

Milmed Yeast Alters the LPS-Induced M1 Microglia Cells to Form M2 Anti-Inflammatory Phenotype
 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Growth
2.2. Cell Culture and Treatment
2.3. Cell Viability Assays
2.3.1. Trypan Blue Exclusion Assay
2.3.2. Immunofluorescence Microscopy
2.4. Real-Time Quantitative PRC Analysis
2.5. Statistical Analysis
3. Results
3.1. BV-2 Cellular Areas
3.2. iNOS and Arg-1 Expression
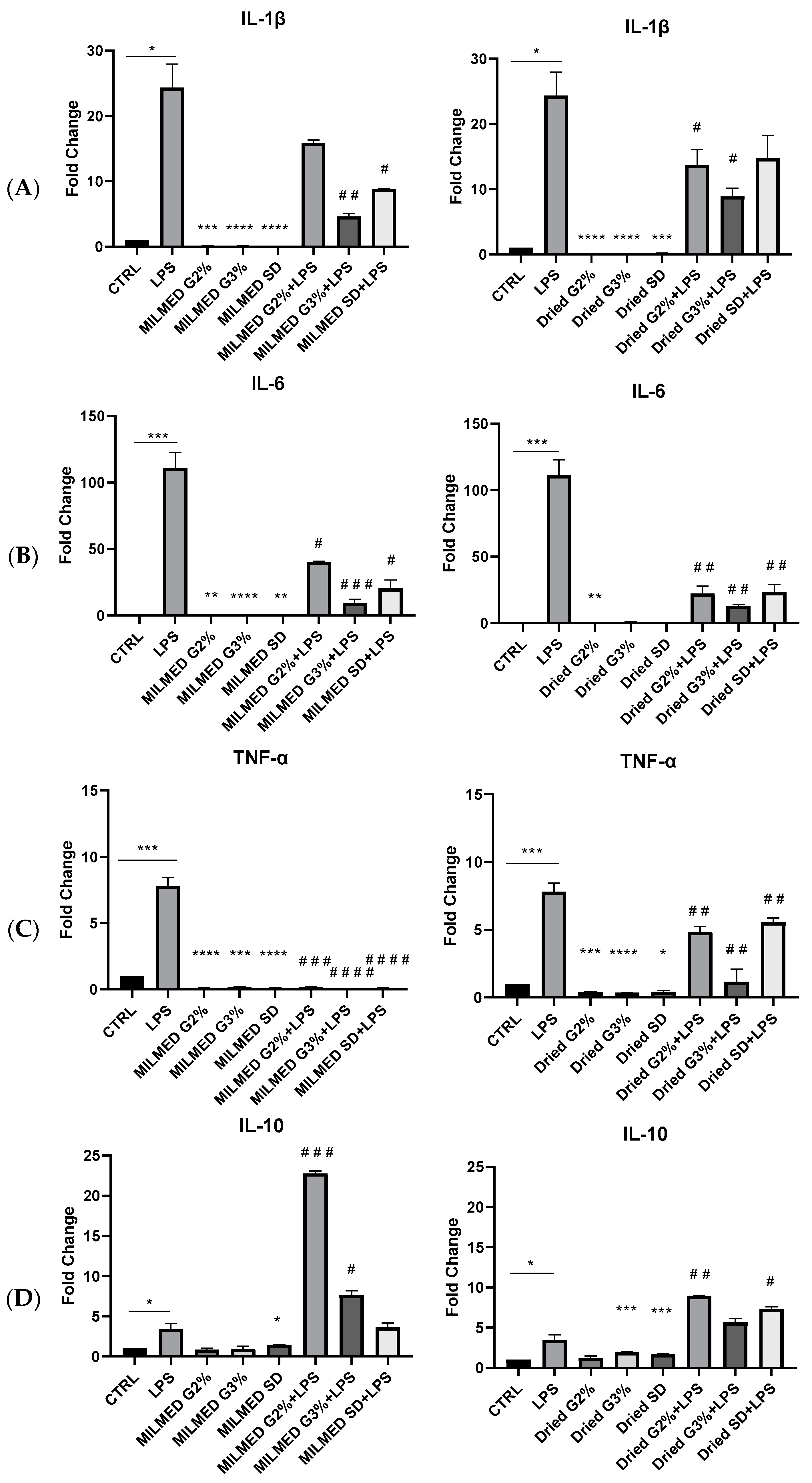
3.3. Cytokines Expression
4. Discussion
5. Conclusions
Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- El Dib, R.; Periyasamy, A.G.; de Barros, J.L.; França, C.G.; Senefonte, F.L.; Vesentini, G.; Alves, M.G.O.; Rodrigues, J.V.D.S.; Gomaa, H.; Júnior, J.R.G.; et al. Probiotics for the Treatment of Depression and Anxiety: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. Clin. Nutr. ESPEN 2021, 45, 75–90. [Google Scholar] [CrossRef] [PubMed]
- Maggi, E.; Armeli, F.; Mengoni, B.; Leo, M.; Filetici, P.; Mancini, P.; Lenz, T.; Businaro, R.; Archer, T. Milmed Saccharomyces cerevisiae Activity on Central Nervous System Cells. J. Toxicol. Pharmacol. 2022, 5, 26. [Google Scholar]
- Shabbir, U.; Arshad, M.S.; Sameen, A.; Oh, D.-H. Crosstalk between Gut and Brain in Alzheimer’s Disease: The Role of Gut Microbiota Modulation Strategies. Nutrients 2021, 13, 690. [Google Scholar] [CrossRef] [PubMed]
- Eltokhi, A.; Sommer, I.E. A Reciprocal Link Between Gut Microbiota, Inflammation and Depression: A Place for Probiotics? Front. Neurosci. 2022, 16, 852506. [Google Scholar] [CrossRef]
- Grieco, M.; De Caris, M.G.; Maggi, E.; Armeli, F.; Coccurello, R.; Bisogno, T.; D’Erme, M.; Maccarrone, M.; Mancini, P.; Businaro, R. Fatty Acid Amide Hydrolase (FAAH) Inhibition Modulates Amyloid-Beta-Induced Microglia Polarization. Int. J. Mol. Sci. 2021, 22, 7711. [Google Scholar] [CrossRef]
- Neta, F.I.; de Souza, F.E.S.; Batista, A.L.; Pinheiro, F.I.; Cobucci, R.N.; Guzen, F.P. Effects of Supplementation with Probiotics in Experimental Models OfAlzheimer’s Disease: A Systematic Review of Animal Experiments. Curr. Alzheimer Res. 2022, 19, 188–201. [Google Scholar] [CrossRef]
- Li, K.; Ly, K.; Mehta, S.; Braithwaite, A. Importance of crosstalk between the microbiota and the neuroimmune system for tissue homeostasis. Clin. Transl. Immunol. 2022, 11, e1394. [Google Scholar] [CrossRef]
- Varela-Trinidad, G.U.; Domínguez-Díaz, C.; Solórzano-Castanedo, K.; Íñiguez-Gutiérrez, L.; Hernández-Flores, T.D.J.; Fafutis-Morris, M. Probiotics: Protecting Our Health from the Gut. Microorganisms 2022, 10, 1428. [Google Scholar] [CrossRef]
- Caputi, V.; Giron, M.C. Microbiome-Gut-Brain Axis and Toll-Like Receptors in Parkinson’s Disease. Int. J. Mol. Sci. 2018, 19, 1689. [Google Scholar] [CrossRef] [Green Version]
- Zhuang, Z.-Q.; Shen, L.-L.; Li, W.-W.; Fu, X.; Zeng, F.; Gui, L.; Lü, Y.; Cai, M.; Zhu, C.; Tan, Y.-L.; et al. Gut Microbiota Is Altered in Patients with Alzheimer’s Disease. J. Alzheimer’s Dis. 2018, 63, 1337–1346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, Y.; Pedersen, O. Gut Microbiota in Human Metabolic Health and Disease. Nat. Rev. Microbiol. 2021, 19, 55–71. [Google Scholar] [CrossRef] [PubMed]
- Piekut, T.; Hurła, M.; Banaszek, N.; Szejn, P.; Dorszewska, J.; Kozubski, W.; Prendecki, M. Infectious Agents and Alzheimer’s Disease. J. Integr. Neurosci. 2022, 21, 073. [Google Scholar] [CrossRef] [PubMed]
- Webberley, T.S.; Masetti, G.; Bevan, R.J.; Kerry-Smith, J.; Jack, A.A.; Michael, D.R.; Thomas, S.; Glymenaki, M.; Li, J.; McDonald, J.A.K.; et al. The Impact of Probiotic Supplementation on Cognitive, Pathological and Metabolic Markers in a Transgenic Mouse Model of Alzheimer’s Disease. Front. Neurosci. 2022, 16, 843105. [Google Scholar] [CrossRef] [PubMed]
- Ghannoum, M.A.; McCormick, T.S.; Retuerto, M.; Bebek, G.; Cousineau, S.; Hartman, L.; Barth, C.; Schrom, K. Evaluation of Microbiome Alterations Following Consumption of BIOHM, a Novel Probiotic. Curr. Issues Mol. Biol. 2021, 43, 2135–2146. [Google Scholar] [CrossRef] [PubMed]
- Birmann, P.T.; Casaril, A.M.; Pesarico, A.P.; Caballero, P.S.; Smaniotto, T.Â.; Rodrigues, R.R.; Moreira, Â.N.; Conceição, F.R.; Sousa, F.S.S.; Collares, T.; et al. Komagataella Pastoris KM71H Modulates Neuroimmune and Oxidative Stress Parameters in Animal Models of Depression: A Proposal for a New Probiotic with Antidepressant-like Effect. Pharmacol. Res. 2021, 171, 105740. [Google Scholar] [CrossRef]
- Durmaz, S.; Kurtoğlu, T.; Barbarus, E.; Çetin, N.K.; Yılmaz, M.; Rahman, F.; Abacıgil, F. Probiotic Saccharomyces boulardii Alleviates Lung Injury by Reduction of Oxidative Stress and Cytokine Response Induced by Supraceliac Aortic Ischemia-Reperfusion Injury in Rats. Braz. J. Cardiovasc. Surg. 2020, 36, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Roy Sarkar, S.; Mitra Mazumder, P.; Chatterjee, K.; Sarkar, A.; Adhikary, M.; Mukhopadhyay, K.; Banerjee, S. Saccharomyces Boulardii Ameliorates Gut Dysbiosis Associated Cognitive Decline. Physiol. Behav. 2021, 236, 113411. [Google Scholar] [CrossRef] [PubMed]
- Rahmani, B.; Alimadadi, N.; Attaran, B.; Nasr, S. Yeasts from Iranian Traditional Milk Kefir Samples: Isolation, Molecular Identification and Their Potential Probiotic Properties. Lett. Appl. Microbiol. 2022, 75, 1264–1274. [Google Scholar] [CrossRef]
- Ye, T.; Yuan, S.; Kong, Y.; Yang, H.; Wei, H.; Zhang, Y.; Jin, H.; Yu, Q.; Liu, J.; Chen, S.; et al. Effect of Probiotic Fungi against Cognitive Impairment in Mice via Regulation of the Fungal Microbiota–Gut–Brain Axis. J. Agric. Food Chem. 2022, 70, 9026–9038. [Google Scholar] [CrossRef]
- Archer, T. Milmed Treatment Alleviates Symptoms of Allergy and Improves General Health. J. Immunol. Allergy 2021, 1, 1–11. [Google Scholar] [CrossRef]
- Prokhorov, A.M.; Dianov, E.M. In Memory of Mikhail B. Golan. Tech. Phys. 2001, 46, 1068. [Google Scholar] [CrossRef]
- Betskii, O.V.; Devyatkov, N.D.; Kislov, V.V. Low Intensity Millimeter Waves in Medicine and Biology. Crit. Rev. Biomed. Eng. 2000, 28, 247–268. [Google Scholar] [CrossRef] [PubMed]
- Devyatkov, N.; Golant, M.; Betsky, O. Brief Information for Physicians about the Physical Characteristics of the Processes Occurring in the Body under MM-Wave Therapy Performed by Installations “Jav-1”, and the Associated Effects on the Body of Electromagnetic Millimeter Waves; Radio i Svyaz: Moscow, Russia, 1991. [Google Scholar]
- Devyatkov, N.D.; Gelvich, É.A.; Davydova, I.B.; Kirillov, V.V.; Kolmakov, D.N.; Mazokhin, V.N.; Sinyagovskiĭ, V.I.; Chilikin, P.I. Microwave and radio-frequency apparatus and methods for use in oncology. Sov. Phys. Uspekhi 1981, 24, 432. [Google Scholar] [CrossRef]
- Golant, M.B.; Mudrik, D.G.; Kruglyakova, O.P.; Izvol’skaya, V.E. Effect of EHF-Radiation Polarization on Yeast Cells. Radiophys. Quantum Electron. 1994, 37, 82–84. [Google Scholar] [CrossRef]
- Golant, M.B. Physical Laws of Medicine and Their Use in the Realization of Interaction of Living Organisms with EHF Radiation. Radiophys. Quantum Electron. 1994, 37, 45–47. [Google Scholar] [CrossRef]
- Ragimov, C.R.; Ter-Asaturov, G.P.; Golant, M.B.; Rogov, K.A.; Balakireva, L.Z. Stimulation of Reparative Osteogenesis by Millimeter Band Electromagnetic Radiation in Experimental Traumatic Defects of the Mandible. Bull. Exp. Biol. Med. 1991, 111, 562–565. [Google Scholar] [CrossRef]
- Archer, T.; Fredriksson, A. The Yeast Product Milmed Enhances the Effect of Physical Exercise on Motor Performance and Dopamine Neurochemistry Recovery in MPTP-Lesioned Mice. Neurotox. Res. 2013, 24, 393–406. [Google Scholar] [CrossRef]
- Orihuela, R.; McPherson, C.A.; Harry, G.J. Microglial M1/M2 Polarization and Metabolic States: Microglia Bioenergetics with Acute Polarization. Br. J. Pharmacol. 2016, 173, 649–665. [Google Scholar] [CrossRef] [Green Version]
- Chidambaram, S.B.; Essa, M.M.; Rathipriya, A.G.; Bishir, M.; Ray, B.; Mahalakshmi, A.M.; Tousif, A.H.; Sakharkar, M.K.; Kashyap, R.S.; Friedland, R.P.; et al. Gut Dysbiosis, Defective Autophagy and Altered Immune Responses in Neurodegenerative Diseases: Tales of a Vicious Cycle. Pharmacol. Ther. 2022, 231, 107988. [Google Scholar] [CrossRef]
- Schroeder, B.O.; Bäckhed, F. Signals from the Gut Microbiota to Distant Organs in Physiology and Disease. Nat. Med. 2016, 22, 1079–1089. [Google Scholar] [CrossRef]
- Goyal, D.; Ali, S.A.; Singh, R.K. Emerging Role of Gut Microbiota in Modulation of Neuroinflammation and Neurodegeneration with Emphasis on Alzheimer’s Disease. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 106, 110112. [Google Scholar] [CrossRef] [PubMed]
- Kowalski, K.; Mulak, A. Brain-Gut-Microbiota Axis in Alzheimer’s Disease. J. Neurogastroenterol. Motil. 2019, 25, 48–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cherry, J.D.; Olschowka, J.A.; O’Banion, M.K. Neuroinflammation and M2 Microglia: The Good, the Bad, and the Inflamed. J. Neuroinflamm. 2014, 11, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, Y.; Le, W. Differential Roles of M1 and M2 Microglia in Neurodegenerative Diseases. Mol. Neurobiol. 2016, 53, 1181–1194. [Google Scholar] [CrossRef]
- Dubbelaar, M.L.; Kracht, L.; Eggen, B.J.L.; Boddeke, E.W.G.M. The Kaleidoscope of Microglial Phenotypes. Front. Immunol. 2018, 9, 1753. [Google Scholar] [CrossRef]
- Jurga, A.M.; Paleczna, M.; Kuter, K.Z. Overview of General and Discriminating Markers of Differential Microglia Phenotypes. Front. Cell. Neurosci. 2020, 14, 198. [Google Scholar] [CrossRef]
- Kwon, H.S.; Koh, S.-H. Neuroinflammation in Neurodegenerative Disorders: The Roles of Microglia and Astrocytes. Transl. Neurodegener. 2020, 9, 42. [Google Scholar] [CrossRef]
- Palepu, M.S.K.; Dandekar, M.P. Remodeling of Microbiota Gut-Brain Axis Using Psychobiotics in Depression. Eur. J. Pharmacol. 2022, 931, 175171. [Google Scholar] [CrossRef]
- Rahman, Z.; Dandekar, M.P. Crosstalk between Gut Microbiome and Immunology in the Management of Ischemic Brain Injury. J. Neuroimmunol. 2021, 353, 577498. [Google Scholar] [CrossRef]
- Chen, Z.; Maqbool, J.; Sajid, F.; Hussain, G.; Sun, T. Human Gut Microbiota and Its Association with Pathogenesis and Treatments of Neurodegenerative Diseases. Microb. Pathog. 2021, 150, 104675. [Google Scholar] [CrossRef]
- Westfall, S.; Pasinetti, G.M. The Gut Microbiota Links Dietary Polyphenols With Management of Psychiatric Mood Disorders. Front. Neurosci. 2019, 13, 1196. [Google Scholar] [CrossRef] [PubMed]
- Rahimian, R.; Belliveau, C.; Chen, R.; Mechawar, N. Microglial Inflammatory-Metabolic Pathways and Their Potential Therapeutic Implication in Major Depressive Disorder. Front. Psychiatry 2022, 13, 871997. [Google Scholar] [CrossRef] [PubMed]
- Princiotta Cariddi, L.; Mauri, M.; Cosentino, M.; Versino, M.; Marino, F. Alzheimer’s Disease: From Immune Homeostasis to Neuroinflammatory Condition. Int. J. Mol. Sci. 2022, 23, 13008. [Google Scholar] [CrossRef]
- Choi, I.; Heaton, G.R.; Lee, Y.-K.; Yue, Z. Regulation of α-Synuclein Homeostasis and Inflammasome Activation by Microglial Autophagy. Sci. Adv. 2022, 8, eabn1298. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.; Zhenxin, Y.; Chen, S.; Tan, Z.; Zong, Z.; Zhang, H.; Xiong, X. The Innate and Adaptive Immune Cells in Alzheimer’s and Parkinson’s Diseases. Oxidative Med. Cell. Longev. 2022, 2022, 1315248. [Google Scholar] [CrossRef] [PubMed]
- Araújo, B.; Caridade-Silva, R.; Soares-Guedes, C.; Martins-Macedo, J.; Gomes, E.D.; Monteiro, S.; Teixeira, F.G. Neuroinflammation and Parkinson’s Disease—From Neurodegeneration to Therapeutic Opportunities. Cells 2022, 11, 2908. [Google Scholar] [CrossRef] [PubMed]
- Thakur, S.; Dhapola, R.; Sarma, P.; Medhi, B.; Reddy, D.H. Neuroinflammation in Alzheimer’s Disease: Current Progress in Molecular Signaling and Therapeutics. Inflammation 2022, 1–17. [Google Scholar] [CrossRef]
- Sarkar, A.; Lehto, S.M.; Harty, S.; Dinan, T.G.; Cryan, J.F.; Burnet, P.W.J. Psychobiotics and the Manipulation of Bacteria–Gut–Brain Signals. Trends Neurosci. 2016, 39, 763–781. [Google Scholar] [CrossRef] [Green Version]
- Blander, J.M.; Longman, R.S.; Iliev, I.D.; Sonnenberg, G.F.; Artis, D. Regulation of Inflammation by Microbiota Interactions with the Host. Nat. Immunol. 2017, 18, 851–860. [Google Scholar] [CrossRef]
- Chen, Y.; Peng, F.; Xing, Z.; Chen, J.; Peng, C.; Li, D. Beneficial Effects of Natural Flavonoids on Neuroinflammation. Front. Immunol. 2022, 13, 1006434. [Google Scholar] [CrossRef]
- Marć, M.A.; Jastrząb, R.; Mytych, J. Does the Gut Microbial Metabolome Really Matter? The Connection between GUT Metabolome and Neurological Disorders. Nutrients 2022, 14, 3967. [Google Scholar] [CrossRef] [PubMed]
- Zhu, G.; Zhao, J.; Zhang, H.; Chen, W.; Wang, G. Administration of Bifidobacterium Breve Improves the Brain Function of Aβ1-42-Treated Mice via the Modulation of the Gut Microbiome. Nutrients 2021, 13, 1602. [Google Scholar] [CrossRef] [PubMed]
- Akbari, E.; Asemi, Z.; Daneshvar Kakhaki, R.; Bahmani, F.; Kouchaki, E.; Tamtaji, O.R.; Hamidi, G.A.; Salami, M. Effect of probiotic supplementation on cognitive function and metabolic status in Alzheimer’s disease: A randomized, double-blind and controlled trial. Front. Aging Neurosci. 2016, 8, 256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gur, T.L.; Bailey, M.T. Effects of Stress on Commensal Microbes and Immune System Activity. In Microbial Endocrinology: Interkingdom Signaling in Infectious Disease and Health; Lyte, M., Ed.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2016; Volume 874, pp. 289–300. ISBN 978-3-319-20214-3. [Google Scholar]
- Archer, T.; Garcia, D.; Fredriksson, A. Restoration of MPTP-Induced Deficits by Exercise and Milmed® Co-Treatment. PeerJ 2014, 2, e531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, X.; Xie, Q.; Xu, H.; Zhang, T.; Li, X.; Tian, Y.; Lan, H.; Kong, L.; Zhang, Z. Yeast Microcapsule Mediated Natural Products Delivery for Treating Ulcerative Colitis through Anti-Inflammatory and Regulation of Macrophage Polarization. ACS Appl. Mater. Interfaces 2022, 14, 31085–31098. [Google Scholar] [CrossRef]
- Taylor, P.R.; Tsoni, S.V.; Willment, J.A.; Dennehy, K.M.; Rosas, M.; Findon, H.; Haynes, K.; Steele, C.; Botto, M.; Gordon, S.; et al. Dectin-1 Is Required for β-Glucan Recognition and Control of Fungal Infection. Nat. Immunol. 2007, 8, 31–38. [Google Scholar] [CrossRef]
- Ranuh, R.; Athiyyah, A.F.; Darma, A.; Risky, V.P.; Riawan, W.; Surono, I.S.; Sudarmo, S.M. Effect of the Probiotic Lactobacillus Plantarum IS-10506 on BDNF and 5HT Stimulation: Role of Intestinal Microbiota on the Gut-Brain Axis. Iran. J. Microbiol. 2019, 11, 145–150. [Google Scholar] [CrossRef]
- Businaro, R.; Vauzour, D.; Sarris, J.; Münch, G.; Gyengesi, E.; Brogelli, L.; Zuzarte, P. Therapeutic Opportunities for Food Supplements in Neurodegenerative Disease and Depression. Front. Nutr. 2021, 8, 669846. [Google Scholar] [CrossRef]
- De Caris, M.G.; Grieco, M.; Maggi, E.; Francioso, A.; Armeli, F.; Mosca, L.; Pinto, A.; D’Erme, M.; Mancini, P.; Businaro, R. Blueberry Counteracts BV-2 Microglia Morphological and Functional Switch after LPS Challenge. Nutrients 2020, 12, 1830. [Google Scholar] [CrossRef]
- Angeloni, C.; Businaro, R.; Vauzour, D. The Role of Diet in Preventing and Reducing Cognitive Decline. Curr. Opin. Psychiatry 2020, 33, 432–438. [Google Scholar] [CrossRef]
- Businaro, R.; Corsi, M.; Asprino, R.; Di Lorenzo, C.; Laskin, D.; Corbo, R.M.; Ricci, S.; Pinto, A. Modulation of Inflammation as a Way of Delaying Alzheimer’s Disease Progression: The Diet’s Role. Curr. Alzheimer Res. 2018, 15, 363–380. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhao, H.; Wang, L.; Tao, Y.; Du, G.; Guan, W.; Liu, J.; Brennan, C.; Ho, C.-T.; Li, S. Effects of Selected Resveratrol Analogues on Activation and Polarization of Lipopolysaccharide-Stimulated BV-2 Microglial Cells. J. Agric. Food Chem. 2020, 68, 3750–3757. [Google Scholar] [CrossRef] [PubMed]
- Buttari, B.; Profumo, E.; Segoni, L.; D’Arcangelo, D.; Rossi, S.; Facchiano, F.; Saso, L.; Businaro, R.; Iuliano, L.; Riganò, R. Resveratrol Counteracts Inflammation in Human M1 and M2 Macrophages upon Challenge with 7-Oxo-Cholesterol: Potential Therapeutic Implications in Atherosclerosis. Oxidative Med. Cell. Longev. 2014, 2014, 257543. [Google Scholar] [CrossRef] [Green Version]
- Román, G.C.; Jackson, R.E.; Gadhia, R.; Román, A.N.; Reis, J. Mediterranean Diet: The Role of Long-Chain ω-3 Fatty Acids in Fish; Polyphenols in Fruits, Vegetables, Cereals, Coffee, Tea, Cacao and Wine; Probiotics and Vitamins in Prevention of Stroke, Age-Related Cognitive Decline, and Alzheimer Disease. Rev. Neurol. 2019, 175, 724–741. [Google Scholar] [CrossRef] [PubMed]
- Pinto, A.; Bonucci, A.; Maggi, E.; Corsi, M.; Businaro, R. Anti-Oxidant and Anti-Inflammatory Activity of Ketogenic Diet: New Perspectives for Neuroprotection in Alzheimer’s Disease. Antioxidants 2018, 7, 63. [Google Scholar] [CrossRef] [Green Version]
- Versele, R.; Corsi, M.; Fuso, A.; Sevin, E.; Businaro, R.; Gosselet, F.; Fenart, L.; Candela, P. Ketone Bodies Promote Amyloid-Β1–40 Clearance in a Human in Vitro Blood–Brain Barrier Model. Int. J. Mol. Sci. 2020, 21, 934. [Google Scholar] [CrossRef] [Green Version]
- Lazo-Vélez, M.A.; Serna-Saldívar, S.O.; Rosales-Medina, M.F.; Tinoco-Alvear, M.; Briones-García, M. Application of Saccharomyces cerevisiae var. boulardii in Food Processing: A Review. J. Appl. Microbiol. 2018, 125, 943–951. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.E.; Lee, E. The Probiotic Effects of the Saccharomyces cerevisiae 28-7 Strain Isolated from Nuruk in a DSS-Induced Colitis Mouse Model. J. Microbiol. Biotechnol. 2022, 32, 877–884. [Google Scholar] [CrossRef]
- Pothoulakis, C. Review Article: Anti-Inflammatory Mechanisms of Action of Saccharomyces boulardii. Aliment. Pharmacol. Ther. 2009, 30, 826–833. [Google Scholar] [CrossRef]
- Wang, C.; Li, W.; Wang, H.; Ma, Y.; Zhao, X.; Zhang, X.; Yang, H.; Qian, J.; Li, J. Saccharomyces boulardii Alleviates Ulcerative Colitis Carcinogenesis in Mice by Reducing TNF-α and IL-6 Levels and Functions and by Rebalancing Intestinal Microbiota. BMC Microbiol. 2019, 19, 246. [Google Scholar] [CrossRef]
- Archer, T. Anti-Inflammatory Action of the Treated-Yeast, Milmed, Under IBS-IBD Conditions. J. Immunol. Allergy 2022, 3. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′–3′) | Reverse Primer (5′–3′) | Accession Numbers |
| mIL-1β | GAAATGCCACCTTTTGACAGTG | TGGATGCTCTCATCAGGACAG | NM_008361.4 |
| mTNF-α | CTGAACTTCGGGGTGATCGG | GGCTTGTCACTCGAATTTTGAGA | BC137720.1 |
| mIL-10 | GCCCTTTGCTATGGTGTCCTTTC | TCCCTGGTTTCTCTTCCCAAGAC | NM_010548.2 |
| mARG1 | ATGTGCCCTCTGTCTTTTAGGG | GGTCTCTCACGTCATACTCTGT | NM_007482.3 |
| miNOS | GGCAGCCTGTGAGACCTTTG | GCATTGGAAGTGAAGCGTTTC | AF427516.1 |
| mIL-6 | CGGAGAGGAGACTTCACAGAGGA | TTTCCACGATTTCCCAGAGAACA | NM_001314054.1 |
| mACT-β | GGCTGTATTCCCCTCCATCG | CCAGTTGGTAACAATGCCATGT | NM_007393.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Armeli, F.; Mengoni, B.; Maggi, E.; Mazzoni, C.; Preziosi, A.; Mancini, P.; Businaro, R.; Lenz, T.; Archer, T. Milmed Yeast Alters the LPS-Induced M1 Microglia Cells to Form M2 Anti-Inflammatory Phenotype. Biomedicines 2022, 10, 3116. https://doi.org/10.3390/biomedicines10123116
Armeli F, Mengoni B, Maggi E, Mazzoni C, Preziosi A, Mancini P, Businaro R, Lenz T, Archer T. Milmed Yeast Alters the LPS-Induced M1 Microglia Cells to Form M2 Anti-Inflammatory Phenotype. Biomedicines. 2022; 10(12):3116. https://doi.org/10.3390/biomedicines10123116
Chicago/Turabian StyleArmeli, Federica, Beatrice Mengoni, Elisa Maggi, Cristina Mazzoni, Adele Preziosi, Patrizia Mancini, Rita Businaro, Thomas Lenz, and Trevor Archer. 2022. "Milmed Yeast Alters the LPS-Induced M1 Microglia Cells to Form M2 Anti-Inflammatory Phenotype" Biomedicines 10, no. 12: 3116. https://doi.org/10.3390/biomedicines10123116