
The Anti-Inflammatory Effects of Iberin on TNF-α-Stimulated Human Oral Epithelial Cells: In Vitro Research

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Cytotoxicity Assay
2.3. IL-6 and CXCL10 Production in TR146 Cells
2.4. Western Blot Analysis
2.5. Statistical Analysis
3. Results
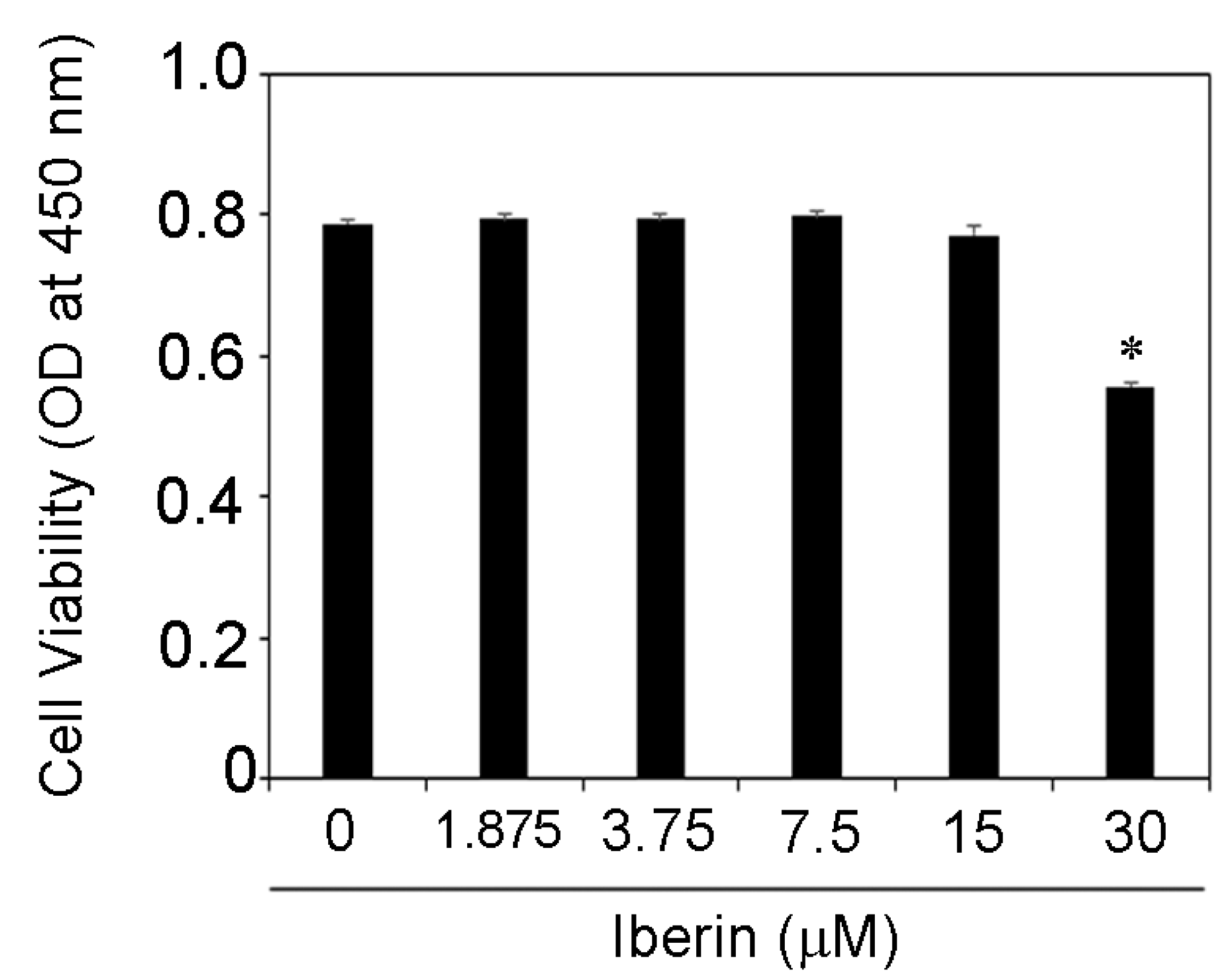
3.1. Effects of Iberin on Cell Viability of TR146 Cells
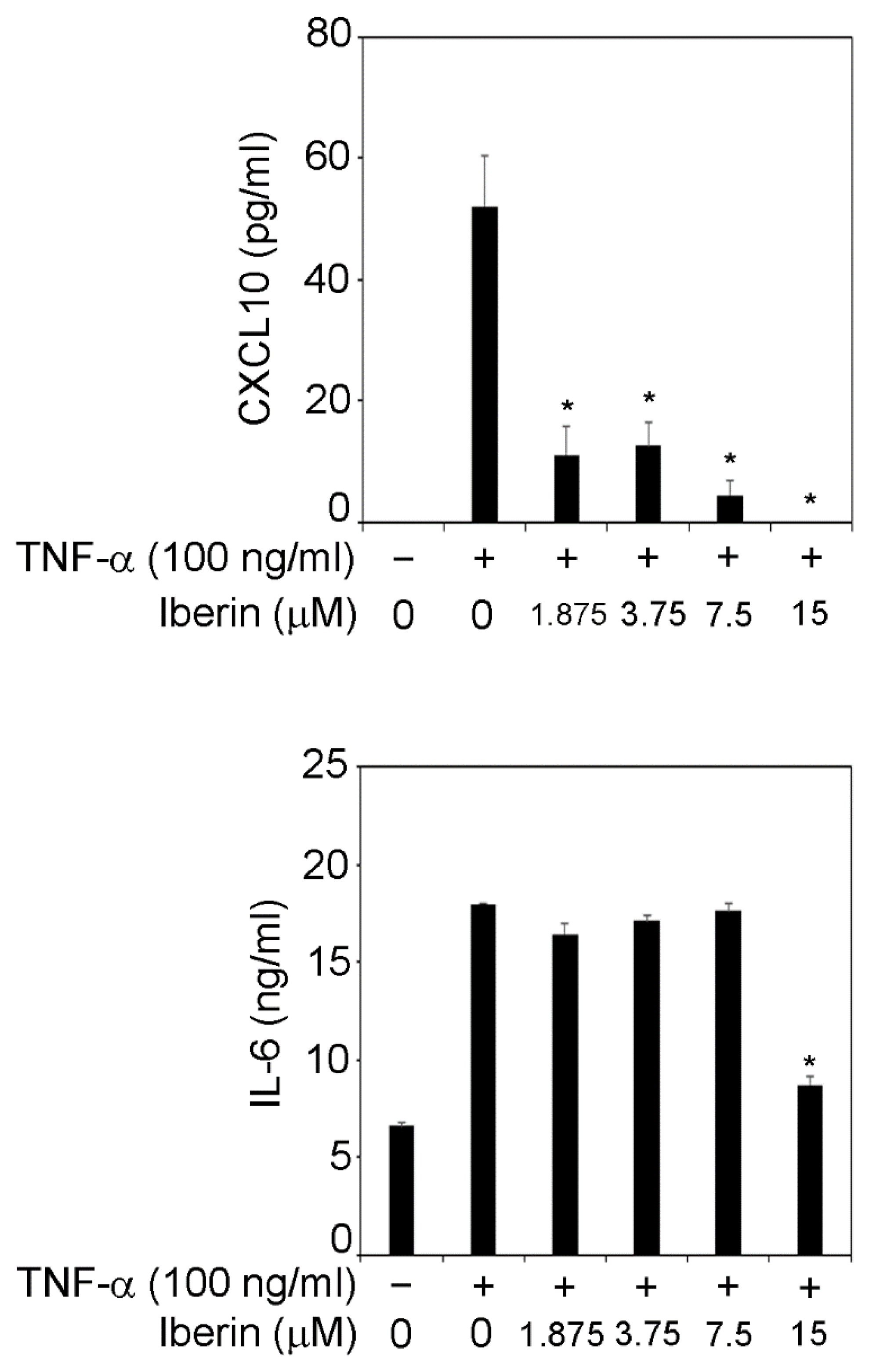
3.2. Iberin Suppresses TNF-α-Induced IL-6 and CXCL10 Production in TR146 Cells
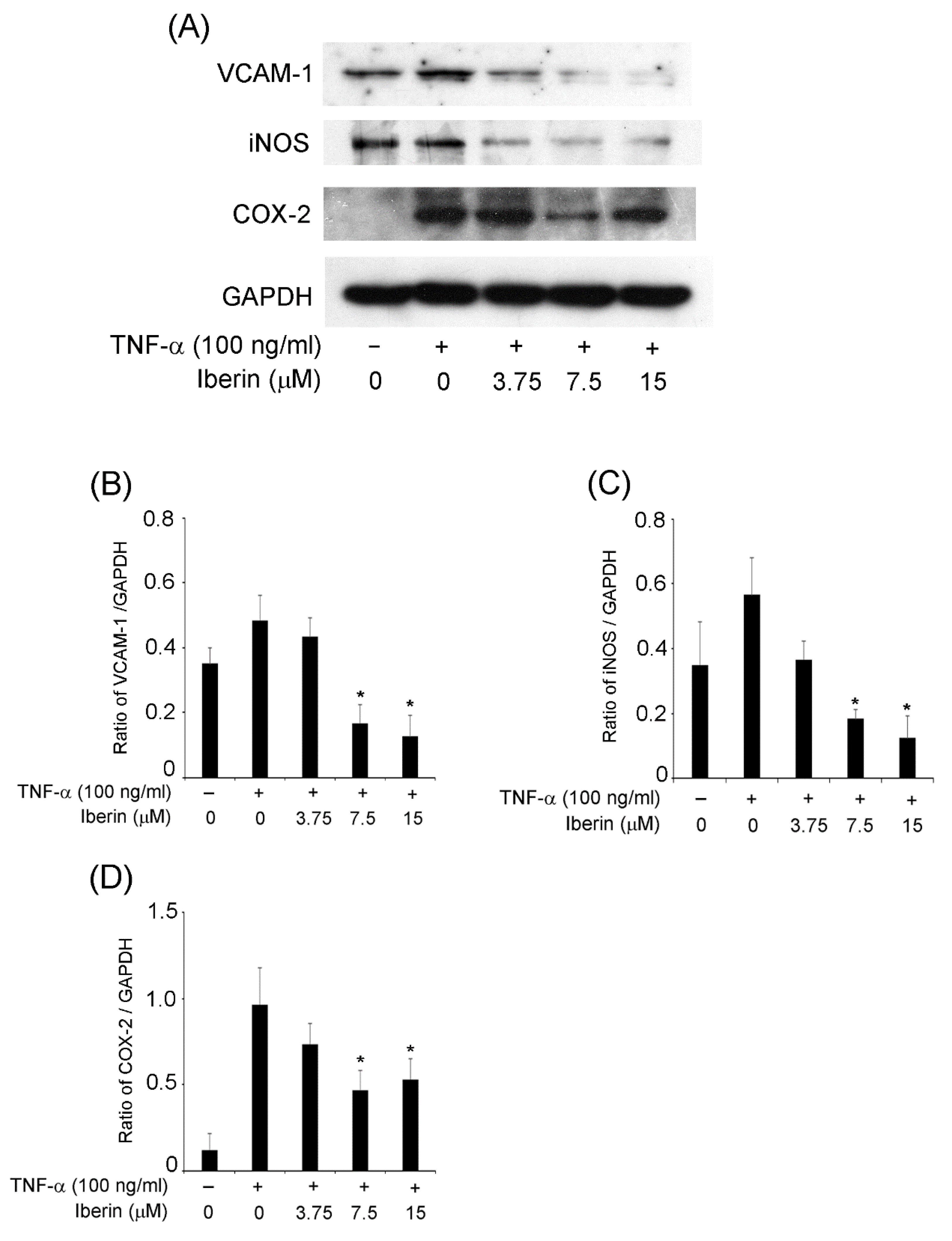
3.3. Iberin Inhibits VCAM-1, iNOS, and COX-2 Expression in TNF-α-Stimulated TR146 Cells
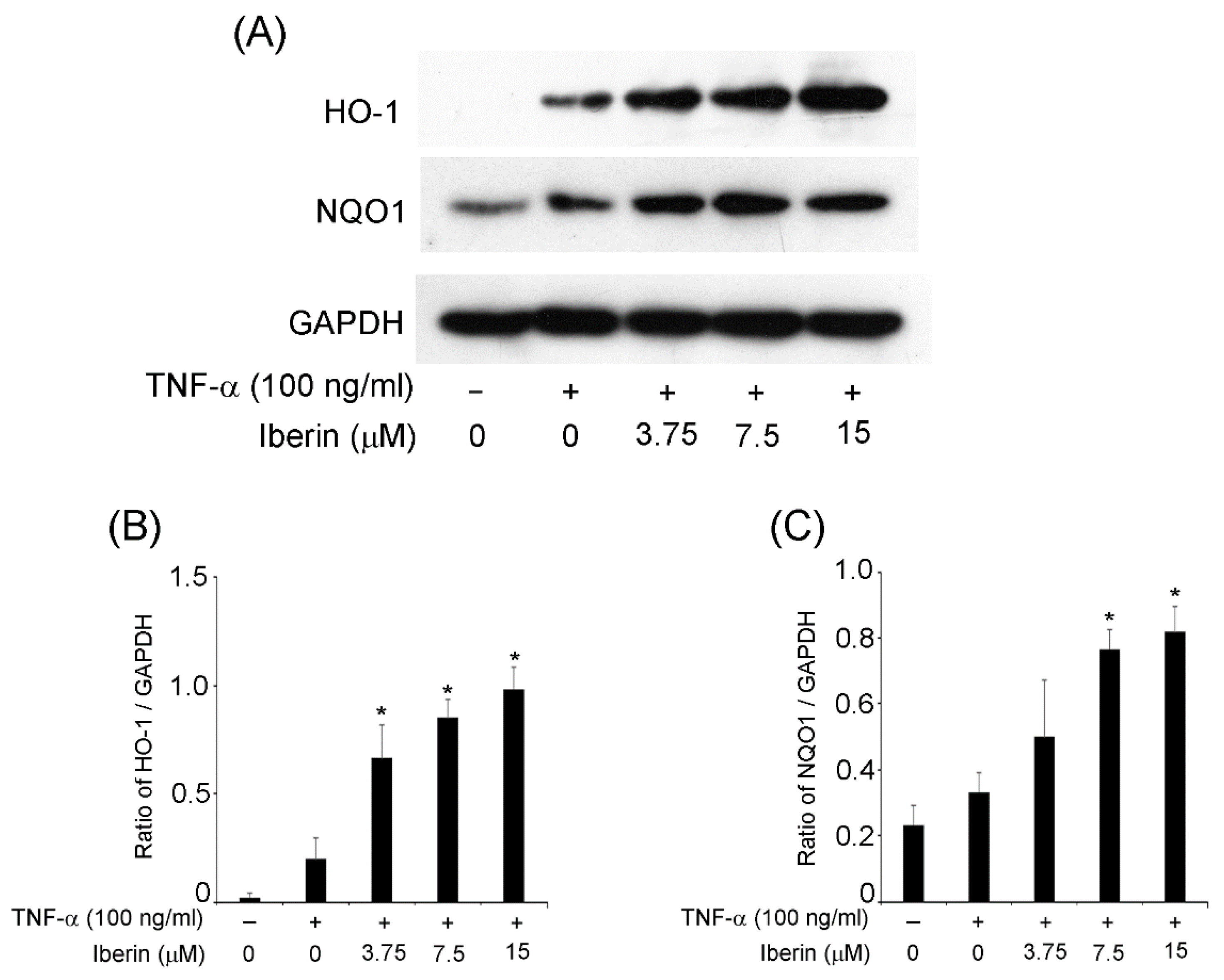
3.4. Effects of Iberin on HO-1 and NQO1 Expression in TR146 Cells
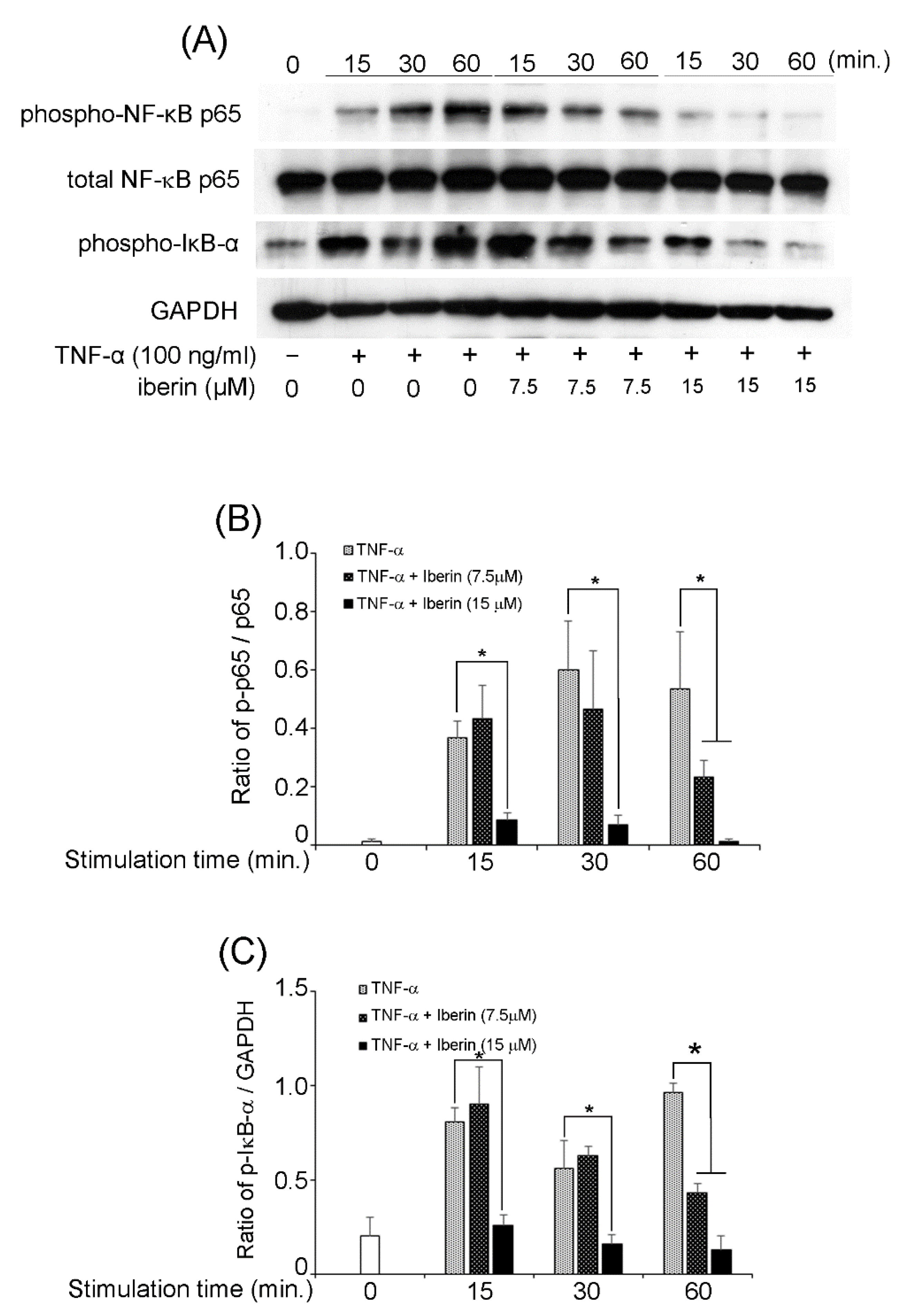
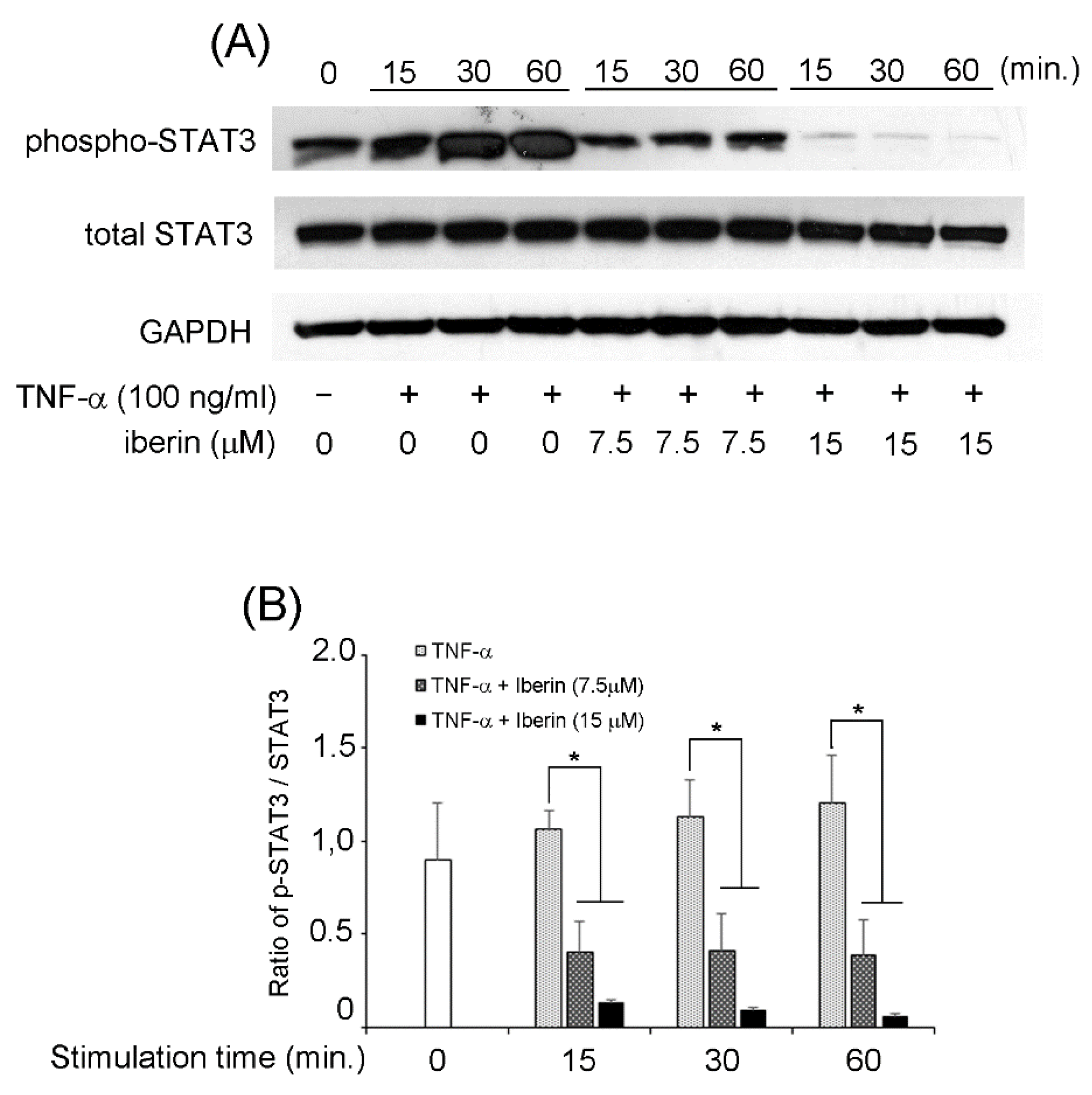
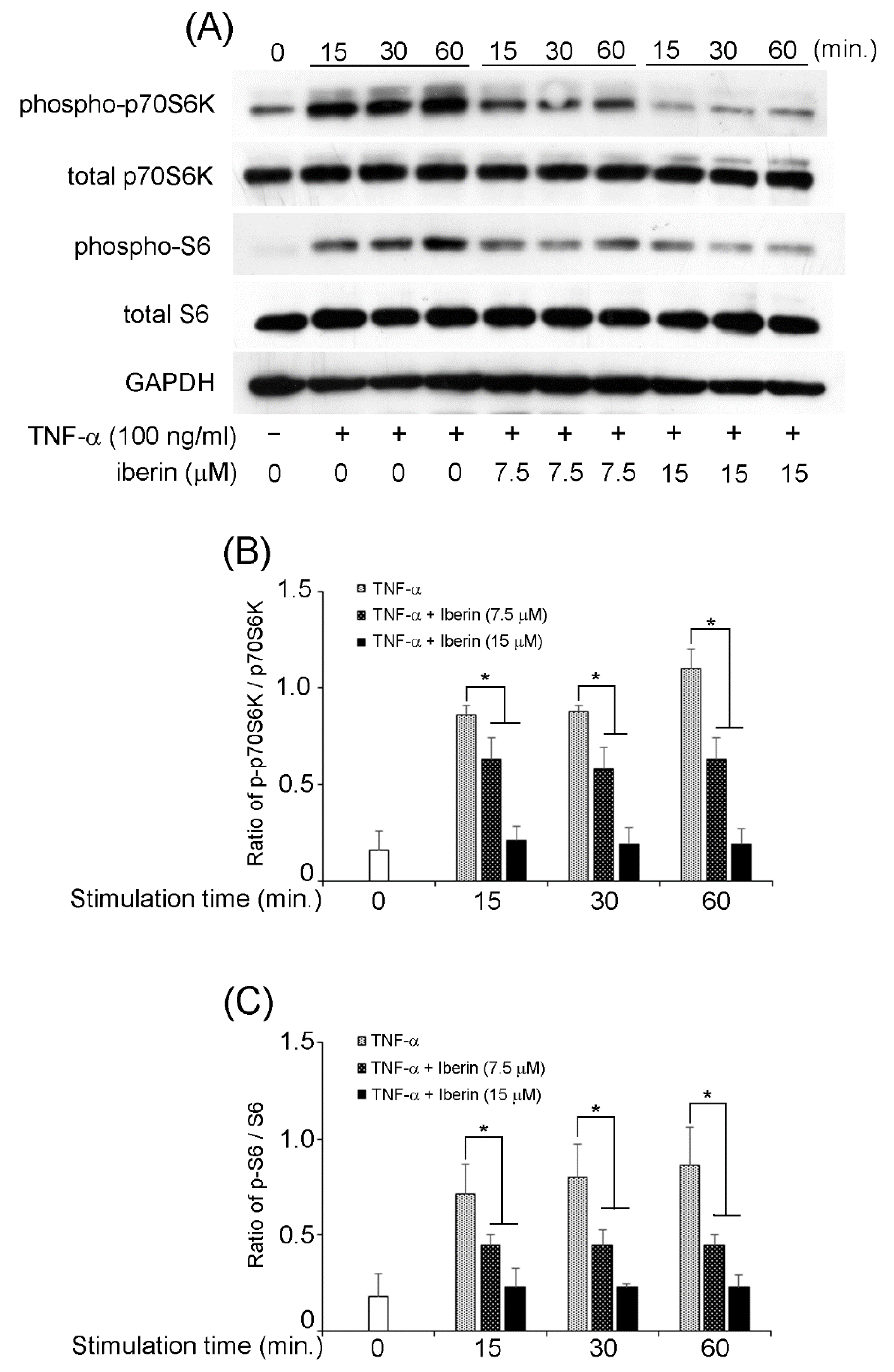
3.5. Effects of Iberin on NF-κB, STAT3, and p70S6K-S6 Signaling Pathways in TNF-α-Stimulated TR146 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Correction Statement
References
- Darveau, R.P. Periodontitis: A polymicrobial disruption of host homeostasis. Nat. Rev. Microbiol. 2010, 8, 481–490. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Cai, X.; Ou, Y.; Zhou, Y.; Wang, Y. Resolution of inflammation in periodontitis: A review. Int. J. Clin. Exp. Pathol. 2018, 11, 4283–4295. [Google Scholar]
- Mahuli, S.A.; Zorair, A.M.; Jafer, M.A.; Sultan, A.; Sarode, G.; Baeshen, H.A.; Raj, A.T.; Sarode, S.; Patil, S. Antibiotics for Periodontal Infections: Biological and Clinical Perspectives. J. Contemp. Dent. Pract. 2020, 21, 372–376. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y. The molecular basis that unifies the metabolism, cellular uptake and chemopreventive activities of dietary isothiocyanates. Carcinogenesis 2012, 33, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Huang, H.; Jin, L.; Lin, S. Anticarcinogenic Effects of Isothiocyanates on Hepatocellular Carcinoma. Int. J. Mol. Sci. 2022, 23, 13834. [Google Scholar] [CrossRef]
- Tarar, A.; Peng, S.; Cheema, S.; Peng, C.A. Anticancer Activity, Mechanism, and Delivery of Allyl Isothiocyanate. Bioengineering 2022, 9, 470. [Google Scholar] [CrossRef]
- Jadhav, U.; Ezhilarasan, R.; Vaughn, S.F.; Berhow, M.A.; Mohanam, S. Iberin induces cell cycle arrest and apoptosis in human neuroblastoma cells. Int. J. Mol. Med. 2007, 19, 353–361. [Google Scholar] [CrossRef]
- Gong, T.T.; Guo, Q.; Li, X.; Zhang, T.N.; Liu, F.H.; He, X.H.; Lin, B.; Wu, Q.J. Isothiocyanate Iberin inhibits cell proliferation and induces cell apoptosis in the progression of ovarian cancer by mediating ROS accumulation and GPX1 expression. Biomed. Pharmacother. 2021, 142, 111533. [Google Scholar] [CrossRef]
- Pocasap, P.; Weerapreeyakul, N.; Thumanu, K. Alyssin and Iberin in Cruciferous Vegetables Exert Anticancer Activity in HepG2 by Increasing Intracellular Reactive Oxygen Species and Tubulin Depolymerization. Biomol. Ther. 2019, 27, 540–552. [Google Scholar] [CrossRef]
- Suda, T.; Udagawa, N.; Nakamura, I.; Miyaura, C.; Takahashi, N. Modulation of osteoclast differentiation by local factors. Bone 1995, 17 (Suppl. 2), 87S–91S. [Google Scholar] [CrossRef]
- Cavalla, F.; Letra, A.; Silva, R.M.; Garlet, G.P. Determinants of Periodontal/Periapical Lesion Stability and Progression. J. Dent. Res. 2021, 100, 29–36. [Google Scholar] [CrossRef]
- Crawford, J.M.; Watanab, K. Cell adhesion molecules in inflammation and immunity: Relevance to periodontal diseases. Crit. Rev. Oral Biol. Med. 1994, 5, 91–123. [Google Scholar] [CrossRef]
- Uğar-Cankal, D.; Ozmeric, N. A multifaceted molecule, nitric oxide in oral and periodontal diseases. Clin. Chim. Acta 2006, 366, 90–100. [Google Scholar] [CrossRef]
- Fracon, R.N.; Teófilo, J.M.; Satin, R.B.; Lamano, T. Prostaglandins and bone: Potential risks and benefits related to the use of nonsteroidal anti-inflammatory drugs in clinical dentistry. J. Oral Sci. 2008, 50, 247–252. [Google Scholar] [CrossRef]
- Paul, O.; Arora, P.; Mayer, M.; Chatterjee, S. Inflammation in Periodontal Disease: Possible Link to Vascular Disease. Front. Physiol. 2021, 11, 609614. [Google Scholar] [CrossRef]
- Shimoyama, M.; Hosokawa, Y.; Hosokawa, I.; Ozaki, K.; Hosaka, K. 6-(Methylsulfinyl) Hexyl Isothiocyanate Inhibits IL-6 and CXCL10 Production in TNF-α-Stimulated Human Oral Epithelial Cells. Curr. Issues Mol. Biol. 2022, 44, 2915–2922. [Google Scholar] [CrossRef]
- Shibata, T.; Nakashima, F.; Honda, K.; Lu, Y.J.; Kondo, T.; Ushida, Y.; Aizawa, K.; Suganuma, H.; Oe, S.; Tanaka, H.; et al. Toll-like receptors as a target of food-derived anti-inflammatory compounds. J. Biol. Chem. 2014, 289, 32757–32772. [Google Scholar] [CrossRef]
- Thiruvengadam, M.; Venkidasamy, B.; Subramanian, U.; Samynathan, R.; Ali Shariati, M.; Rebezov, M.; Girish, S.; Thangavel, S.; Dhanapal, A.R.; Fedoseeva, N.; et al. Bioactive Compounds in Oxidative Stress-Mediated Diseases: Targeting the NRF2/ARE Signaling Pathway and Epigenetic Regulation. Antioxidants 2021, 10, 1859. [Google Scholar] [CrossRef]
- Wei, J.; Zhao, Q.; Zhang, Y.; Shi, W.; Wang, H.; Zheng, Z.; Meng, L.; Xin, Y.; Jiang, X. Sulforaphane-Mediated Nrf2 Activation Prevents Radiation-Induced Skin Injury through Inhibiting the Oxidative-Stress-Activated DNA Damage and NLRP3 Inflammasome. Antioxidants 2021, 10, 1850. [Google Scholar] [CrossRef]
- Ruhee, R.T.; Ma, S.; Suzuki, K. Protective Effects of Sulforaphane on Exercise-Induced Organ Damage via Inducing Antioxidant Defense Responses. Antioxidants 2020, 9, 136. [Google Scholar] [CrossRef]
- Ruhee, R.T.; Ma, S.; Suzuki, K. Sulforaphane Protects Cells against Lipopolysaccharide-Stimulated Inflammation in Murine Macrophages. Antioxidants 2019, 8, 577. [Google Scholar] [CrossRef] [PubMed]
- Ernst, I.M.; Palani, K.; Esatbeyoglu, T.; Schwarz, K.; Rimbach, G. Synthesis and Nrf2-inducing activity of the isothiocyanates iberverin, iberin and cheirolin. Pharmacol. Res. 2013, 70, 155–162. [Google Scholar] [CrossRef] [PubMed]
- Bahiraii, S.; Brenner, M.; Yan, F.; Weckwerth, W.; Heiss, E.H. Sulforaphane diminishes moonlighting of pyruvate kinase M2 and interleukin 1β expression in M1 (LPS) macrophages. Front. Immunol. 2022, 13, 935692. [Google Scholar] [CrossRef] [PubMed]
- Shawky, N.M.; Segar, L. Sulforaphane inhibits platelet-derived growth factor-induced vascular smooth muscle cell proliferation by targeting mTOR/p70S6kinase signaling independent of Nrf2 activation. Pharmacol. Res. 2017, 119, 251–264. [Google Scholar] [CrossRef]
- Qin, C.; Qin, Z.; Zhao, D.; Pan, Y.; Zhuang, L.; Wan, H.; Di Pizio, A.; Malach, E.; Niv, M.Y.; Huang, L.; et al. A bioinspired in vitro bioelectronic tongue with human T2R38 receptor for high-specificity detection of N-C=S-containing compounds. Talanta 2019, 199, 131–139. [Google Scholar] [CrossRef]
- Gil, S.; Coldwell, S.; Drury, J.L.; Arroyo, F.; Phi, T.; Saadat, S.; Kwong, D.; Chung, W.O. Genotype-specific regulation of oral innate immunity by T2R38 taste receptor. Mol. Immunol. 2015, 68 Pt. C, 663–670. [Google Scholar] [CrossRef]
- Medapati, M.R.; Singh, N.; Bhagirath, A.Y.; Duan, K.; Triggs-Raine, B.; Batista, E.L., Jr.; Chelikani, P. Bitter taste receptor T2R14 detects quorum sensing molecules from cariogenic Streptococcus mutans and mediates innate immune responses in gingival epithelial cells. FASEB J. 2021, 35, e21375. [Google Scholar] [CrossRef]
- Medapati, M.R.; Bhagirath, A.Y.; Singh, N.; Schroth, R.J.; Bhullar, R.P.; Duan, K.; Chelikani, P. Bitter Taste Receptor T2R14 Modulates Gram-Positive Bacterial Internalization and Survival in Gingival Epithelial Cells. Int. J. Mol. Sci. 2021, 22, 9920. [Google Scholar] [CrossRef]
- Xi, R.; Zheng, X.; Tizzano, M.J. Role of Taste Receptors in Innate Immunity and Oral Health. J. Dent. Res. 2022, 101, 759–768. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hosokawa, Y.; Hosokawa, I.; Shimoyama, M.; Fujii, A.; Sato, J.; Kadena, K.; Ozaki, K.; Hosaka, K. The Anti-Inflammatory Effects of Iberin on TNF-α-Stimulated Human Oral Epithelial Cells: In Vitro Research. Biomedicines 2022, 10, 3155. https://doi.org/10.3390/biomedicines10123155
Hosokawa Y, Hosokawa I, Shimoyama M, Fujii A, Sato J, Kadena K, Ozaki K, Hosaka K. The Anti-Inflammatory Effects of Iberin on TNF-α-Stimulated Human Oral Epithelial Cells: In Vitro Research. Biomedicines. 2022; 10(12):3155. https://doi.org/10.3390/biomedicines10123155
Chicago/Turabian StyleHosokawa, Yoshitaka, Ikuko Hosokawa, Masahiro Shimoyama, Ayumi Fujii, Juri Sato, Kimitake Kadena, Kazumi Ozaki, and Keiichi Hosaka. 2022. "The Anti-Inflammatory Effects of Iberin on TNF-α-Stimulated Human Oral Epithelial Cells: In Vitro Research" Biomedicines 10, no. 12: 3155. https://doi.org/10.3390/biomedicines10123155