
Periostin as a Biomarker in the Setting of Glomerular Diseases—A Review of the Current Literature
{kind=link}
{kind=link}
Abstract
:1. Introduction
- -
- A C-terminal domain which is a site of proteolytic cleavage that renders the different isoforms of periostin;
- -
- An N-terminal EMI domain which renders the capacity to interact with collagen I and fibronectin;
- -
- A middle region consisting of four fasciclin-I domains that contain binding sites for bone morphogenetic protein 1 (BMP-1) and αVβ3 and αVβ5 integrins, through which it manifests its aforementioned capacity to mediate cell adhesion and migration.
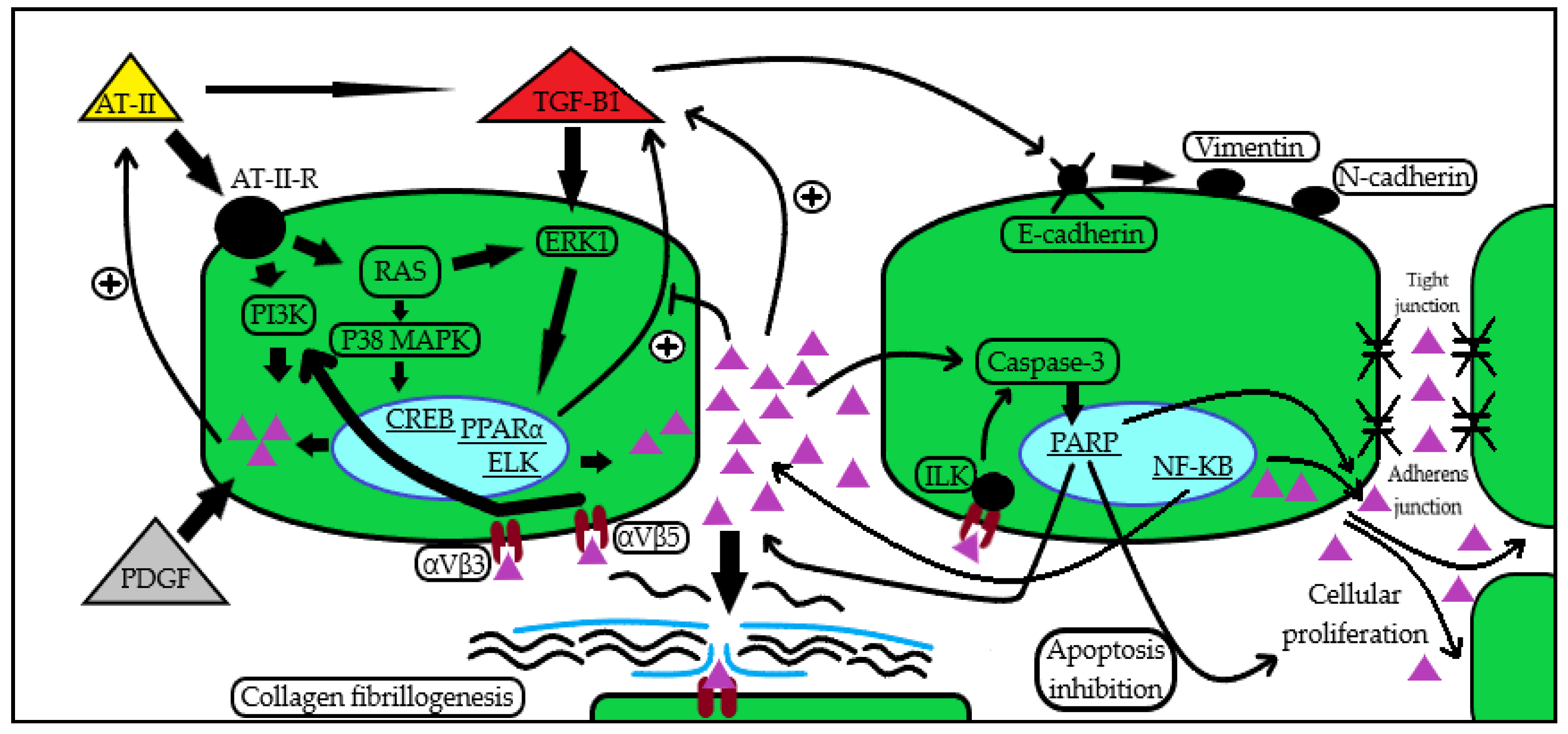
2. Periostin Functions and Mechanisms
3. Periostin in CKD and in the Kidney
4. Periostin as a Biomarker in Glomerular Disease
5. Clinical Implications
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, A.Y.; Zheng, H. Periostin, a multifunctional matricellular protein in inflammatory and tumor microenvironments. Matrix Biol. 2014, 37, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Takeshita, S.; Kikuno, R. Osteoblast—Specific factor 2: Cloning of a putative bone adhesion protein with homology with the insect protein fasciclin I. Biochem. J. 1993, 294, 27127–27128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bornstein, P.; Sage, E.H. Matricellular proteins: Extracellular modulators of cell function. Curr. Opin. Cell Biol. 2002, 14, 608–616. [Google Scholar] [CrossRef] [PubMed]
- Prakoura, N.; Chatziantoniou, C. Periostin and Discoidin Domain Receptor 1: New Biomarkers or Targets for Therapy of Renal Disease. Front. Med. 2017, 4, 52. [Google Scholar] [CrossRef] [Green Version]
- Hoersch, S.; Andrade-Navarro, M.A. Periostin shows increased evolutionary plasticity in its alternatively spliced region. BMC Evol. Biol. 2010, 10, 30. [Google Scholar] [CrossRef] [Green Version]
- Gillan, L.; Matei, D. Periostin secreted by epithelial ovarian carcinoma is a ligand for alphaVbeta3 and alphaVbeta5 integrins and promotes cell motility. Cancer Res. 2002, 62, 5358–5364. [Google Scholar]
- Conway, S.J.; Izuhara, K. The role of periostin in tissue remodeling across health and disease. Cell. Mol. Life Sci. 2014, 71, 1279–1288. [Google Scholar] [CrossRef] [Green Version]
- Shimazaki, M.; Nakamura, K. Periostin is essential for cardiac healing after acute myocardial infarction. J. Exp. Med. 2008, 205, 295–303. [Google Scholar] [CrossRef]
- Oka, T.; Xu, J. Genetic manipulation of periostin expression reveals a role in cardiac hypertrophy and ventricular remodeling. Circ. Res. 2007, 101, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Naik, P.K.; Bozyk, P.D. Periostin promotes fibrosis and predicts progression in patients with idiopathic pulmonary fibrosis. Am. J. Physiol. Lung Cell. Mol. Physiol. 2012, 303, L1046–L1056. [Google Scholar] [CrossRef] [Green Version]
- Sidhu, S.S.; Yuan, S. Roles of epithelial cell-derived periostin in TGF-beta activation, collagen production, and collagen gel elasticity in asthma. Proc. Natl. Acad. Sci. USA 2010, 107, 14170–14175. [Google Scholar] [CrossRef] [PubMed]
- Corren, J.; Lemanske, R.F. Lebrikizumab treatment in adults with asthma. N. Engl. J. Med. 2011, 365, 1088–1098. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izuhara, K.; Matsumoto, H. Recent developments regarding periostin in bronchial asthma. Allergol. Int. 2015, 64 (Suppl. S1), S3–S10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takayama, G.; Arima, K. Periostin: A novel component of subepithelial fibrosis of bronchial asthma downstream of IL-4 and IL-13 signals. J. Allergy Clin. Immunol. 2006, 118, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Hanania, N.A.; Korenblat, P. Efficacy and safety of lebrikizumab in patients with uncontrolled asthma (LAVOLTA I and LAVOLTA II): Replicate, phase 3, randomised, double-blind, placebo-controlled trials. Lancet Respir. Med. 2016, 4, 781–796. [Google Scholar] [CrossRef] [PubMed]
- Ruan, K.; Bao, S. The multifaceted role of periostin in tumorigenesis. Cell. Mol. Life Sci. 2009, 66, 2219–2230. [Google Scholar] [CrossRef] [Green Version]
- Malanchi, I.; Santamaria-Martínez, A. Interactions between cancer stem cells and their niche govern metastatic colonization. Nature 2011, 481, 85–89. [Google Scholar] [CrossRef] [Green Version]
- Morra, L.; Rechsteiner, M. Relevance of periostin splice variants in renal cell carcinoma. Am. J. Pathol. 2011, 179, 1513–1521. [Google Scholar] [CrossRef]
- Li, G.; Oparil, S.; Sanders, J.M.; Zhang, L.; Dai, M.; Chen, L.B.; Conway, S.J.; McNamara, C.A.; Sarembock, I.J. Phosphatidylinositol- 3-kinase signaling mediates vascular smooth muscle cell expression of peri-ostin in vivo and in vitro. Atherosclerosis 2006, 188, 292–300. [Google Scholar] [CrossRef] [Green Version]
- Li, L.; Fan, D. Angiotensin II increases periostin expression via Ras/p38 MAPK/CREB and ERK1/2/TGF-β1 path-ways in cardiac fibroblasts. Cardiovasc. Res. 2011, 91, 80–89. [Google Scholar] [CrossRef]
- Bian, X.; Bai, Y.; Su, X. Knockdown of periostin attenuates 5/6 nephrectomy-induced intrarenal renin-angiotensin system activation, fibrosis, and inflammation in rats. J. Cell. Physiol. 2019, 234, 22857–22873. [Google Scholar] [CrossRef] [PubMed]
- Guerrot, D.; Dussaule, J.C. Identification of periostin as a critical marker of progression/reversal of hypertensive nephropathy. PLoS ONE 2012, 7, e31974. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mael-Ainin, M.; Abed, A. Inhibition of periostin expression protects against the development of renal inflammation and fibrosis. J. Am. Soc. Nephrol. 2014, 8, 1724–1736. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satirapoj, B.; Wang, Y. Periostin: Novel tissue and urinary biomarker of progressive renal injury induces a coordinated mesenchymal phenotype in tubular cells. Nephrol. Dial. Transplant. 2012, 7, 2702–2711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izuhara, K.; Nunomura, S. Periostin in inflammation and allergy. Cell. Mol. Life Sci. 2017, 74, 4293–4303. [Google Scholar] [CrossRef]
- Sen, K.; Lindenmeyer, M.T. Periostin is induced in glomerular injury and expressed de novo in interstitial renal fibrosis. Am. J. Pathol. 2011, 179, 1756–1767. [Google Scholar] [CrossRef]
- Prakoura, N.; Kavvadas, P. NFκB-Induced Periostin Activates Integrin-β3 Signaling to Promote Renal Injury in GN. J. Am. Soc. Nephrol. 2017, 28, 1475–1490. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Hao, J. Phosphoinositide 3-kinase/protein kinase B/periostin mediated platelet-derived growth factor-induced cell proliferation and extracellular matrix production in lupus nephritis. Exp. Biol. Med. 2017, 242, 160–168. [Google Scholar] [CrossRef] [Green Version]
- Turczyn, A.; Pańczyk-Tomaszewska, M. The role of periostin in kidney diseases. Cent. Eur. J. Immunol. 2021, 46, 494–501. [Google Scholar] [CrossRef]
- Norris, R.A.; Damon, B. Periostin regulates collagen fibrillogenesis and the biome-chanical properties of connective tissues. J. Cell. Biochem. 2007, 101, 695–711. [Google Scholar] [CrossRef] [Green Version]
- Kang, Y.S.; Li, Y. Inhibition of integrin-linked kinase blocks podocyte epithelial-mesenchymal transition and ameliorates proteinuria. Kidney Int. 2010, 78, 363–373. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.H.; Yang, S.H. Experimental inhibition of periostin attenuates kidney fibrosis. Am. J. Nephrol. 2017, 46, 501–517. [Google Scholar] [CrossRef] [PubMed]
- Bian, X.; Su, X.; Wang, Y.; Zhao, G.; Zhang, B.; Li, D. Periostin contributes to renal and cardiac dysfunction in rats with chronic kidney disease: Reduction of PPARα. Biochimie 2019, 160, 172–182. [Google Scholar] [CrossRef] [PubMed]
- Lyu, H.; Li, X. Overexpression of microRNA-21 mediates Ang II-induced renal fibrosis by activating the TGF-β1/Smad3 pathway via suppressing PPARα. J. Pharmacol. Sci. 2019, 141, 70–78. [Google Scholar] [CrossRef]
- Levey, A.S.; Atkins, R. Chronic kidney disease as a global public health problem: Approaches and initiatives. A position statement from Kidney Disease Improving Global Outcomes. Kidney Int. 2007, 72, 247–259. [Google Scholar] [CrossRef] [Green Version]
- Schnaper, H.W. Remnant nephron physiology and the progression of chronic kidney disease. Pediatr. Nephrol. 2014, 29, 193–202. [Google Scholar] [CrossRef] [Green Version]
- Panizo, S.; Martínez-Arias, L. Fibrosis in Chronic Kidney Disease: Pathogenesis and Consequences. Int. J. Mol. Sci. 2021, 22, 408. [Google Scholar] [CrossRef]
- Hsu, C.Y.; Xie, D. Urine biomarkers of tubular injury do not improve on the clinical model predicting chronic kidney disease progression. Kidney Int. 2017, 91, 196–203. [Google Scholar] [CrossRef] [Green Version]
- Satirapoj, B.; Witoon, R. Urine periostin as a biomarker of renal injury in chronic allograft nephropathy. Transplant. Proc. 2014, 46, 135–140. [Google Scholar] [CrossRef]
- Satirapoj, B.; Tassanasorn, S. Periostin as a tissue and urinary biomarker of renal injury in type 2 diabetes mellitus. PLoS ONE 2015, 10, e0124055. [Google Scholar] [CrossRef]
- Wantanasiri, P.; Satirapoj, B. Periostin: A novel tissue biomarker correlates with chronicity index and renal function in lupus nephritis patients. Lupus 2015, 24, 835–845. [Google Scholar] [CrossRef] [PubMed]
- Wallace, D.P.; Quante, M.T. Periostin induces proliferation of human autosomal dominant polycystic kidney cells through alphaV-integrin receptor. Am. J. Physiol. Renal. Physiol. 2008, 295, F1463–F1471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chimenz, R.; Chirico, V. HMGB-1 and TGFβ-1 highlight immuno-inflammatory and fibrotic processes before proteinuria onset in pediatric patients with Alport syndrome. J. Nephrol. 2021, 34, 1915–1924. [Google Scholar] [CrossRef] [PubMed]
- Bennett, M.R.; Devarajan, P. Chapter 1—Characteristics of an Ideal Biomarker of Kidney Diseases. In Biomarkers of Kidney Disease; Academic Press: Cambridge, MA, USA, 2011; pp. 1–24. ISBN 9780123756725. [Google Scholar]
- Cui, D.; Huang, Z. The multifaceted role of periostin in priming the tumor microenvironments for tumor progression. Cell. Mol. Life Sci. 2017, 74, 4287–4291. [Google Scholar] [CrossRef] [PubMed]
- Izuhara, K.; Nunomura, S. Periostin: An emerging biomarker for allergic diseases. Allergy 2019, 74, 2116–2128. [Google Scholar] [CrossRef] [Green Version]
- Fujitani, H.; Kasuga, S. Age-related changes in serum periostin level in allergic and non-allergic children. Allergol. Int. 2019, 68, 285–286. [Google Scholar] [CrossRef]
- Turczyn, A.; Krzemień, G. Serum and urine periostin and cytokeratin-18 in children with congenital obstructive nephropathy. Cent. Eur. J. Immunol. 2022, 47, 63–72. [Google Scholar] [CrossRef]
- Rodrigues, J.C.; Haas, M. IgA Nephropathy. Clin. J. Am. Soc. Nephrol. 2017, 12, 677–686. [Google Scholar] [CrossRef] [Green Version]
- Trimarchi, H.; Barratt, J. Oxford Classification of IgA nephropathy 2016, An update from the IgA Nephropathy Classification Working Group. Kidney Int. 2017, 91, 1014–1021. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.H.; Lee, J.P. Urinary Periostin Excretion Predicts Renal Outcome in IgA Nephropathy. Am. J. Nephrol. 2016, 44, 481–492. [Google Scholar] [CrossRef]
- Wantanasiri, P.; Satirapoj, B. Potential of Periostin as a Urinary Biomarker Correlated with Renal Function in Lupus Nephritis and IgA Nephropathy Patients. J. Med. Assoc. Thail. 2018, 101, 1479–1485. [Google Scholar]
- Wantanasiri, P.; Satirapoj, B. Periostin staining within renal tissues from IgA nephropathy patients. Songklanakarin J. Sci. Technol. 2018, 40, 155–161. [Google Scholar]
- Mizerska-Wasiak, M.; Płatos, E. The Usefulness of Vanin-1 and Periostin as Markers of an Active Autoimmune Process or Renal Fibrosis in Children with IgA Nephropathy and IgA Vasculitis with Nephritis-A Pilot Study. J. Clin. Med. 2022, 11, 1265. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Lin, Q. Periostin Contributes to Immunoglobulin a Nephropathy by Promoting the Proliferation of Mesangial Cells: A Weighted Gene Correlation Network Analysis. Front. Genet. 2021, 11, 595757. [Google Scholar] [CrossRef]
- Swaminathan, S.; Leung, N.; Lager, D.J.; Melton, L.J.; Bergstralh, E.J.; Rohlinger, A.; Fervenza, F.C. Changing Incidence of Glomerular Disease in Olmsted County, Minnesota: A 30-Year Renal Biopsy Study. Clin. J. Am. Soc. Nephrol. 2006, 1, 483–487. [Google Scholar]
- Zhou, X.; Zhang, Y. Systematic identification of key extracellular proteins as the potential biomarkers in lupus nephritis. Front. Immunol. 2022, 13, 915784. [Google Scholar] [CrossRef] [PubMed]
- Ronco, P.; Beck, L. Membranous nephropathy. Nat. Rev. Dis. Primers 2021, 7, 69. [Google Scholar] [CrossRef] [PubMed]
- Mella, A.; Deambrosis, I. Detection of urinary podocytes by flow cytometry in idiopathic membranous nephropathy. Sci. Rep. 2020, 10, 16362. [Google Scholar] [CrossRef]
- Wickman, L.; Afshinnia, F. Urine podocyte mRNAs, proteinuria, and progression in human glomerular diseases. J. Am. Soc. Nephrol. 2013, 24, 2081–2095. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, A.; Wickman, L.T. Angiotensin II-dependent persistent podocyte loss from destabilized glomeruli causes progression of end stage kidney disease. Kidney Int. 2012, 81, 40–55. [Google Scholar] [CrossRef] [Green Version]
- Sellarés, J.; de Freitas, D.G. Understanding the causes of kidney transplant failure: The dominant role of antibody-mediated rejection and nonadherence. Am. J. Transplant. 2012, 12, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Chauveau, B.; Raymond, A.A. The Proteome of Antibody-Mediated Rejection: From Glomerulitis to Transplant Glomerulopathy. Biomedicines 2022, 10, 569. [Google Scholar] [CrossRef] [PubMed]
- Lei, L.; Li, L.P. Value of urinary KIM-1 and NGAL combined with serum Cys C for predicting acute kidney injury secondary to decompensated cirrhosis. Sci. Rep. 2018, 8, 7962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomaru, A.; Kobayashi, T. Oligonucleotide-targeting periostin ameliorates pulmonary fibrosis. Gene Ther. 2017, 24, 706–716. [Google Scholar] [CrossRef]
- Kobayashi, T.; Kanno, K. Periostin antisense oligonucleotide prevents hepatic steatosis and fibrosis in a mouse model of non-alcoholic steatohepatitis. J. Gastroenterol. Hepatol. 2020, 35, 2140–2150. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pană, N.; Căpușă, C. Periostin as a Biomarker in the Setting of Glomerular Diseases—A Review of the Current Literature. Biomedicines 2022, 10, 3211. https://doi.org/10.3390/biomedicines10123211
Pană N, Căpușă C. Periostin as a Biomarker in the Setting of Glomerular Diseases—A Review of the Current Literature. Biomedicines. 2022; 10(12):3211. https://doi.org/10.3390/biomedicines10123211
Chicago/Turabian StylePană, Nicolae, and Cristina Căpușă. 2022. "Periostin as a Biomarker in the Setting of Glomerular Diseases—A Review of the Current Literature" Biomedicines 10, no. 12: 3211. https://doi.org/10.3390/biomedicines10123211