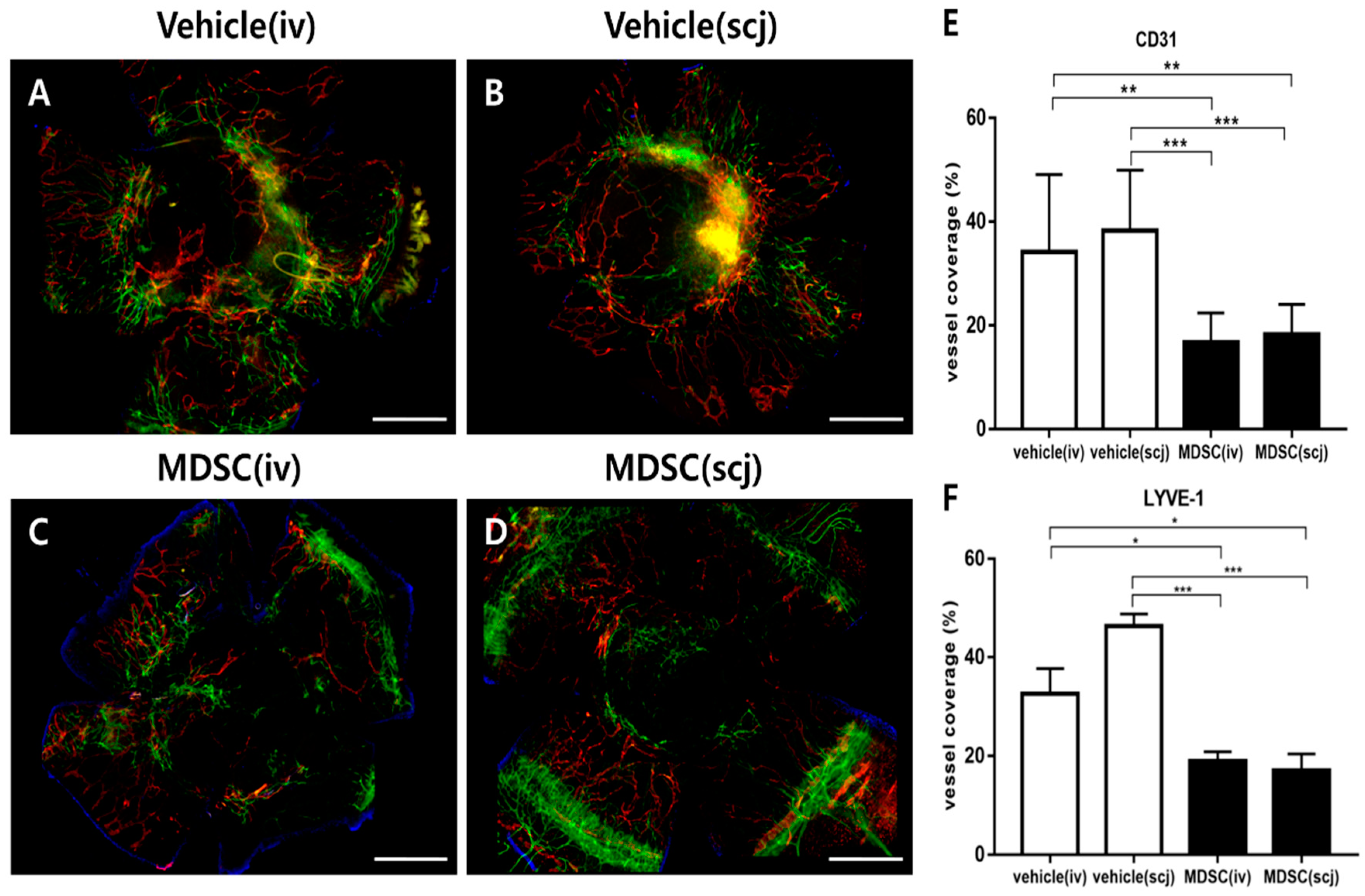
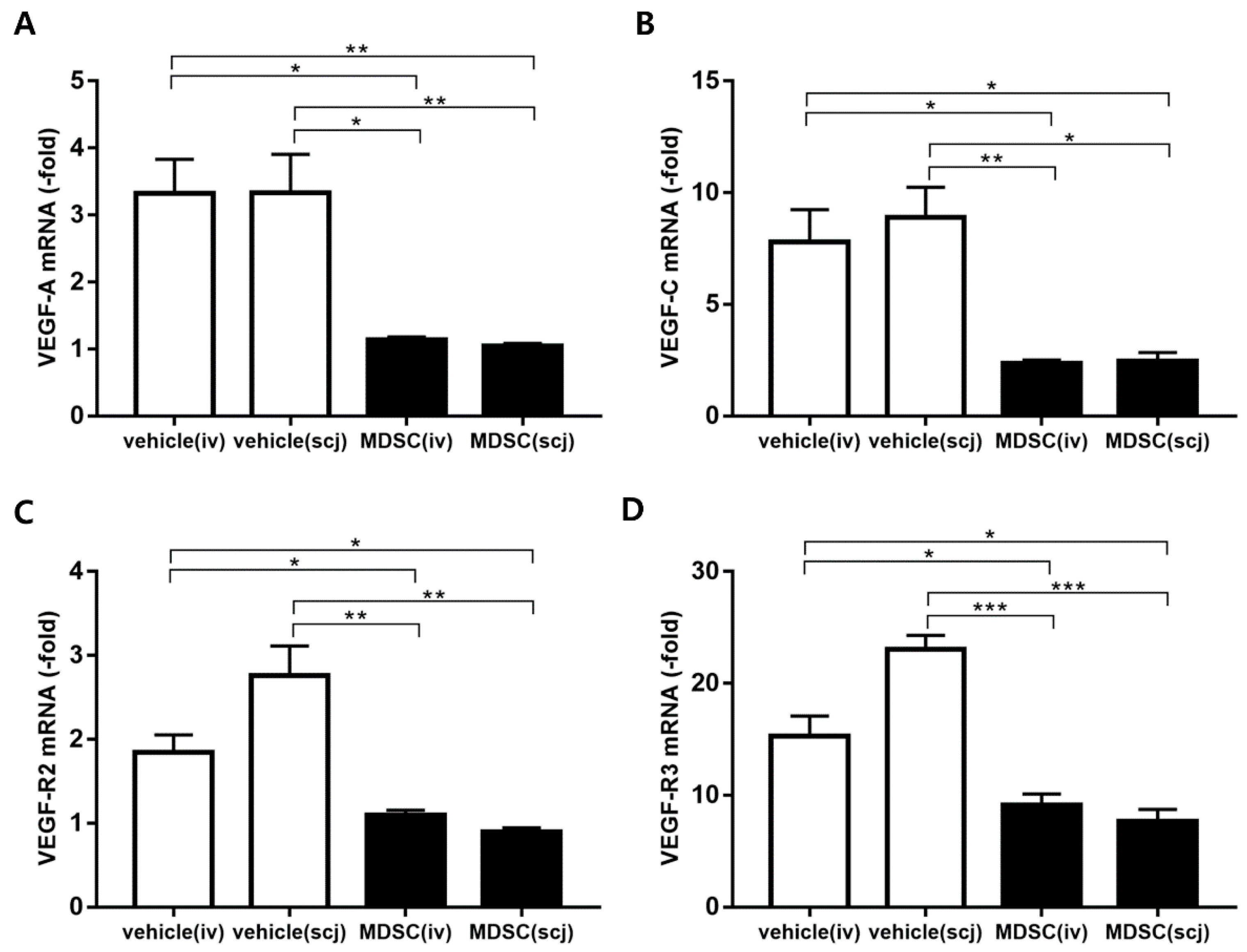
3.5. MDSCs Alleviated Neovascularization and Lymphangiogenesis on Grafted Corneas
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gain, P.; Jullienne, R.; He, Z.; Aldossary, M.; Acquart, S.; Cognasse, F.; Thuret, G. Global Survey of Corneal Transplantation and Eye Banking. JAMA Ophthalmol. 2016, 134, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Sanfilippo, F.; MacQueen, J.M.; Vaughn, W.K.; Foulks, G.N. Reduced graft rejection with good HLA-A and B matching in high-risk corneal transplantation. N. Engl. J. Med. 1986, 315, 29–35. [Google Scholar] [CrossRef]
- Amouzegar, A.; Chauhan, S.K.; Dana, R. Alloimmunity and Tolerance in Corneal Transplantation. J. Immunol. 2016, 196, 3983–3991. [Google Scholar] [CrossRef] [PubMed]
- Coster, D.J.; Williams, K.A. The impact of corneal allograft rejection on the long-term outcome of corneal transplantation. Am. J. Ophthalmol. 2005, 140, 1112–1122. [Google Scholar] [CrossRef] [PubMed]
- Williams, K.A.; Muehlberg, S.M.; Lewis, R.F.; Coster, D.J. How successful is corneal transplantation? A report from the Australian corneal graft register. Eye 1995, 9, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Koay, P.Y.; Lee, W.H.; Figueiredo, F.C. Opinions on risk factors and management of corneal graft rejection in the United kingdom. Cornea 2005, 24, 292–296. [Google Scholar] [CrossRef]
- Panda, A.; Vanathi, M.; Kumar, A.; Dash, Y.; Priya, S. Corneal graft rejection. Surv. Ophthalmol. 2007, 52, 375–396. [Google Scholar] [CrossRef]
- Poon, A.C.; Forbes, J.E.; Dart, J.K.; Subramaniam, S.; Bunce, C.; Madison, P.; Ficker, L.A.; Tuft, S.J.; Gartry, D.S.; Buckley, R.J. Systemic cyclosporin A in high risk penetrating keratoplasties: A case-control study. Br. J. Ophthalmol. 2001, 85, 1464–1469. [Google Scholar] [CrossRef]
- Uchiyama, E.; Papaliodis, G.N.; Lobo, A.M.; Sobrin, L. Side-effects of anti-inflammatory therapy in uveitis. Semin. Ophthalmol. 2014, 29, 456–467. [Google Scholar] [CrossRef]
- Yamada, J.; Hamuro, J.; Sano, Y.; Maruyama, K.; Kinoshita, S. Allogeneic corneal tolerance in rodents with long-term graft survival. Transplantation 2005, 79, 1362–1369. [Google Scholar] [CrossRef]
- Inomata, T.; Fujimoto, K.; Okumura, Y.; Zhu, J.; Fujio, K.; Shokirova, H.; Miura, M.; Okano, M.; Funaki, T.; Sung, J.; et al. Novel immunotherapeutic effects of topically administered ripasudil (K-115) on corneal allograft survival. Sci. Rep. 2020, 10, 19817. [Google Scholar] [CrossRef]
- Azimzade, Y.; Hong, J.; Mashaghi, A. Immunophysical analysis of corneal neovascularization: Mechanistic insights and implications for pharmacotherapy. Sci. Rep. 2017, 22, 12220. [Google Scholar] [CrossRef] [PubMed]
- Slegers, T.P.; Broersma, L.; van Rooijen, N.; Hooymans, J.M.; van Rij, G.; van der Gaag, R. Macrophages play a role in the early phase of corneal allograft rejection in rats. Transplantation 2004, 77, 1641–1646. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, S.K.; Jurkunas, U.; Funaki, T.; Dastjerdi, M.; Dana, R. Quantification of allospecific and nonspecific corneal endothelial cell damage after corneal transplantation. Eye 2015, 29, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Hargrave, S.L.; Hay, C.; Mellon, J.; Mayhew, E.; Niederkorn, J.Y. Fate of MHC-matched corneal allografts in Th1-deficient hosts. Investig. Ophthalmol. Vis. Sci. 2004, 45, 1188–1193. [Google Scholar] [CrossRef]
- Hamrah, P.; Liu, Y.; Zhang, Q.; Dana, M.R. Alterations in corneal stromal dendritic cell phenotype and distribution in inflammation. Arch. Ophthalmol. 2003, 121, 1132–1140. [Google Scholar] [CrossRef]
- Gabrilovich, D.I.; Nagaraj, S. Myeloid-derived suppressor cells as regulators of the immune system. Nat. Rev. Immunol. 2009, 9, 162–174. [Google Scholar] [CrossRef]
- Sinha, P.; Okoro, C.; Foell, D.; Freeze, H.H.; Ostrand-Rosenberg, S.; Srikrishna, G. Proinflammatory S100 proteins regulate the accumulation of myeloid-derived suppressor cells. J. Immunol. 2008, 181, 4666–4675. [Google Scholar] [CrossRef]
- Nicholson, L.B.; Raveney, B.J.; Munder, M. Monocyte dependent regulation of autoimmune inflammation. Curr. Mol. Med. 2009, 9, 23–29. [Google Scholar] [CrossRef]
- Suh, Y.G.; Kim, J.K.; Byun, J.S.; Yi, H.S.; Lee, Y.S.; Eun, H.S.; Kim, S.Y.; Han, K.H.; Lee, K.S.; Duester, G.; et al. CD11b(+) Gr1(+) bone marrow cells ameliorate liver fibrosis by producing interleukin-10 in mice. Hepatology 2012, 56, 1902–1912. [Google Scholar] [CrossRef]
- Fujimura, T.; Mahnke, K.; Enk, A.H. Myeloid derived suppressor cells and their role in tolerance induction in cancer. J. Dermatol. Sci. 2010, 59, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Park, M.Y.; Lim, B.G.; Kim, S.Y.; Sohn, H.J.; Kim, S.; Kim, T.G. GM-CSF Promotes the Expansion and Differentiation of Cord Blood Myeloid-Derived Suppressor Cells, Which Attenuate Xenogeneic Graft-vs.-Host Disease. Front. Immunol. 2019, 10, 183. [Google Scholar] [CrossRef] [PubMed]
- Ostanin, D.V.; Bhattacharya, D. Myeloid-derived suppressor cells in the inflammatory bowel diseases. Inflamm. Bowel Dis. 2013, 19, 2468–2477. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Wang, S.; Li, J.; Zhang, W.; Zheng, L.; Yang, C.; Zhu, T.; Rong, R. The mTOR signal regulates myeloid-derived suppressor cells differentiation and immunosuppressive function in acute kidney injury. Cell Death Dis. 2017, 8, e2695. [Google Scholar] [CrossRef]
- Jeong, H.J.; Lee, H.J.; Ko, J.H.; Cho, B.J.; Park, S.Y.; Park, J.W.; Choi, S.R.; Heo, J.W.; Yoon, S.O.; Oh, J.Y. Myeloid-Derived Suppressor Cells Mediate Inflammation Resolution in Humans and Mice with Autoimmune Uveoretinitis. J. Immunol. 2018, 200, 1306–1315. [Google Scholar] [CrossRef]
- Lee, H.S.; Kwon, J.Y.; Joo, C.K. Comparative immunological study of penetrating and anterior lamellar keratoplasty in a murine model. Exp. Eye Res. 2018, 169, 141–148. [Google Scholar] [CrossRef]
- Lim, J.Y.; Ryu, D.B.; Park, M.Y.; Lee, S.E.; Park, G.; Kim, T.G.; Min, C.K. Ex Vivo Generated Human Cord Blood Myeloid-Derived Suppressor Cells Attenuate Murine Chronic Graft-versus-Host Diseases. Biol. Blood Marrow Transplant. 2018, 24, 2381–2396. [Google Scholar] [CrossRef]
- Lee, P.; Wang, C.C.; Adamis, A.P. Ocular neovascularization: An epidemiologic review. Surv. Ophthalmol. 1998, 43, 245–269. [Google Scholar] [CrossRef]
- Beebe, D.C. Maintaining transparency: A review of the developmental physiology and pathophysiology of two avascular tissues. Semin. Cell Dev. Biol. 2008, 19, 125–133. [Google Scholar] [CrossRef]
- Tshionyi, M.; Shay, E.; Lunde, E.; Lin, A.; Han, K.Y.; Jain, S.; Chang, J.H.; Azar, D.T. Hemangiogenesis and lymphangiogenesis in corneal pathology. Cornea 2012, 31, 74–80. [Google Scholar] [CrossRef]
- Inomata, T.; Mashaghi, A.; Di Zazzo, A.; Lee, S.M.; Chiang, H.; Dana, R. Kinetics of Angiogenic Responses in Corneal Transplantation. Cornea 2017, 36, 491–496. [Google Scholar] [CrossRef] [PubMed]
- Dohlman, T.H.; Omoto, M.; Hua, J.; Stevenson, W.; Lee, S.M.; Chauhan, S.K.; Dana, R. VEGF-trap aflibercept significantly improves long-term graft survival in high-risk corneal transplantation. Transplantation 2015, 99, 678–686. [Google Scholar] [CrossRef] [PubMed]
- Bizymi, N.; Georgopoulou, A.; Mastrogamvraki, N.; Matheakakis, A.; Gontika, I.; Fragiadaki, I.; Mavroudi, I.; Papadaki, H.A. Myeloid-Derived Suppressor Cells (MDSC) in the Umbilical Cord Blood: Biological Significance and Possible Therapeutic Applications. J. Clin. Med. 2022, 11, 727. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.N.; Sun, E.G.; Cho, S.H. IL-12 Enhances Immune Response by Modulation of Myeloid Derived Suppressor Cells in Tumor Microenvironment. Chonnam Med. J. 2019, 55, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Bunt, S.K.; Clements, V.K.; Hanson, E.M.; Sinha, P.; Ostrand-Rosenberg, S. Inflammation enhances myeloid-derived suppressor cell cross-talk by signaling through Toll-like receptor 4. J. Leukoc. Biol. 2009, 85, 996–1004. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.E.; Gan, J.; Zhang, R.D.; Cheng, Y.R.; Huang, G.J. Up-regulated myeloid-derived suppressor cell contributes to hepatocellular carcinoma development by impairing dendritic cell function. Scand. J. Gastroenterol. 2011, 46, 156–164. [Google Scholar] [CrossRef]
- Dietz, S.; Schwarz, J.; Vogelmann, M.; Spring, B.; Molnár, K.; Orlikowsky, T.W.; Wiese, F.; Holzer, U.; Poets, C.F.; Gille, C.; et al. Cord blood granulocytic myeloid-derived suppressor cells impair monocyte T cell stimulatory capacity and response to bacterial stimulation. Pediatr. Res. 2019, 86, 608–615. [Google Scholar] [CrossRef]
- Stein-Streilein, J.; Streilein, J.W. Anterior chamber associated immune deviation (ACAID): Regulation, biological relevance, and implications for therapy. Int. Rev. Immunol. 2002, 21, 123–152. [Google Scholar] [CrossRef]
- Cabeza-Cabrerizo, M.; Cardoso, A.; Minutti, C.M.; Pereira da Costa, M.; Reis e Sousa, C. Dendritic Cells Revisited. Annu. Rev. Immunol. 2021, 39, 131–166. [Google Scholar] [CrossRef]
- Oh, J.Y.; Lee, H.J.; Ko, A.Y.; Ko, J.H.; Kim, M.K.; Wee, W.R. Analysis of macrophage phenotype in rejected corneal allografts. Investig. Ophthalmol. Vis. Sci. 2013, 54, 7779–7784. [Google Scholar] [CrossRef]
- Jensen, K.P.; Hongo, D.A.; Ji, X.; Zheng, P.; Pawar, R.; Wu, T.H.; Busque, S.; Scandling, J.D.; Shizuru, J.A.; Lowsky, R.; et al. Development of immunosuppressive myeloid cells to induce tolerance in solid organ and hematopoietic cell transplant recipients. Blood Adv. 2021, 5, 3290–3302. [Google Scholar] [CrossRef] [PubMed]
- Kammrath Betancor, P.; Hildebrand, A.; Böhringer, D.; Emmerich, F.; Schlunck, G.; Reinhard, T.; Lapp, T. Activation of human macrophages by human corneal allogen in vitro. PLoS ONE 2018, 13, e0194855. [Google Scholar] [CrossRef]
- Ren, Y.; Dong, X.; Zhao, H.; Feng, J.; Chen, B.; Zhou, Y.; Peng, Y.; Zhang, L.; Zhou, Q.; Li, Y.; et al. Myeloid-derived suppressor cells improve corneal graft survival through suppressing angiogenesis and lymphangiogenesis. Am. J. Transplant. 2021, 21, 552–566. [Google Scholar] [CrossRef] [PubMed]
- Cursiefen, C.; Chen, L.; Borges, L.P.; Jackson, D.; Cao, J.; Radziejewski, C.; D’Amore, P.A.; Dana, M.R.; Wiegand, S.J.; Streilein, J.W. VEGF-A stimulates lymphangiogenesis and hemangiogenesis in inflammatory neovascularization via macrophage recruitment. J. Clin. Investig. 2004, 113, 1040–1050. [Google Scholar] [CrossRef] [PubMed]
- Maruyama, K.; Ii, M.; Cursiefen, C.; Jackson, D.G.; Keino, H.; Tomita, M.; Van Rooijen, N.; Takenaka, H.; D’Amore, P.A.; Stein-Streilein, J.; et al. Inflammation-induced lymphangiogenesis in the cornea arises from CD11b-positive macrophages. J. Clin. Investig. 2005, 115, 2363–2372. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Wang, B.; Jia, B.; Guan, J.; Zeng, H.; Pan, Z. Effects of Adoptive Transferring Different Sources of Myeloid-Derived Suppressor Cells in Mice Corneal Transplant Survival. Transplantation 2015, 99, 2102–2108. [Google Scholar] [CrossRef]
- Fujimoto, K.; Uchida, K.; Yin, E.; Zhu, J.; Kojima, Y.; Uchiyama, M.; Yamamoto, Y.; Bashuda, H.; Matsumoto, R.; Tokushige, K.; et al. Analysis of therapeutic potential of monocytic myeloid-derived suppressor cells in cardiac allotransplantation. Transpl. Immunol. 2021, 67, 101405. [Google Scholar] [CrossRef]
- Cao, P.; Sun, Z.; Zhang, F.; Zhang, J.; Zheng, X.; Yu, B.; Zhao, Y.; Wang, W.; Wang, W. TGF-β Enhances Immunosuppression of Myeloid-Derived Suppressor Cells to Induce Transplant Immune Tolerance Through Affecting Arg-1 Expression. Front. Immunol. 2022, 13, 919674. [Google Scholar] [CrossRef]
- Gao, J.; Dennis, J.E.; Muzic, R.F.; Lundberg, M.; Caplan, A.I. The dynamic in vivo distribution of bone marrow-derived mesenchymal stem cells after infusion. Cells Tissues Organs. 2001, 169, 12–20. [Google Scholar] [CrossRef]
- Zhang, L.; Li, K.; Liu, X.; Li, D.; Luo, C.; Fu, B.; Cui, S.; Zhu, F.; Zhao, R.C.; Chen, X. Repeated systemic administration of human adipose-derived stem cells attenuates overt diabetic nephropathy in rats. Stem Cells Dev. 2013, 22, 3074–3086. [Google Scholar] [CrossRef]
- Zheng, B.; von See, M.P.; Yu, E.; Gunel, B.; Lu, K.; Vazin, T.; Schaffer, D.V.; Goodwill, P.W.; Conolly, S.M. Quantitative Magnetic Particle Imaging Monitors the Transplantation, Biodistribution, and Clearance of Stem Cells In Vivo. Theranostics 2016, 6, 291–301. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Hodges, A.; Chen, S.H.; Pan, P.Y. Myeloid-derived suppressor cells as cellular immunotherapy in transplantation and autoimmune diseases. Cell Immunol. 2021, 362, 104300. [Google Scholar] [CrossRef]
- Zhu, J.; Inomata, T.; Fujimoto, K.; Uchida, K.; Fujio, K.; Nagino, K.; Miura, M.; Negishi, N.; Okumura, Y.; Akasaki, Y.; et al. Ex Vivo-Induced Bone Marrow-Derived Myeloid Suppressor Cells Prevent Corneal Allograft Rejection in Mice. Investig. Ophthalmol. Vis. Sci. 2021, 62, 3. [Google Scholar] [CrossRef]
- Galindo, S.; de la Mata, A.; López-Paniagua, M.; Herreras, J.M.; Pérez, I.; Calonge, M.; Nieto-Miguel, T. Subconjunctival injection of mesenchymal stem cells for corneal failure due to limbal stem cell deficiency: State of the art. Stem Cell Res. Ther. 2021, 12, 60. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Liang, C.; Hu, H.; Zhou, L.; Xu, B.; Wang, X.; Han, Y.; Nie, Y.; Jia, S.; Liang, J.; et al. Intraperitoneal injection (IP), Intravenous injection (IV) or anal injection (AI)? Best way for mesenchymal stem cells transplantation for colitis. Sci. Rep. 2016, 6, 30696. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Xu, L.; Zhang, Y.; Sun, Y.; Li, G. Systemic and Local Administration of Allogeneic Bone Marrow-Derived Mesenchymal Stem Cells Promotes Fracture Healing in Rats. Cell Transplant. 2015, 24, 2643–2655. [Google Scholar] [CrossRef]
- Ostrand-Rosenberg, S. Myeloid-derived suppressor cells: Multi-talented immune suppressive cells that can be either helpful or harmful. Cell. Immunol. 2021, 365, 104374. [Google Scholar] [CrossRef]
- van Geffen, C.; Heiss, C.; Deißler, A.; Kolahian, S. Pharmacological modulation of myeloid-derived suppressor cells to dampen inflammation. Front. Immunol. 2022, 30, 933847. [Google Scholar] [CrossRef]
- Vanhaver, C.; van der Bruggen, P.; Bruger, A.M. MDSC in Mice and Men: Mechanisms of Immunosuppression in Cancer. J. Clin. Med. 2021, 10, 2872. [Google Scholar] [CrossRef]
- Bizymi, N.; Matthaiou, A.M.; Matheakakis, A.; Voulgari, I.; Aresti, N.; Zavitsanou, K.; Karasachinidis, A.; Mavroudi, I.; Pontikoglou, C.; Papadaki, H.A. New Perspectives on Myeloid-Derived Suppressor Cells and Their Emerging Role in Haematology. J. Clin. Med. 2022, 11, 5326. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.-y.; Sohn, H.-J.; Kim, C.-H.; Kim, T.-G.; Lee, H.S. Local and Systemic Injections of Human Cord Blood Myeloid-Derived Suppressor Cells to Prevent Graft Rejection in Corneal Transplantation. Biomedicines 2022, 10, 3223. https://doi.org/10.3390/biomedicines10123223
Lee J-y, Sohn H-J, Kim C-H, Kim T-G, Lee HS. Local and Systemic Injections of Human Cord Blood Myeloid-Derived Suppressor Cells to Prevent Graft Rejection in Corneal Transplantation. Biomedicines. 2022; 10(12):3223. https://doi.org/10.3390/biomedicines10123223
Chicago/Turabian StyleLee, Jae-young, Hyun-Jung Sohn, Chang-Hyun Kim, Tai-Gyu Kim, and Hyun Soo Lee. 2022. "Local and Systemic Injections of Human Cord Blood Myeloid-Derived Suppressor Cells to Prevent Graft Rejection in Corneal Transplantation" Biomedicines 10, no. 12: 3223. https://doi.org/10.3390/biomedicines10123223