
Chondrogenic Potential of Human Umbilical Cord Mesenchymal Stem Cells Cultured with Exosome-Depleted Fetal Bovine Serum in an Osteoarthritis Mouse Model

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Derivation and Culture of Human Umbilical Mesenchymal Stem Cells
2.3. Exosome-Depleted FBS (Exo(-)FBS)
2.4. XTT Assay
2.5. Flow Cytometry
2.6. Annexin V/PI Staining
2.7. Induction to Adipocytes
2.8. Induction to Osteoblast
2.9. Induction to Chondrocyte
2.10. Quantitative RT-PCR Analyses
2.11. Type VII Collagenase-Induced Osteoarthritis in a Mouse Model
2.12. Transplantation of Human Umbilical Cord Mesenchymal Stem Cells
2.13. The Assessment of the Rotarod Behavior
2.14. Tissue Collection
2.15. Histological Evaluation
2.16. Immunohistochemistry
2.17. Statistical Analysis
3. Results
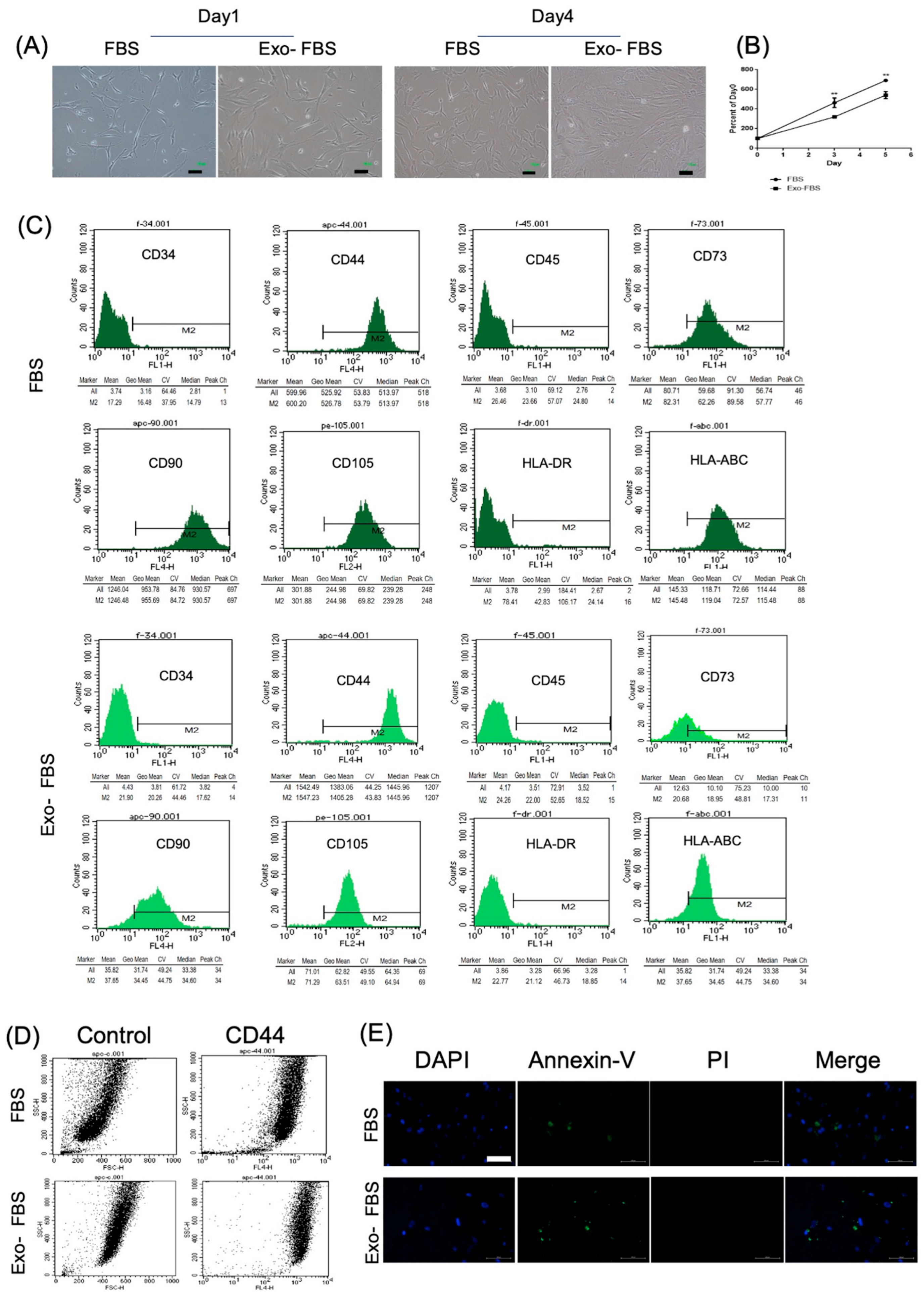
3.1. HUCMSCs Showed Characteristics of MSCs
3.1.1. Morphology, Proliferation, and Surface Marker Expression Levels
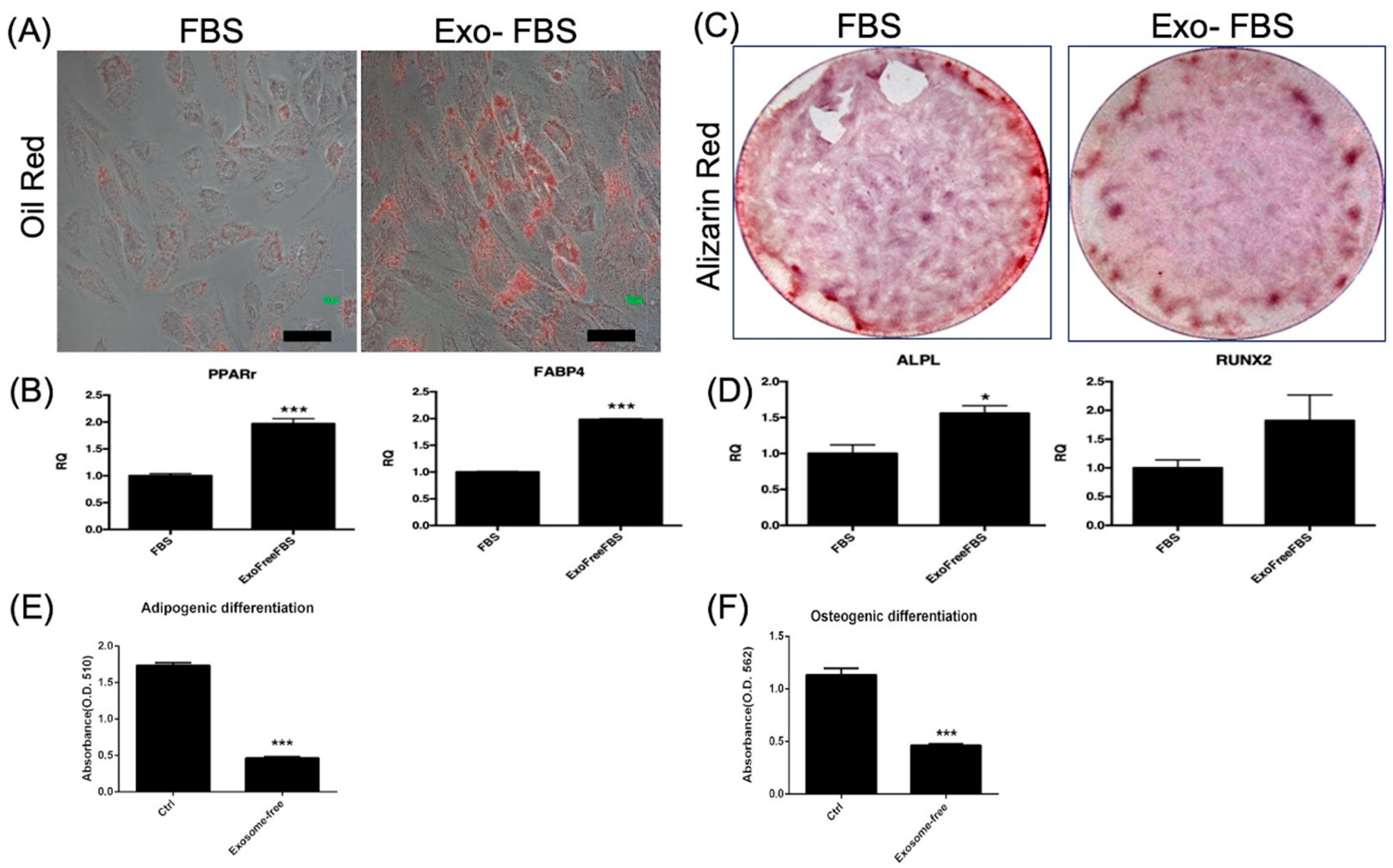
3.1.2. Adipocyte and Osteoblast Differentiation of HUCMSCs
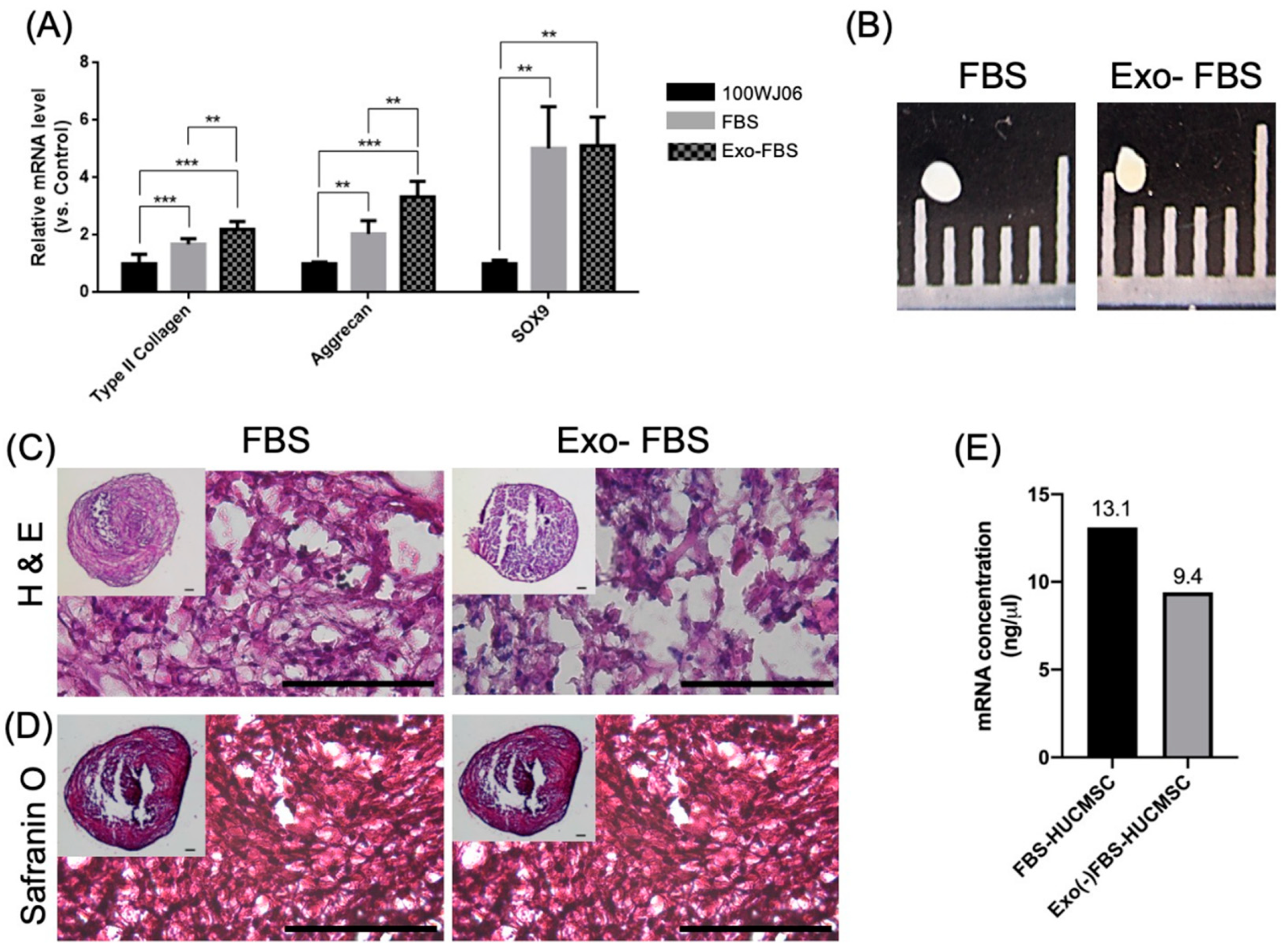
3.1.3. Chondrogenesis of FBS- and Exo(-)FBS-Cultured HUCMSCs
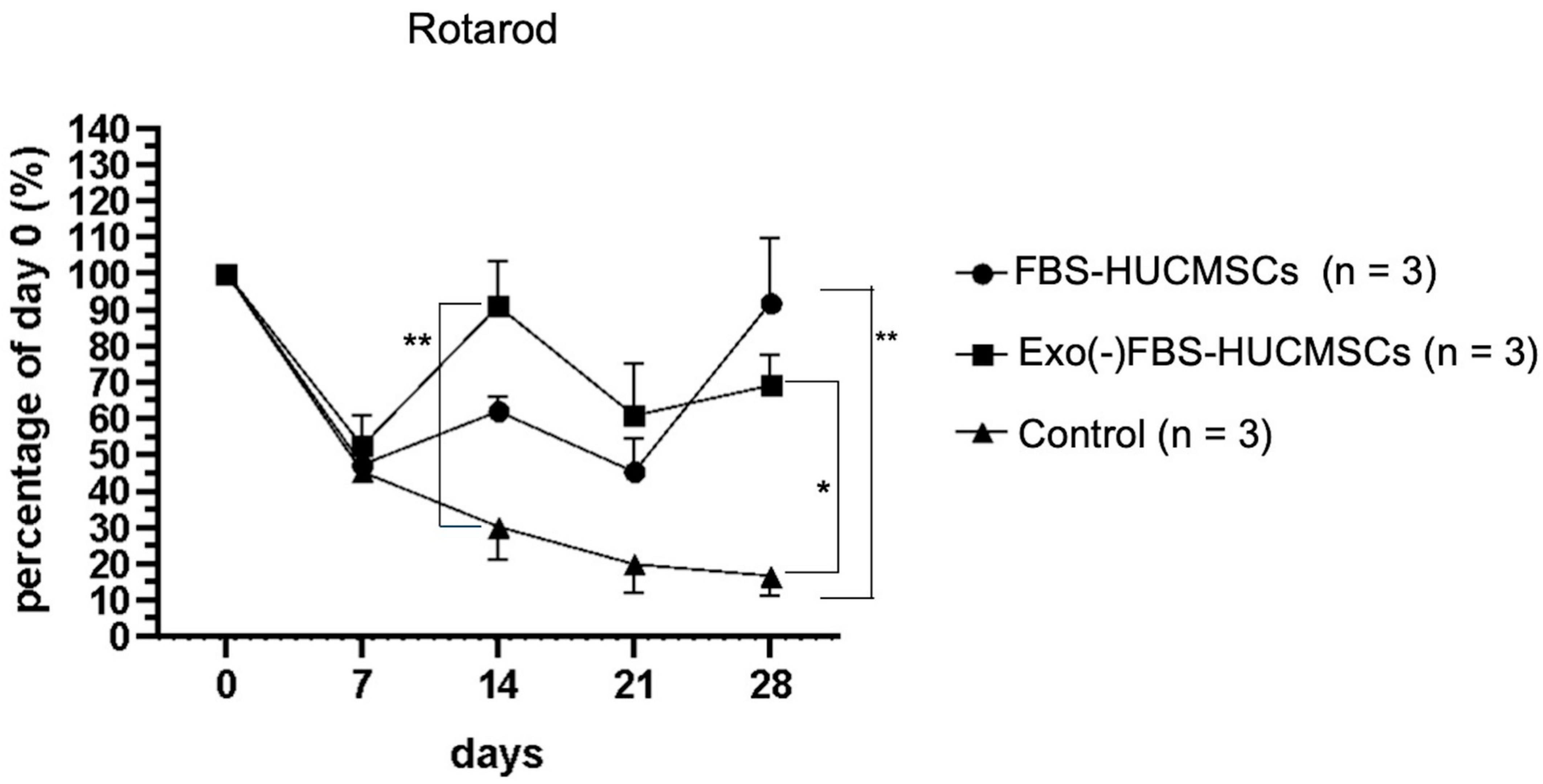
3.2. Rotarod Behavior after Treatment
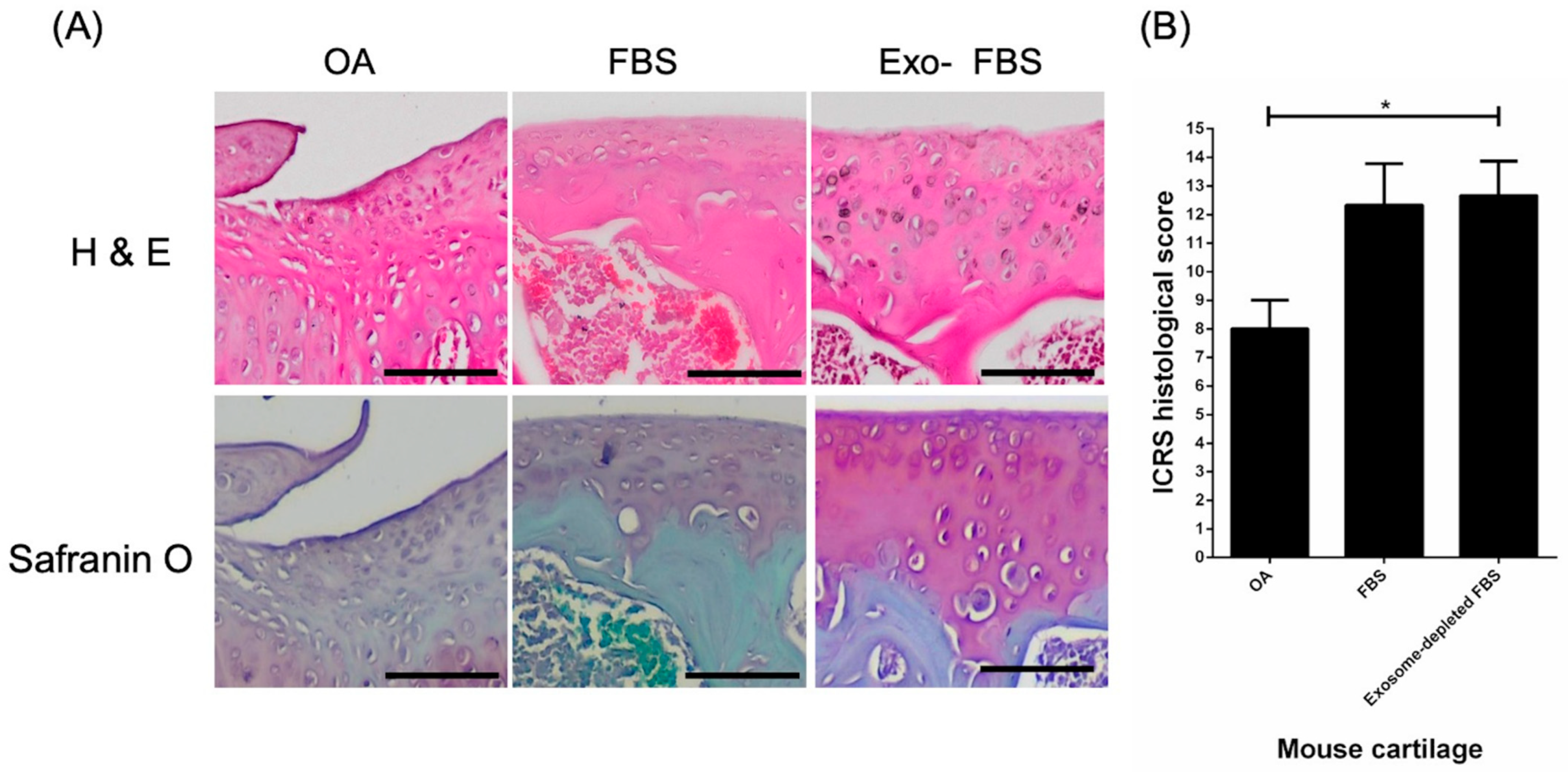
3.3. Histological Evidence of Cartilage Repair with HUCMSCs in Collagenase-Treated Mice
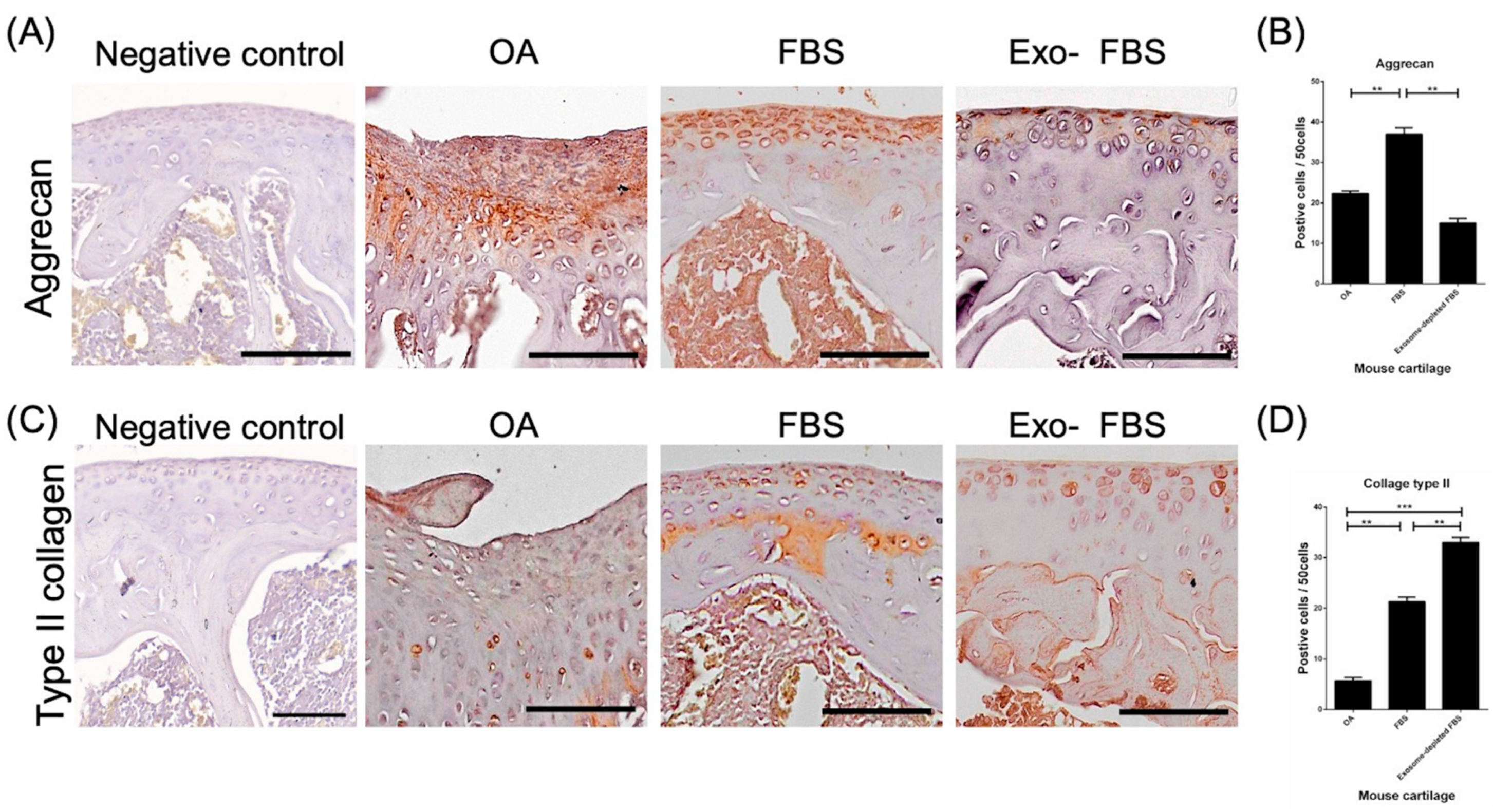
3.4. Increased Expression of Type II Collagen and Aggrecan after Transplantation of HUCMSCs
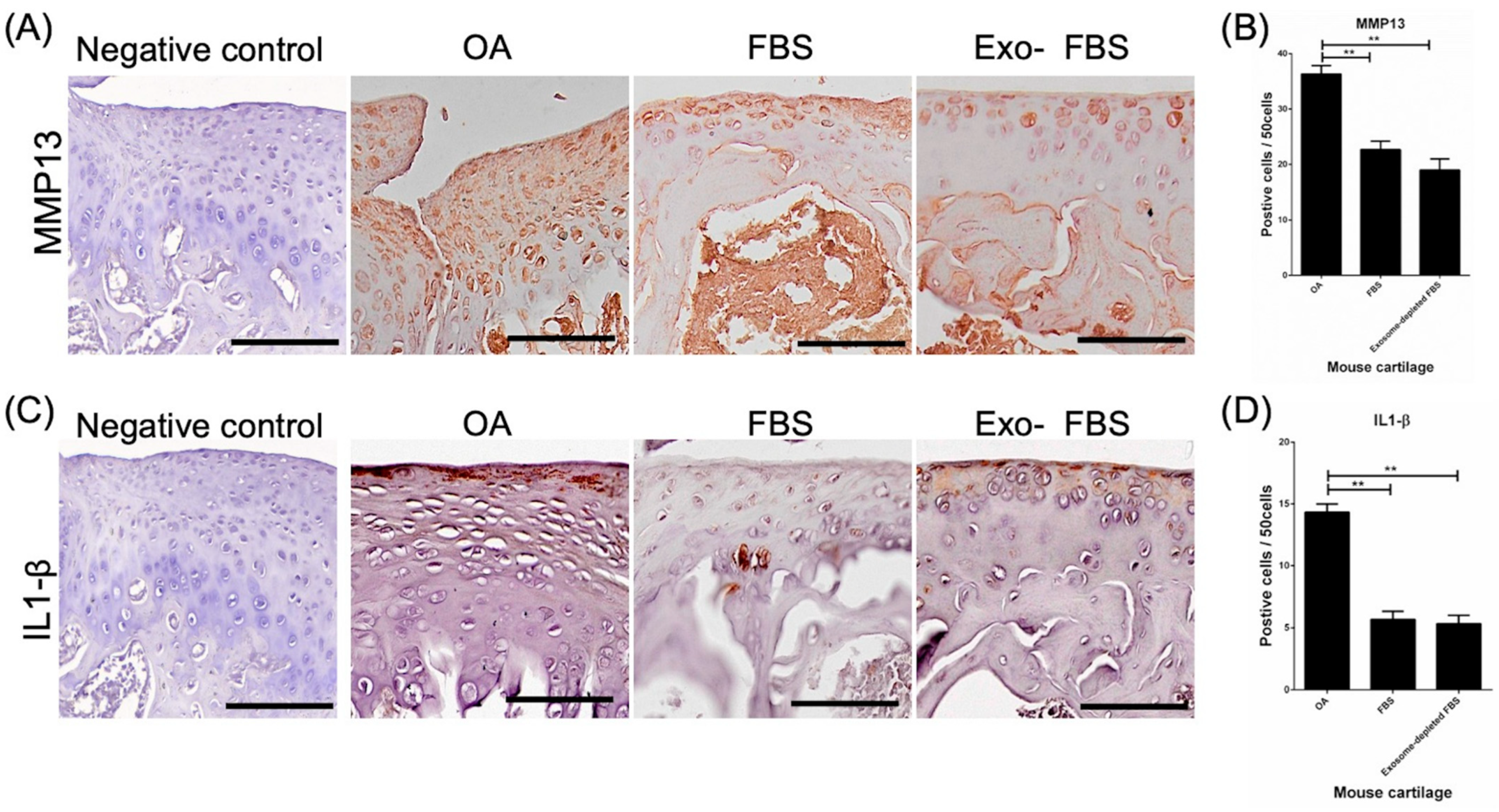
3.5. Decreased Expression of MMP13 and IL-1β in HUCMSC-Treated Cartilages
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kalamegam, G.; Memic, A.; Budd, E.; Abbas, M.; Mobasheri, A. A Comprehensive Review of Stem Cells for Cartilage Regeneration in Osteoarthritis. Adv. Exp. Med. Biol. 2018, 1089, 23–36. [Google Scholar]
- Wu, K.-C.; Chang, Y.-H.; Liu, H.-W.; Ding, D.-C. Transplanting Human Umbilical Cord Mesenchymal Stem Cells and Hyaluronate Hydrogel Repairs Cartilage of Osteoarthritis in the Minipig Model. Tzu Chi Med. J. 2018, 11, 11–19. [Google Scholar]
- Nosoudi, N.; Hart, C.; McKnight, I.; Esmaeilpour, M.; Ghomian, T.; Zadeh, A.; Raines, R.; Ramirez Vick, J.E. Differentiation of Adipose-Derived Stem Cells to Chondrocytes Using Electrospraying. Sci. Rep. 2021, 11, 24301. [Google Scholar] [CrossRef]
- Zhang, X.; Xue, K.; Zhou, J.; Xu, P.; Huang, H.; Liu, K. Chondrogenic Differentiation of Bone Marrow-derived Stem Cells Cultured in the Supernatant of Elastic Cartilage Cells. Mol. Med. Rep. 2015, 12, 5355–5360. [Google Scholar] [CrossRef] [Green Version]
- Najar, M.; Martel-Pelletier, J.; Pelletier, J.-P.; Fahmi, H. Mesenchymal Stromal Cell Immunology for Efficient and Safe Treatment of Osteoarthritis. Front. Cell Dev. Biol. 2020, 8, 567813. [Google Scholar] [CrossRef]
- Ding, D.-C.; Chang, Y.-H.; Shyu, W.-C.; Lin, S.-Z. Human Umbilical Cord Mesenchymal Stem Cells: A New Era for Stem Cell Therapy. Cell Transplant. 2015, 24, 339–347. [Google Scholar] [CrossRef]
- Angelini, F.; Ionta, V.; Rossi, F.; Miraldi, F.; Messina, E.; Giacomello, A. Foetal Bovine Serum-Derived Exosomes Affect Yield and Phenotype of Human Cardiac Progenitor Cell Culture. Bioimpacts 2016, 6, 15–24. [Google Scholar] [CrossRef]
- Ochieng, J.; Pratap, S.; Khatua, A.K.; Sakwe, A.M. Anchorage-Independent Growth of Breast Carcinoma Cells Is Mediated by Serum Exosomes. Exp. Cell Res. 2009, 315, 1875–1888. [Google Scholar] [CrossRef] [Green Version]
- Shelke, G.V.; Lässer, C.; Gho, Y.S.; Lötvall, J. Importance of Exosome Depletion Protocols to Eliminate Functional and RNA-Containing Extracellular Vesicles from Fetal Bovine Serum. J. Extracell. Vesicles 2014, 3, 24783. [Google Scholar] [CrossRef] [Green Version]
- Beninson, L.A.; Fleshner, M. Exosomes in Fetal Bovine Serum Dampen Primary Macrophage IL-1β Response to Lipopolysaccharide (LPS) Challenge. Immunol. Lett. 2015, 163, 187–192. [Google Scholar] [CrossRef]
- Théry, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and Characterization of Exosomes from Cell Culture Supernatants and Biological Fluids. Curr. Protoc. Cell Biol. 2006, 30, 3–22. [Google Scholar] [CrossRef]
- Aswad, H.; Jalabert, A.; Rome, S. Depleting Extracellular Vesicles from Fetal Bovine Serum Alters Proliferation and Differentiation of Skeletal Muscle Cells in Vitro. BMC Biotechnol. 2016, 16, 32. [Google Scholar] [CrossRef]
- Ding, D.-C.; Wen, Y.-T.; Tsai, R.-K. Pigment Epithelium-Derived Factor from ARPE19 Promotes Proliferation and Inhibits Apoptosis of Human Umbilical Mesenchymal Stem Cells in Serum-Free Medium. Exp. Mol. Med. 2017, 49, e411. [Google Scholar] [CrossRef]
- Chang, Y.-H.; Wu, K.-C.; Harn, H.-J.; Lin, S.-Z.; Ding, D.-C. Exosomes and Stem Cells in Degenerative Disease Diagnosis and Therapy. Cell Transplant. 2018, 27, 349–363. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Mi, T.; Jin, L.; Li, M.; Zhanghuang, C.; Wang, J.; Tan, X.; Lu, H.; Shen, L.; Long, C.; et al. Comprehensive Proteomic Analysis of Exosome Mimetic Vesicles and Exosomes Derived from Human Umbilical Cord Mesenchymal Stem Cells. Stem Cell Res. Ther. 2022, 13, 312. [Google Scholar] [CrossRef]
- Song, H.; Zhao, J.; Cheng, J.; Feng, Z.; Wang, J.; Momtazi-Borojeni, A.A.; Liang, Y. Extracellular Vesicles in Chondrogenesis and Cartilage Regeneration. J. Cell. Mol. Med. 2021, 25, 4883–4892. [Google Scholar] [CrossRef]
- Ding, D.-C.; Shyu, W.-C.; Chiang, M.-F.; Lin, S.-Z.; Chang, Y.-C.; Wang, H.-J.; Su, C.-Y.; Li, H. Enhancement of Neuroplasticity through Upregulation of beta1-Integrin in Human Umbilical Cord-Derived Stromal Cell Implanted Stroke Model. Neurobiol. Dis. 2007, 27, 339–353. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Rutgers, M.; van Pelt, M.J.P.; Dhert, W.J.A.; Creemers, L.B.; Saris, D.B.F. Evaluation of Histological Scoring Systems for Tissue-Engineered, Repaired and Osteoarthritic Cartilage. Osteoarthr. Cartil. 2010, 18, 12–23. [Google Scholar] [CrossRef] [Green Version]
- Jensen, E.C. Quantitative Analysis of Histological Staining and Fluorescence Using ImageJ: Histological Staining/Fluorescence Using ImageJ. Anat. Rec. 2013, 296, 378–381. [Google Scholar] [CrossRef]
- Thomas, M.P.; Liu, X.; Whangbo, J.; McCrossan, G.; Sanborn, K.B.; Basar, E.; Walch, M.; Lieberman, J. Apoptosis Triggers Specific, Rapid, and Global mRNA Decay with 3’ Uridylated Intermediates Degraded by DIS3L2. Cell Rep. 2015, 11, 1079–1089. [Google Scholar] [CrossRef] [Green Version]
- Mainil-Varlet, P.; Aigner, T.; Brittberg, M.; Bullough, P.; Hollander, A.; Hunziker, E.; Kandel, R.; Nehrer, S.; Pritzker, K.; Roberts, S.; et al. Histological Assessment of Cartilage Repair: A Report by the Histology Endpoint Committee of the International Cartilage Repair Society (ICRS). J. Bone Jt. Surg. Am. 2003, 85, 45–57. [Google Scholar] [CrossRef]
- Lehrich, B.M.; Liang, Y.; Fiandaca, M.S. Foetal Bovine Serum Influence on in Vitro Extracellular Vesicle Analyses. J. Extracell. Vesicles 2021, 10, e12061. [Google Scholar] [CrossRef]
- Eitan, E.; Zhang, S.; Witwer, K.W.; Mattson, M.P. Extracellular Vesicle-Depleted Fetal Bovine and Human Sera Have Reduced Capacity to Support Cell Growth. J. Extracell. Vesicles 2015, 4, 26373. [Google Scholar] [CrossRef]
- Lehrich, B.M.; Liang, Y.; Khosravi, P.; Federoff, H.J.; Fiandaca, M.S. Fetal Bovine Serum-Derived Extracellular Vesicles Persist within Vesicle-Depleted Culture Media. Int. J. Mol. Sci. 2018, 19, 3538. [Google Scholar] [CrossRef] [Green Version]
- Auber, M.; Fröhlich, D.; Drechsel, O.; Karaulanov, E.; Krämer-Albers, E.-M. Serum-Free Media Supplements Carry miRNAs That Co-Purify with Extracellular Vesicles. J. Extracell. Vesicles 2019, 8, 1656042. [Google Scholar] [CrossRef] [Green Version]
- Liao, Z.; Muth, D.C.; Eitan, E.; Travers, M.; Learman, L.N.; Lehrmann, E.; Witwer, K.W. Serum Extracellular Vesicle Depletion Processes Affect Release and Infectivity of HIV-1 in Culture. Sci. Rep. 2017, 7, 2558. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.G.; Park, U.; Park, B.J.; Kim, K. Exosome-Mediated Bidirectional Signaling between Mesenchymal Stem Cells and Chondrocytes for Enhanced Chondrogenesis. Biotechnol. Bioprocess Eng. 2019, 24, 734–744. [Google Scholar] [CrossRef]
- Wu, J.; Kuang, L.; Chen, C.; Yang, J.; Zeng, W.-N.; Li, T.; Chen, H.; Huang, S.; Fu, Z.; Li, J.; et al. miR-100-5p-Abundant Exosomes Derived from Infrapatellar Fat Pad MSCs Protect Articular Cartilage and Ameliorate Gait Abnormalities via Inhibition of mTOR in Osteoarthritis. Biomaterials 2019, 206, 87–100. [Google Scholar] [CrossRef]
- Shen, K.; Duan, A.; Cheng, J.; Yuan, T.; Zhou, J.; Song, H.; Chen, Z.; Wan, B.; Liu, J.; Zhang, X.; et al. Exosomes Derived from Hypoxia Preconditioned Mesenchymal Stem Cells Laden in a Silk Hydrogel Promote Cartilage Regeneration via the miR-205-5p/PTEN/AKT Pathway. Acta Biomater. 2022, 143, 173–188. [Google Scholar] [CrossRef]
- Lian, C.; Wang, X.; Qiu, X.; Wu, Z.; Gao, B.; Liu, L.; Liang, G.; Zhou, H.; Yang, X.; Peng, Y.; et al. Collagen Type II Suppresses Articular Chondrocyte Hypertrophy and Osteoarthritis Progression by Promoting Integrin β1-SMAD1 Interaction. Bone Res. 2019, 7, 8. [Google Scholar] [CrossRef] [Green Version]
- Aigner, T.; Stöve, J. Collagens—Major Component of the Physiological Cartilage Matrix, Major Target of Cartilage Degeneration, Major Tool in Cartilage Repair. Adv. Drug Deliv. Rev. 2003, 55, 1569–1593. [Google Scholar] [CrossRef]
- Roughley, P.J.; Mort, J.S. The Role of Aggrecan in Normal and Osteoarthritic Cartilage. J. Exp. Orthop. 2014, 1, 8. [Google Scholar] [CrossRef] [Green Version]
- Jenei-Lanzl, Z.; Meurer, A.; Zaucke, F. Interleukin-1β Signaling in Osteoarthritis—Chondrocytes in Focus. Cell. Signal. 2019, 53, 212–223. [Google Scholar] [CrossRef]
- Hu, Q.; Ecker, M. Overview of MMP-13 as a Promising Target for the Treatment of Osteoarthritis. Int. J. Mol. Sci. 2021, 22, 1742. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, Y.-H.; Wu, K.-C.; Ding, D.-C. Chondrogenic Potential of Human Umbilical Cord Mesenchymal Stem Cells Cultured with Exosome-Depleted Fetal Bovine Serum in an Osteoarthritis Mouse Model. Biomedicines 2022, 10, 2773. https://doi.org/10.3390/biomedicines10112773
Chang Y-H, Wu K-C, Ding D-C. Chondrogenic Potential of Human Umbilical Cord Mesenchymal Stem Cells Cultured with Exosome-Depleted Fetal Bovine Serum in an Osteoarthritis Mouse Model. Biomedicines. 2022; 10(11):2773. https://doi.org/10.3390/biomedicines10112773
Chicago/Turabian StyleChang, Yu-Hsun, Kun-Chi Wu, and Dah-Ching Ding. 2022. "Chondrogenic Potential of Human Umbilical Cord Mesenchymal Stem Cells Cultured with Exosome-Depleted Fetal Bovine Serum in an Osteoarthritis Mouse Model" Biomedicines 10, no. 11: 2773. https://doi.org/10.3390/biomedicines10112773