
Oncogenic Roles of Polycomb Repressive Complex 2 in Bladder Cancer and Upper Tract Urothelial Carcinoma
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Analysis of Public-Domain Lower Trcat Urinary Cancer Dataset
2.2. Patients and Samples of Upper Tract Urinary Cancers
2.3. Immunohistochemistry Staining of Major Subunits of PRC2 and the Methylated Substrate H3K27me3
2.4. Transcriptomic Profiling by RNA Sequencing
2.5. Statistical Analysis
3. Results
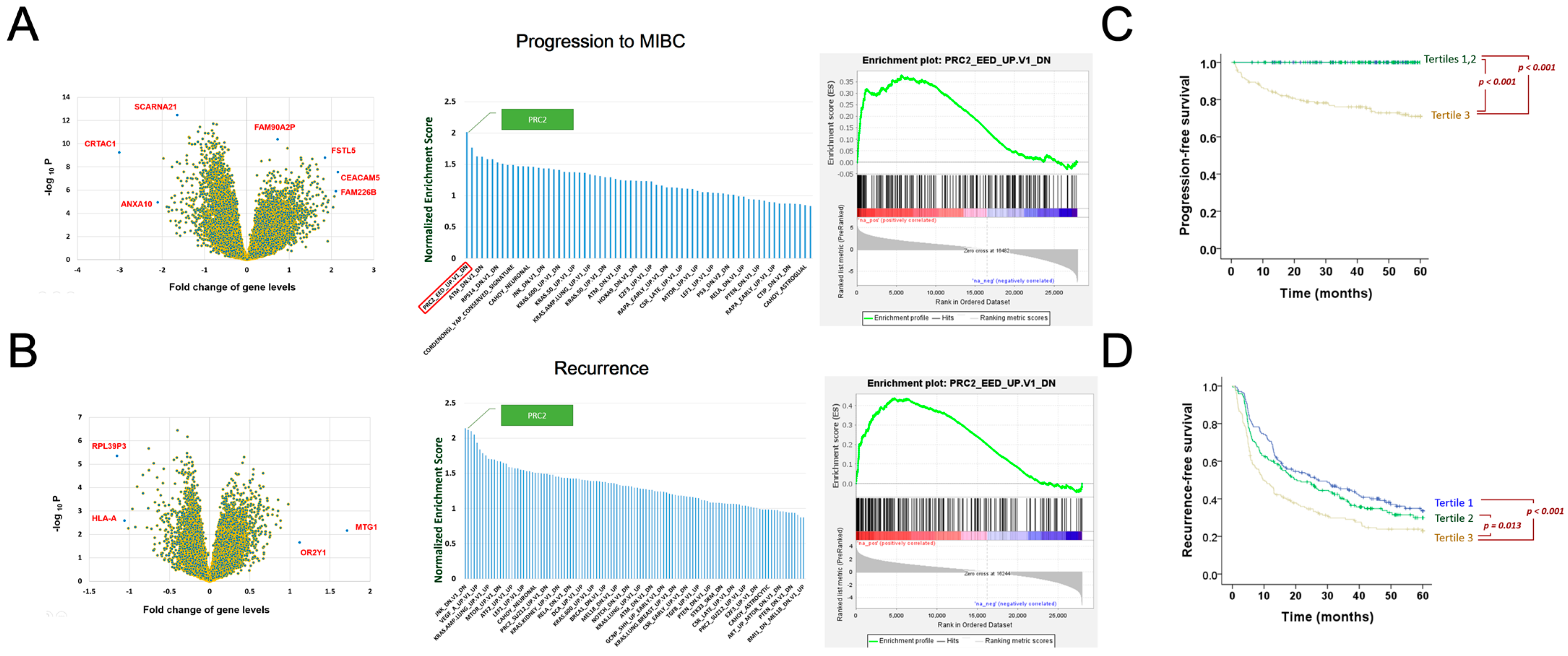
3.1. Polycomb Repressive Complex 2 Identified as a Major Driving Mechanism of Bladder Cancer Recurrence and Progression
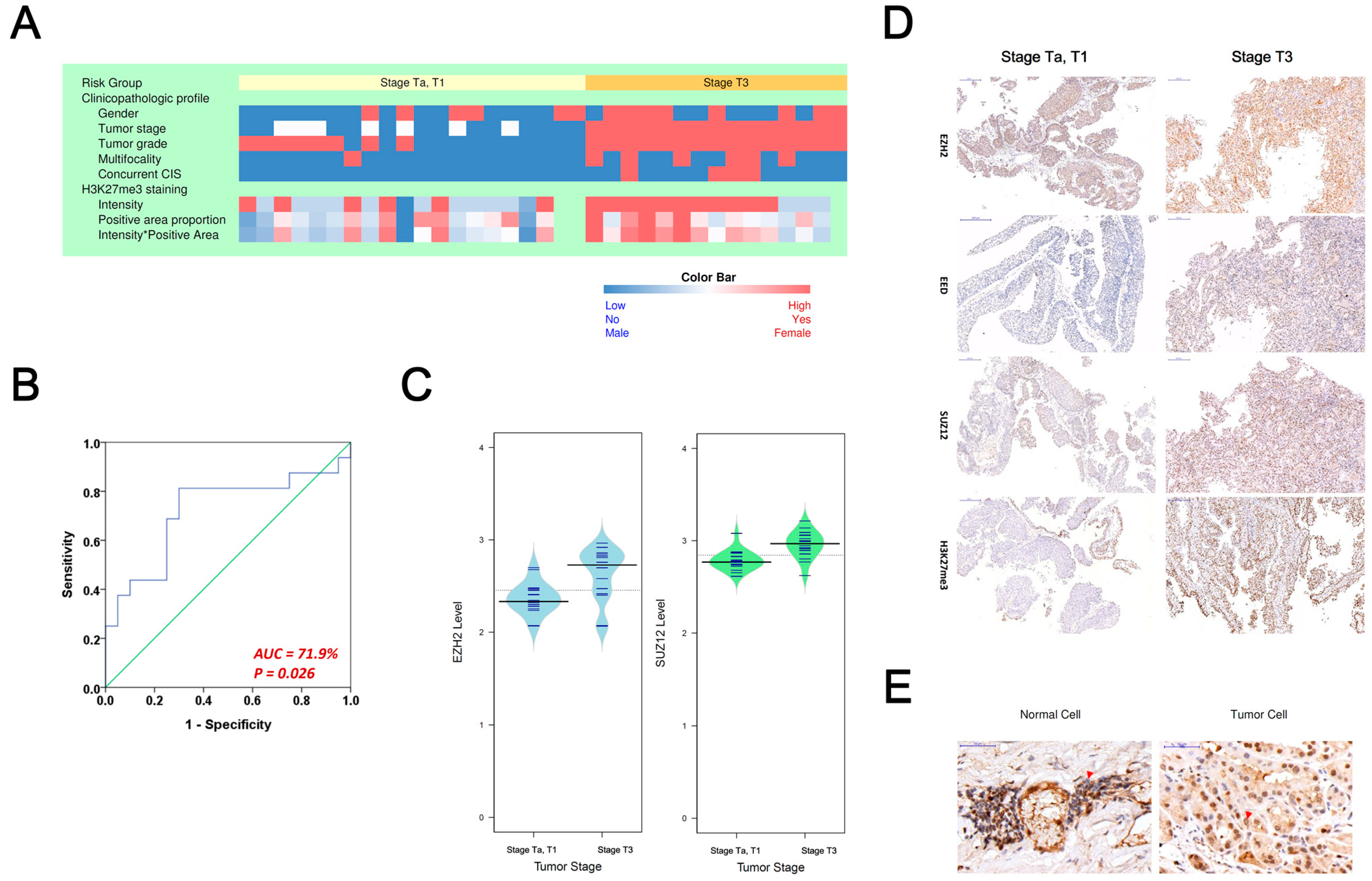
3.2. Evaluating PRC2 Oncogenic Signature in Asian Upper-Tract Urothelial Carcinoma Patients
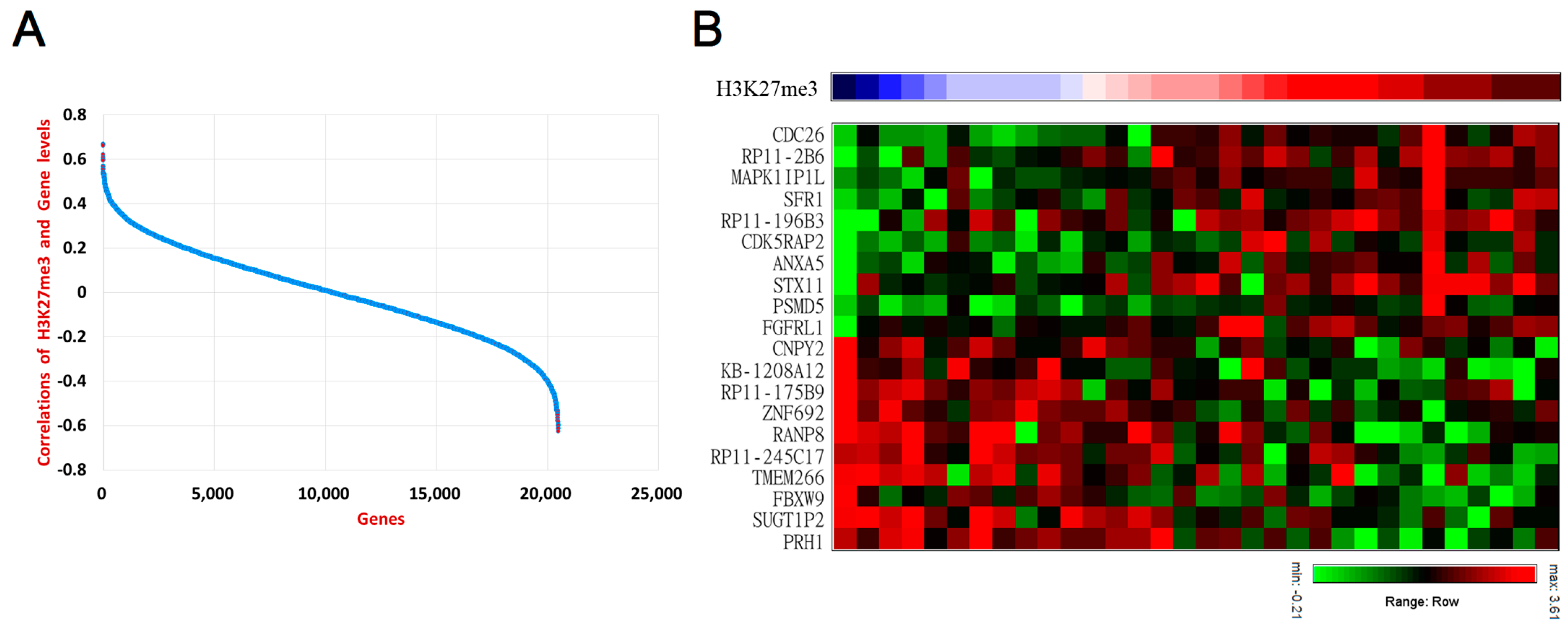
3.3. Deciphering Downsrtream Oncogenic Mechanism of PRC2 in UTUC Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hickling, D.R.; Sun, T.-T.; Wu, X.-R. Anatomy and Physiology of the Urinary Tract: Relation to Host Defense and Microbial Infection. Microbiol. Spectr. 2015, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jazayeri, S.B.; Liu, J.S.; Weissman, B.; Lester, J.; Samadi, D.B.; Feuerstein, M.A. Comparison of Adjuvant Chemotherapy for Upper Tract versus Lower Tract Urothelial Carcinoma: A Systematic Review and Meta-Analysis. Curr. Urol. 2018, 12, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Al-Husseini, M.J.; Kunbaz, A.; Saad, A.M.; Santos, J.V.; Salahia, S.; Iqbal, M.; Alahdab, F. Trends in the incidence and mortality of transitional cell carcinoma of the bladder for the last four decades in the USA: A SEER-based analysis. BMC Cancer 2019, 19, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Doeveren, T.; van de Werken, H.J.G.; van Riet, J.; Aben, K.K.H.; van Leeuwen, P.J.; Zwarthoff, E.C.; Boormans, J.L. Synchronous and metachronous urothelial carcinoma of the upper urinary tract and the bladder: Are they clonally related? A systematic review. Urol. Oncol. 2020, 38, 590–598. [Google Scholar] [CrossRef] [PubMed]
- Green, D.A.; Rink, M.; Xylinas, E.; Matin, S.F.; Stenzl, A.; Roupret, M.; Karakiewicz, P.I.; Scherr, D.S.; Shariat, S.F. Urothelial carcinoma of the bladder and the upper tract: Disparate twins. J. Urol. 2013, 189, 1214–1221. [Google Scholar] [CrossRef] [PubMed]
- Leow, J.J.; Chong, K.T.; Chang, S.L.; Bellmunt, J. Upper tract urothelial carcinoma: A different disease entity in terms of management. ESMO Open 2016, 1, e000126. [Google Scholar] [CrossRef]
- Van Doeveren, T.; van der Mark, M.; van Leeuwen, P.J.; Boormans, J.L.; Aben, K.K.H. Rising incidence rates and unaltered survival rates for primary upper urinary tract urothelial carcinoma: A Dutch population-based study from 1993 to 2017. BJU Int. 2021, 128, 343–351. [Google Scholar] [CrossRef]
- Montella, M.; Di Maso, M.; Crispo, A.; Grimaldi, M.; Bosetti, C.; Turati, F.; Giudice, A.; Libra, M.; Serraino, D.; La Vecchia, C.; et al. Metabolic syndrome and the risk of urothelial carcinoma of the bladder: A case-control study. BMC Cancer 2015, 15, 720. [Google Scholar] [CrossRef] [Green Version]
- Andreassen, B.K.; Aagnes, B.; Gislefoss, R.; Andreassen, M.; Wahlqvist, R. Incidence and Survival of urothelial carcinoma of the urinary bladder in Norway 1981–2014. BMC Cancer 2016, 16, 799. [Google Scholar] [CrossRef] [Green Version]
- Shariat, S.F.; Sfakianos, J.P.; Droller, M.J.; Karakiewicz, P.I.; Meryn, S.; Bochner, B.H. The effect of age and gender on bladder cancer: A critical review of the literature. BJU Int. 2010, 105, 300–308. [Google Scholar] [CrossRef]
- Shen, C.H.; Chiou, H.Y.; Tung, M.C.; Wu, C.C.; Kao, W.T.; Wang, Y.H.; Juang, G.D. Clinical and demographic characteristics among patients with urothelial carcinomas of the upper urinary tract and bladder in Taiwan. J. Chin. Med. Assoc. 2017, 80, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.H.; Chen, K.K.; Yen, C.C.; Wang, W.S.; Chang, Y.H.; Huang, W.J.; Fan, F.S.; Chiou, T.J.; Liu, J.H.; Chen, P.M. Unusually high incidence of upper urinary tract urothelial carcinoma in Taiwan. Urology 2002, 59, 681–687. [Google Scholar] [CrossRef]
- Chiou, H.Y.; Chiou, S.T.; Hsu, Y.H.; Chou, Y.L.; Tseng, C.H.; Wei, M.L.; Chen, C.J. Incidence of transitional cell carcinoma and arsenic in drinking water: A follow-up study of 8,102 residents in an arseniasis-endemic area in northeastern Taiwan. Am. J. Epidemiol. 2001, 153, 411–418. [Google Scholar] [CrossRef]
- Wang, S.M.; Lai, M.N.; Chen, P.C.; Pu, Y.S.; Lai, M.K.; Hwang, J.S.; Wang, J.D. Increased upper and lower tract urothelial carcinoma in patients with end-stage renal disease: A nationwide cohort study in Taiwan during 1997–2008. BioMed Res. Int. 2014, 2014, 149750. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.T.; Luo, H.L.; Wang, H.J.; Chen, Y.T.; Cheng, Y.T.; Chiang, P.H. Gender effect on the oncologic outcomes of upper urinary tract urothelial carcinoma in Taiwan. Int. Urol. Nephrol. 2020, 52, 1043–1048. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Bagrodia, A.; Cha, E.K.; Coleman, J.A. Prognostic Genetic Signatures in Upper Tract Urothelial Carcinoma. Curr. Urol. Rep. 2016, 17, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singla, N.; Margulis, V. Differences between Upper Tract Urothelial Carcinoma and Bladder Cancer. AUA News 2021, 26, 15–16. [Google Scholar]
- Mitra, A.P. Molecular substratification of bladder cancer: Moving towards individualized patient management. Ther. Adv. Urol. 2016, 8, 215–233. [Google Scholar] [CrossRef] [Green Version]
- Fujii, Y.; Sato, Y.; Suzuki, H.; Kakiuchi, N.; Yoshizato, T.; Lenis, A.T.; Maekawa, S.; Yokoyama, A.; Takeuchi, Y.; Inoue, Y.; et al. Molecular classification and diagnostics of upper urinary tract urothelial carcinoma. Cancer Cell 2021, 39, 793–809.e798. [Google Scholar] [CrossRef]
- Esteller, M. Cancer epigenomics: DNA methylomes and histone-modification maps. Nat. Rev. Genet. 2007, 8, 286–298. [Google Scholar] [CrossRef]
- Hoffmann, M.J.; Schulz, W.A. Alterations of Chromatin Regulators in the Pathogenesis of Urinary Bladder Urothelial Carcinoma. Cancers 2021, 13, 6040. [Google Scholar] [CrossRef] [PubMed]
- Schulz, W.A.; Koutsogiannouli, E.A.; Niegisch, G.; Hoffmann, M.J. Epigenetics of urothelial carcinoma. Methods Mol. Biol. 2015, 1238, 183–215. [Google Scholar] [CrossRef] [PubMed]
- Colin, P.; Koenig, P.; Ouzzane, A.; Berthon, N.; Villers, A.; Biserte, J.; Rouprêt, M. Environmental factors involved in carcinogenesis of urothelial cell carcinomas of the upper urinary tract. BJU Int. 2009, 104, 1436–1440. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm-Benartzi, C.S.; Christensen, B.C.; Koestler, D.C.; Houseman, E.A.; Schned, A.R.; Karagas, M.R.; Kelsey, K.T.; Marsit, C.J. Association of secondhand smoke exposures with DNA methylation in bladder carcinomas. Cancer Causes Control 2011, 22, 1205–1213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, T.-H.; Wu, C.-H.; Yeh, C.-T.; Su, S.-C.; Hsia, S.-M.; Liang, K.-H.; Chen, C.-C.; Hsueh, C.; Chen, C.-Y. Melatonin suppresses hepatocellular carcinoma progression via lncRNA-CPS1-IT-mediated HIF-1α inactivation. Oncotarget 2017, 8, 82280–82293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stiborova, M.; Frei, E.; Arlt, V.M.; Schmeiser, H.H. The role of biotransformation enzymes in the development of renal injury and urothelial cancer caused by aristolochic acid: Urgent questions and difficult answers. Biomed. Pap. Med. Fac. Palacky Univ. Olomouc 2009, 153, 5–11. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.H.; Dickman, K.G.; Moriya, M.; Zavadil, J.; Sidorenko, V.S.; Edwards, K.L.; Gnatenko, D.V.; Wu, L.; Turesky, R.J.; Wu, X.R.; et al. Aristolochic acid-associated urothelial cancer in Taiwan. Proc. Natl. Acad. Sci. USA 2012, 109, 8241–8246. [Google Scholar] [CrossRef] [Green Version]
- Lindskrog, S.V.; Prip, F.; Lamy, P.; Taber, A.; Groeneveld, C.S.; Birkenkamp-Demtröder, K.; Jensen, J.B.; Strandgaard, T.; Nordentoft, I.; Christensen, E.; et al. An integrated multi-omics analysis identifies prognostic molecular subtypes of non-muscle-invasive bladder cancer. Nat. Commun. 2021, 12, 2301. [Google Scholar] [CrossRef]
- Wang, T.-H.; Chen, C.-C.; Liang, K.-H.; Chen, C.-Y.; Chuang, W.-Y.; Ueng, S.-H.; Chu, P.-H.; Huang, C.-G.; Chen, T.-C.; Hsueh, C. A Multivariate Evaluation of Factors Affecting the Quality of Freshly Frozen Tissue Specimens. Biopreservation Biobanking 2017, 15, 344–349. [Google Scholar] [CrossRef]
- Liang, K.-H.; Cheng, M.-L.; Lo, C.-J.; Lin, Y.-H.; Lai, M.-W.; Lin, W.-R.; Yeh, C.-T. Plasma phenylalanine and glutamine concentrations correlate with subsequent hepatocellular carcinoma occurrence in liver cirrhosis patients: An exploratory study. Sci. Rep. 2020, 10, 10926. [Google Scholar] [CrossRef]
- Liang, K.-H.; Lai, M.-W.; Lin, Y.-H.; Chu, Y.-D.; Lin, C.-L.; Lin, W.-R.; Huang, Y.-H.; Wang, T.-H.; Chien, R.-N.; Hu, T.-H.; et al. Plasma interleukin-17 and alpha-fetoprotein combination effectively predicts imminent hepatocellular carcinoma occurrence in liver cirrhotic patients. BMC Gastroenterol. 2021, 21, 177. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha Vamsi, K.; Mukherjee, S.; Ebert Benjamin, L.; Gillette Michael, A.; Paulovich, A.; Pomeroy Scott, L.; Golub Todd, R.; Lander Eric, S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barski, A.; Cuddapah, S.; Cui, K.; Roh, T.Y.; Schones, D.E.; Wang, Z.; Wei, G.; Chepelev, I.; Zhao, K. High-resolution profiling of histone methylations in the human genome. Cell 2007, 129, 823–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, Z.; Ye, Z.; Xue, Z.; Wu, L.; Ouyang, Y.; Yao, C.; Cui, C.; Xu, N.; Ma, J.; Hou, G.; et al. Identification of Renal Long Non-coding RNA RP11-2B6.2 as a Positive Regulator of Type I Interferon Signaling Pathway in Lupus Nephritis. Front. Immunol. 2019, 10, 975. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hanahan, D. Hallmarks of Cancer: New Dimensions. Cancer Discov. 2022, 12, 31–46. [Google Scholar] [CrossRef]
- Kouzarides, T. Chromatin modifications and their function. Cell 2007, 128, 693–705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bannister, A.J.; Kouzarides, T. Regulation of chromatin by histone modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Plath, K.; Fang, J.; Mlynarczyk-Evans, S.K.; Cao, R.; Worringer, K.A.; Wang, H.; de la Cruz, C.C.; Otte, A.P.; Panning, B.; Zhang, Y. Role of histone H3 lysine 27 methylation in X inactivation. Science 2003, 300, 131–135. [Google Scholar] [CrossRef] [Green Version]
- Heard, E. Delving into the diversity of facultative heterochromatin: The epigenetics of the inactive X chromosome. Curr. Opin. Genet. Dev. 2005, 15, 482–489. [Google Scholar] [CrossRef]
- Portela, A.; Esteller, M. Epigenetic modifications and human disease. Nat. Biotechnol. 2010, 28, 1057–1068. [Google Scholar] [CrossRef]
- Park, P.J. ChIP–seq: Advantages and challenges of a maturing technology. Nat. Rev. Genet. 2009, 10, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Albert, M.; Helin, K. Histone methyltransferases in cancer. Semin. Cell Amp Dev. Biol. 2010, 21, 209–220. [Google Scholar] [CrossRef] [PubMed]
- Margueron, R.; Reinberg, D. The Polycomb complex PRC2 and its mark in life. Nature 2011, 469, 343–349. [Google Scholar] [CrossRef] [Green Version]
- Ren, G.; Baritaki, S.; Marathe, H.; Feng, J.; Park, S.; Beach, S.; Bazeley, P.S.; Beshir, A.B.; Fenteany, G.; Mehra, R.; et al. Polycomb Protein EZH2 Regulates Tumor Invasion via the Transcriptional Repression of the Metastasis Suppressor RKIP in Breast and Prostate Cancer. Cancer Res. 2012, 72, 3091–3104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comet, I.; Riising, E.M.; Leblanc, B.; Helin, K. Maintaining cell identity: PRC2-mediated regulation of transcription and cancer. Nat. Rev. Cancer 2016, 16, 803–810. [Google Scholar] [CrossRef]
- Laugesen, A.; Højfeldt, J.W.; Helin, K. Role of the Polycomb Repressive Complex 2 (PRC2) in Transcriptional Regulation and Cancer. Cold Spring Harb. Perspect. Med. 2016, 6, a026575. [Google Scholar] [CrossRef] [Green Version]
- Pekmezci, M.; Cuevas-Ocampo, A.K.; Perry, A.; Horvai, A.E. Significance of H3K27me3 loss in the diagnosis of malignant peripheral nerve sheath tumors. Mod. Pathol. 2017, 30, 1710–1719. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.J.; Kim, S.H.; Kim, E.K.; Han, J.W.; Shin, K.-H.; Hu, H.; Kim, K.S.; Choi, Y.D.; Kim, S.; Lee, Y.H.; et al. Prognostic implications of polycomb proteins ezh2, suz12, and eed1 and histone modification by H3K27me3 in sarcoma. BMC Cancer 2018, 18, 158. [Google Scholar] [CrossRef] [Green Version]
- Dong, P.; Xiong, Y.; Yue, J.; Hanley, S.J.B.; Kobayashi, N.; Todo, Y.; Watari, H. Long Non-coding RNA NEAT1: A Novel Target for Diagnosis and Therapy in Human Tumors. Front. Genet. 2018, 9, 471. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, F.; Niebel, D.; Aymans, P.; Ferring-Schmitt, S.; Dietrich, D.; Landsberg, J. H3K27me3 and EZH2 expression in melanoma: Relevance for melanoma progression and response to immune checkpoint blockade. Clin. Epigenetics 2020, 12, 24. [Google Scholar] [CrossRef] [Green Version]
- Massa, A.T.; Mousel, M.R.; Herndon, M.K.; Herndon, D.R.; Murdoch, B.M.; White, S.N. Genome-Wide Histone Modifications and CTCF Enrichment Predict Gene Expression in Sheep Macrophages. Front. Genet. 2021, 11, 612031. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.-H.; Song, C.G. Epigenetic regulation in bladder cancer: Development of new prognostic targets and therapeutic implications. Transl. Cancer Res. 2017, 6, S677–S688. [Google Scholar] [CrossRef]
- Varambally, S.; Dhanasekaran, S.M.; Zhou, M.; Barrette, T.R.; Kumar-Sinha, C.; Sanda, M.G.; Ghosh, D.; Pienta, K.J.; Sewalt, R.G.; Otte, A.P.; et al. The polycomb group protein EZH2 is involved in progression of prostate cancer. Nature 2002, 419, 624–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, C.J.; Yang, J.Y.; Xia, W.; Chen, C.T.; Xie, X.; Chao, C.H.; Woodward, W.A.; Hsu, J.M.; Hortobagyi, G.N.; Hung, M.C. EZH2 promotes expansion of breast tumor initiating cells through activation of RAF1-β-catenin signaling. Cancer Cell 2011, 19, 86–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amatangelo, M.D.; Garipov, A.; Li, H.; Conejo-Garcia, J.R.; Speicher, D.W.; Zhang, R. Three-dimensional culture sensitizes epithelial ovarian cancer cells to EZH2 methyltransferase inhibition. Cell Cycle 2013, 12, 2113–2119. [Google Scholar] [CrossRef] [Green Version]
- Takawa, M.; Masuda, K.; Kunizaki, M.; Daigo, Y.; Takagi, K.; Iwai, Y.; Cho, H.S.; Toyokawa, G.; Yamane, Y.; Maejima, K.; et al. Validation of the histone methyltransferase EZH2 as a therapeutic target for various types of human cancer and as a prognostic marker. Cancer Sci. 2011, 102, 1298–1305. [Google Scholar] [CrossRef]
- Ernst, T.; Chase, A.J.; Score, J.; Hidalgo-Curtis, C.E.; Bryant, C.; Jones, A.V.; Waghorn, K.; Zoi, K.; Ross, F.M.; Reiter, A.; et al. Inactivating mutations of the histone methyltransferase gene EZH2 in myeloid disorders. Nat. Genet. 2010, 42, 722–726. [Google Scholar] [CrossRef]
- Nikoloski, G.; Langemeijer, S.M.; Kuiper, R.P.; Knops, R.; Massop, M.; Tönnissen, E.R.; van der Heijden, A.; Scheele, T.N.; Vandenberghe, P.; de Witte, T.; et al. Somatic mutations of the histone methyltransferase gene EZH2 in myelodysplastic syndromes. Nat. Genet. 2010, 42, 665–667. [Google Scholar] [CrossRef]
- Kleer, C.G.; Cao, Q.; Varambally, S.; Shen, R.; Ota, I.; Tomlins, S.A.; Ghosh, D.; Sewalt, R.G.; Otte, A.P.; Hayes, D.F.; et al. EZH2 is a marker of aggressive breast cancer and promotes neoplastic transformation of breast epithelial cells. Proc. Natl. Acad. Sci. USA 2003, 100, 11606–11611. [Google Scholar] [CrossRef] [Green Version]
- Davidovich, C.; Cech, T.R. The recruitment of chromatin modifiers by long noncoding RNAs: Lessons from PRC2. RNA 2015, 21, 2007–2022. [Google Scholar] [CrossRef] [Green Version]
- Taheri, M.; Omrani, M.D.; Ghafouri-Fard, S. Long non-coding RNA expression in bladder cancer. Biophys. Rev. 2018, 10, 1205–1213. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.R.; Roh, Y.G.; Kim, S.K.; Lee, J.S.; Seol, S.Y.; Lee, H.H.; Kim, W.T.; Kim, W.J.; Heo, J.; Cha, H.J.; et al. Activation of EZH2 and SUZ12 Regulated by E2F1 Predicts the Disease Progression and Aggressive Characteristics of Bladder Cancer. Clin. Cancer Res. 2015, 21, 5391–5403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abudurexiti, M.; Xie, H.; Jia, Z.; Zhu, Y.; Zhu, Y.; Shi, G.; Zhang, H.; Dai, B.; Wan, F.; Shen, Y.; et al. Development and External Validation of a Novel 12-Gene Signature for Prediction of Overall Survival in Muscle-Invasive Bladder Cancer. Front. Oncol. 2019, 9, 856. [Google Scholar] [CrossRef] [PubMed]
- Weikert, S.; Christoph, F.; Köllermann, J.; Müller, M.; Schrader, M.; Miller, K.; Krause, H. Expression levels of the EZH2 polycomb transcriptional repressor correlate with aggressiveness and invasive potential of bladder carcinomas. Int. J. Mol. Med. 2005, 16, 349–353. [Google Scholar] [CrossRef]
- Martínez-Fernández, M.; Rubio, C.; Segovia, C.; López-Calderón, F.F.; Dueñas, M.; Paramio, J.M. EZH2 in Bladder Cancer, a Promising Therapeutic Target. Int. J. Mol. Sci. 2015, 16, 27107–27132. [Google Scholar] [CrossRef] [Green Version]
- Crocetto, F.; Barone, B.; Ferro, M.; Busetto, G.M.; La Civita, E.; Buonerba, C.; Di Lorenzo, G.; Terracciano, D.; Schalken, J.A. Liquid biopsy in bladder cancer: State of the art and future perspectives. Crit. Rev. Oncol. Hematol. 2022, 170, 103577. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-T.; Chen, H.-W.; Domanski, D.; Smith, D.S.; Liang, K.-H.; Wu, C.-C.; Chen, C.-L.; Chung, T.; Chen, M.-C.; Chang, Y.-S.; et al. Multiplexed quantification of 63 proteins in human urine by multiple reaction monitoring-based mass spectrometry for discovery of potential bladder cancer biomarkers. J. Proteom. 2012, 75, 3529–3545. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Tumor Stage Ta/T1 | Tumor Stage T3 | p Value | |
|---|---|---|---|
| Number | 20 | 16 | |
| Gender | 0.190 | ||
| Male | 14 (70%) | 8 (50%) | |
| Female | 6 (30%) | 8 (50%) | |
| Age | 62.25 ± 13.93 | 62.56 ± 8.18 | 0.934 |
| Hydronephrosis | 12 (60%) | 13 (81%) | 0.156 |
| Smoking | 8 (40%) | 2 (13%) | 0.071 |
| Hypertension | 9 (45%) | 4 (25%) | 0.187 |
| Diabetes | 7 (35%) | 2 (7.0%) | 0.122 |
| Coronary artery disease | 3 (15%) | 0 (0.0%) | 0.160 |
| ECOG | 0.577 | ||
| 0 | 8 (40%) | 6 (37%) | |
| 1 | 12 (60%) | 10 (63%) | |
| Location | 0.090 | ||
| Renal pelvis | 10 (53%) | 11 (69%) | |
| Ureter | 9 (47%) | 3 (19%) | |
| Renal pelvis + ureter | 0 (0.0%) | 2 (13%) | |
| Morphology | 0.444 | ||
| Papillary | 20 (100.0%) | 15 (94%) | |
| Non-papillary | 0 (0.0%) | 1 (6%) | |
| Pathological T stage | <0.001 | ||
| Ta | 13 (65.0%) | ||
| T1 | 7 (35.0%) | ||
| T3 | 16 (100%) | ||
| Tumor grade | <0.001 | ||
| Low grade | 12 (60%) | 0 (0%) | |
| High grade | 8 (40%) | 16 (100%) | |
| Tumor size | 0.007 | ||
| <3 cm | 11 (55.0%) | 2 (13%) | |
| ≧3 cm | 8 (40.0%) | 14 (88%) | |
| Multifocal | 1 (5.0%) | 6 (37.5%) | 0.021 |
| Concurrent CIS | 0 (0.0%) | 4 (25%) | 0.031 |
| Lymphovascular invasion | 0 (0.0%) | 2 (13%) | 0.190 |
| H3K27me3 staining | |||
| Intensity | 2.17 ± 0.79 | 2.79 ± 0.43 | 0.006 |
| Positive area proportion | 0.63 ± 0.27 | 0.78 ± 0.13 | 0.094 |
| Intensity × Area | 1.50 ± 0.83 | 2.20 ± 0.57 | 0.015 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, E.Y.-H.; Chen, Y.-K.; Ou, C.-P.; Chen, Y.-T.; Chen, S.-F.; Huang, W.J.; Liang, K.-H. Oncogenic Roles of Polycomb Repressive Complex 2 in Bladder Cancer and Upper Tract Urothelial Carcinoma. Biomedicines 2022, 10, 2925. https://doi.org/10.3390/biomedicines10112925
Huang EY-H, Chen Y-K, Ou C-P, Chen Y-T, Chen S-F, Huang WJ, Liang K-H. Oncogenic Roles of Polycomb Repressive Complex 2 in Bladder Cancer and Upper Tract Urothelial Carcinoma. Biomedicines. 2022; 10(11):2925. https://doi.org/10.3390/biomedicines10112925
Chicago/Turabian StyleHuang, Eric Yi-Hsiu, Yu-Kuang Chen, Chen-Pu Ou, Yi-Ting Chen, Sung-Fang Chen, William J. Huang, and Kung-Hao Liang. 2022. "Oncogenic Roles of Polycomb Repressive Complex 2 in Bladder Cancer and Upper Tract Urothelial Carcinoma" Biomedicines 10, no. 11: 2925. https://doi.org/10.3390/biomedicines10112925