
Plasmacytoma Variant Translocation 1 (PVT1) Gene as a Potential Novel Target for the Treatment of Diabetic Nephropathy


Abstract
:Research in context
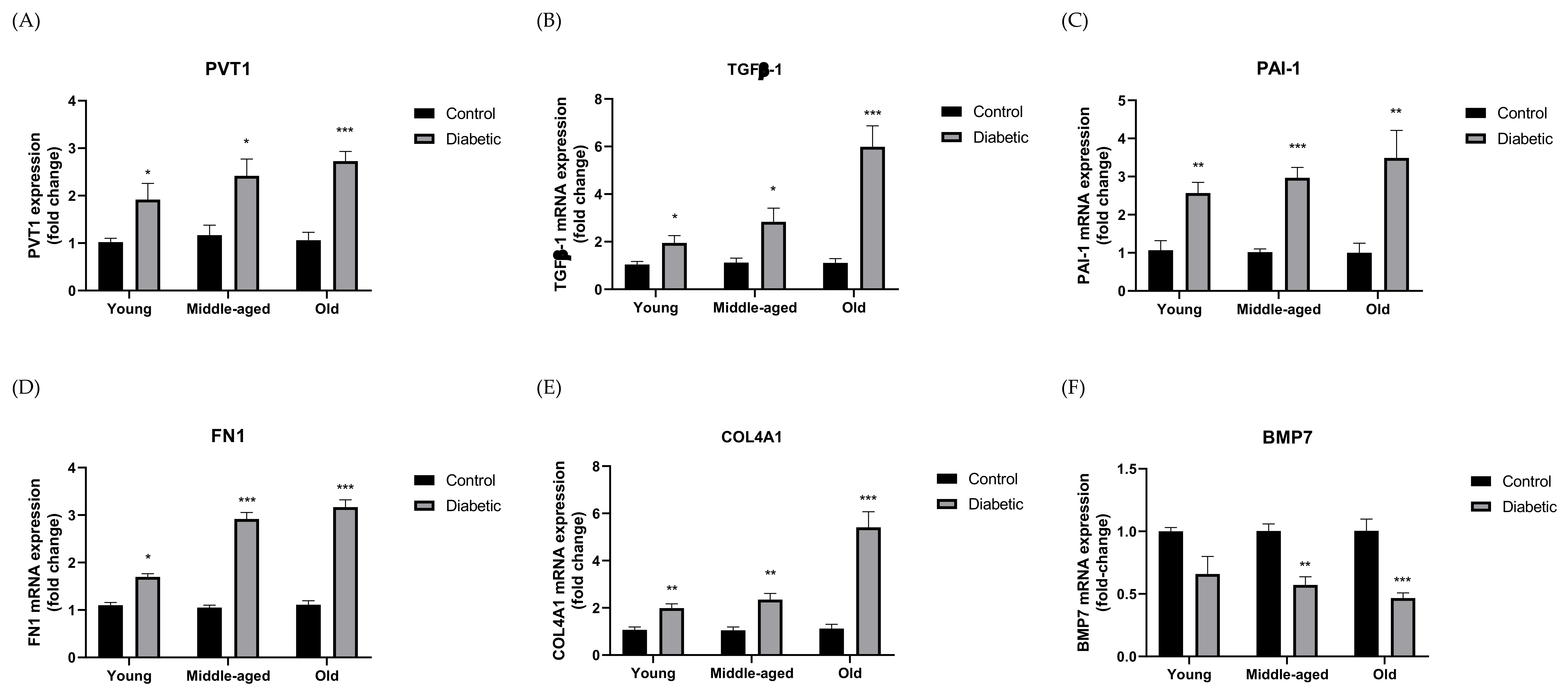
- The noncoding gene PVT1 was highly expressed in mice and patients with DN;
- PVT1 promotes DN through increasing extracellular matrix (ECM) accumulation in vitro;
- Silencing of PVT1 attenuated secretion of ECM proteins and delayed podocyte apoptosis in vitro.
- Does the inhibition of the noncoding gene PVT1 in a diabetic nephropathy mouse model either prevent or delay the onset of diabetic nephropathy?
- Silencing of PVT1 ameliorates DN in terms of kidney function in a diabetic nephropathy mouse model independent of blood glucose change;
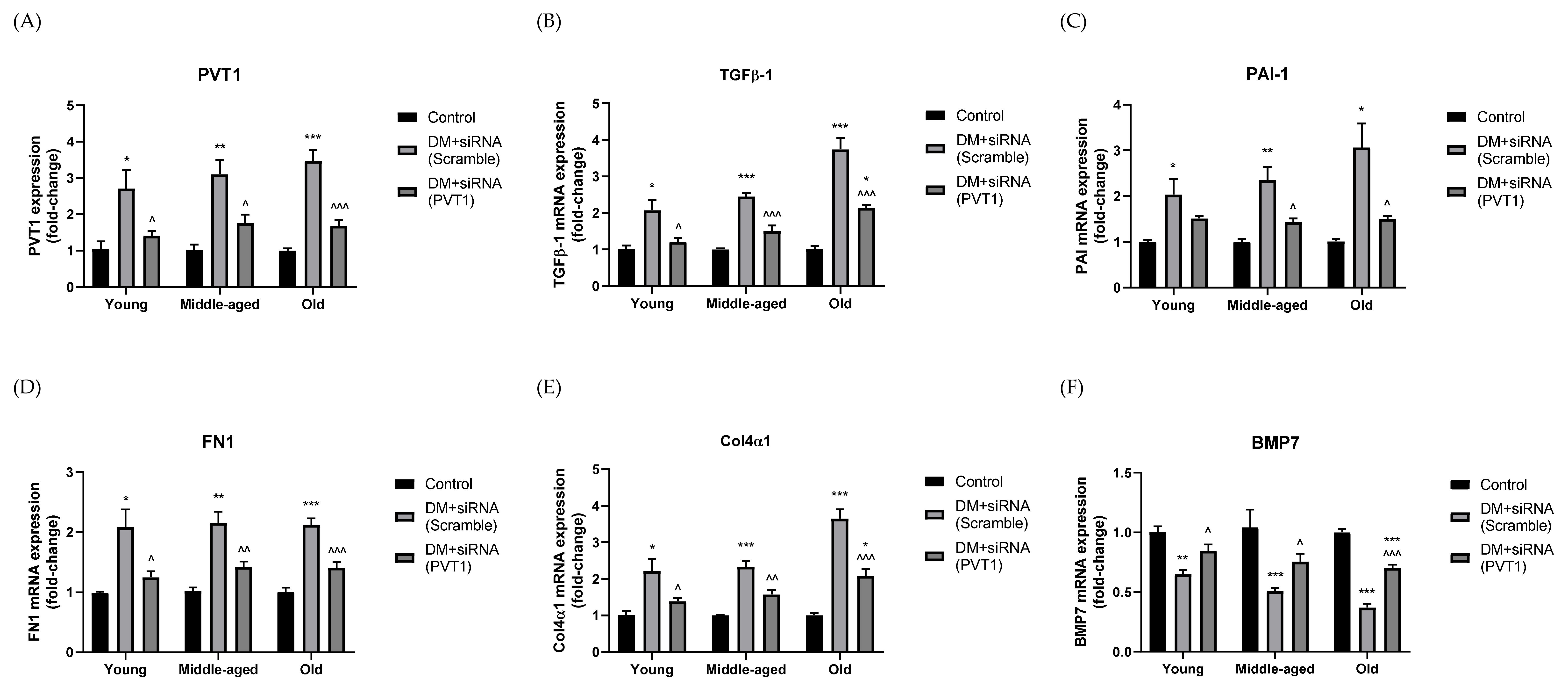
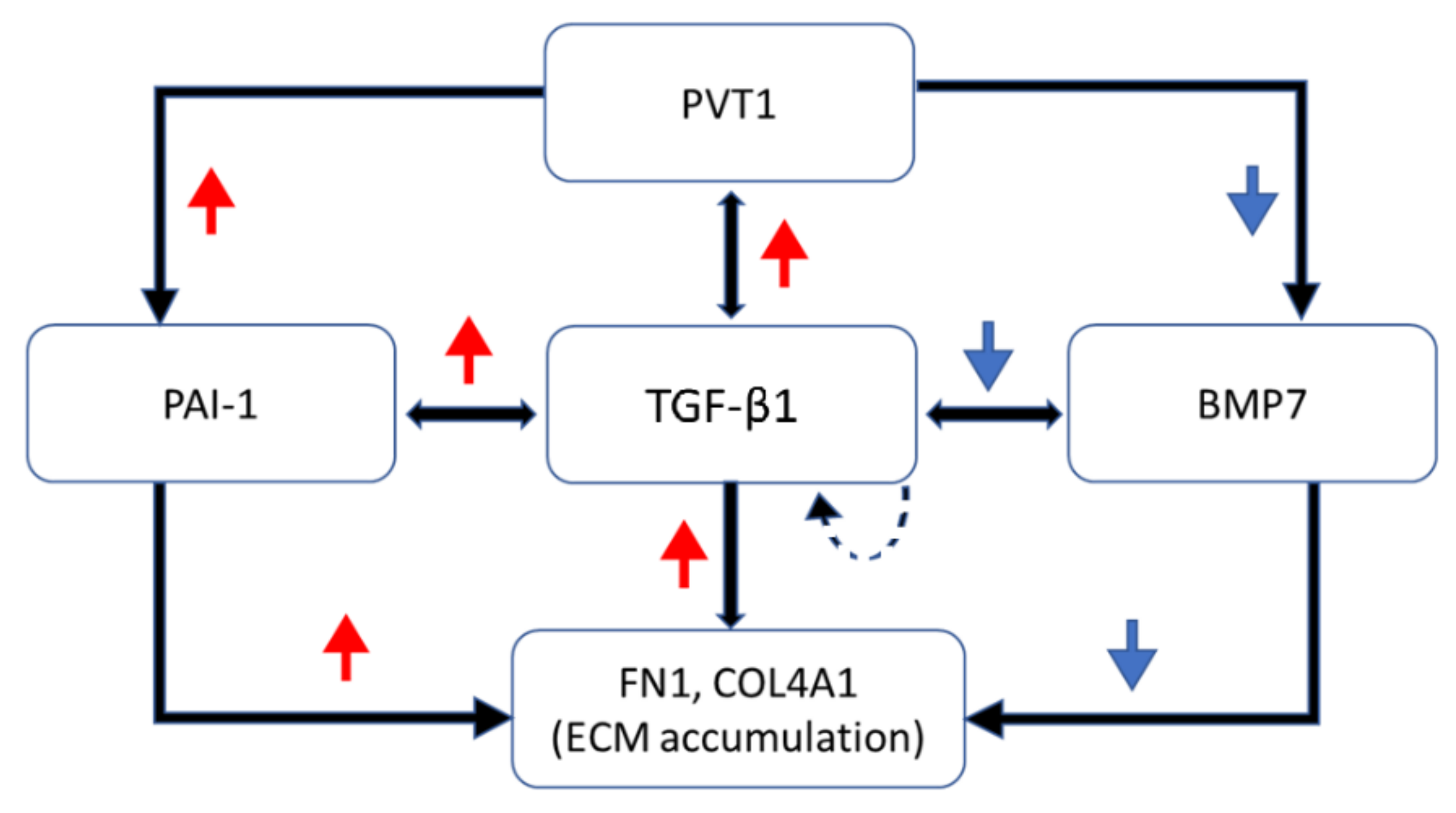
- The mechanism involves downregulating TGF-β1 and PAI-1 while preserving BMP7;
- PVT1 plays an important role in ECM accumulation and is a potential target for the treatment of DN.
- The present study demonstrates the application of RNAi-based therapy to delay DN progression in a murine DN model. RNAi-based therapeutics are emerging treatment strategies in cancers, infectious diseases and single gene disorders. There has already been a clinical trial using siRNA for acute kidney injury, resulting in favorable safety data. Our study provided a foundation for a possible clinical long-term RNAi-based prevention/treatment of DN.
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Ethics Statement
2.3. Blood Glucose, Serum Creatinine, Insulin and TGF-β1 Concentrations
2.4. Urine Albumin, Protein and Creatinine Concentrations
2.5. Calculation of Urine Albumin, Protein and Creatinine Clearance
2.6. Kidney Histological Analysis
2.7. Isolation of Glomeruli
2.8. Gene Expression Analysis
2.9. Statistical Analyses
3. Results
3.1. Physical and Biochemical Characteristics of Murine DN Models
3.2. Expression of PVT1 and ECM Components in Murine DN Models
3.3. Effect of Silencing PVT1 on Blood Glucose, Serum Insulin and TGF-β1
3.4. Effect of Silencing PVT1 on Kidney Hypertrophy and Renal Function
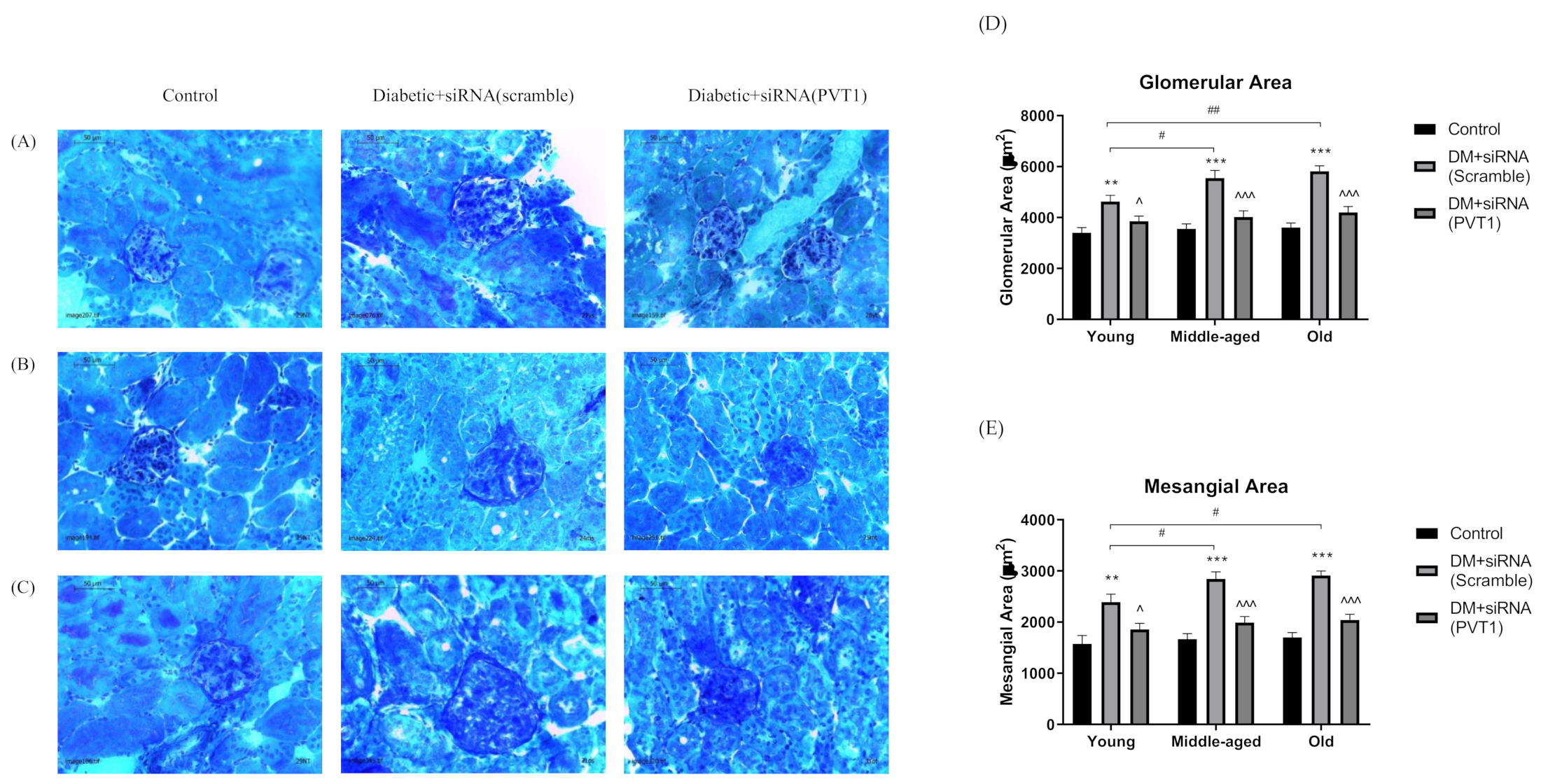
3.5. Effect of Silencing PVT1 on Glomerular and Mesangial Areas
3.6. Effect of Silencing PVT1 on ECM Accumulation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Fineberg, D.; Jandeleit-Dahm, K.A.M.; Cooper, M.E. Diabetic nephropathy: Diagnosis and treatment. Nat. Rev. Endocrinol. 2013, 9, 713–723. [Google Scholar] [CrossRef] [PubMed]
- Kanasaki, K.; Taduri, G.; Koya, D. Diabetic nephropathy: The role of inflammation in fibroblast activation and kidney fibrosis. Front. Endocrinol. 2013, 4, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pugliese, G. Updating the natural history of diabetic nephropathy. Geol. Rundsch. 2014, 51, 905–915. [Google Scholar] [CrossRef]
- Satirapoj, B.; Adler, S.G. Comprehensive approach to diabetic nephropathy. Kidney Res. Clin. Pract. 2014, 33, 121–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Locatelli, F.; Pozzoni, P.; Del Vecchio, L. Renal Replacement Therapy in Patients with Diabetes and End-Stage Renal Disease. J. Am. Soc. Nephrol. 2004, 15, S25–S29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liyanage, T.; Ninomiya, T.; Jha, V.; Neal, B.; Patrice, H.M.; Okpechi, I.; Zhao, M.-h.; Lv, J.; Garg, A.X.; Knight, J.; et al. World-wide access to treatment for end-stage kidney disease: A systematic review. Lancet 2015, 385, 1975–1982. [Google Scholar] [CrossRef]
- Schieppati, A.; Remuzzi, G. Chronic renal diseases as a public health problem: Epidemiology, social, and economic implications. Kidney Int. 2005, 68, S7–S10. [Google Scholar] [CrossRef] [Green Version]
- Silva Junior, G.B.D.; Oliveira, J.G.R.D.; Oliveira, M.R.B.D.; Vieira, L.J.E.D.S.; Dias, E.R. Global costs attributed to chronic kidney disease: A systematic review. Rev. Assoc. Médica Bras. 2018, 64, 1108–1116. [Google Scholar] [CrossRef]
- Susan van, D.; Beulens, J.W.; Yvonne, T.; van der, S.; Grobbee, D.E.; Nealb, B. The global burden of diabetes and its complications: An emerging pandemic. Eur. J. Cardiovasc. Prev. Rehabil. 2010, 17 (Suppl. S1), S3–S8. [Google Scholar] [CrossRef]
- Rizvi, S.; Raza, S.T.; Mahdi, F. Association of genetic variants with diabetic nephropathy. World J. Diabetes 2014, 5, 809. [Google Scholar] [CrossRef]
- Hanson, R.L.; Craig, D.W.; Millis, M.P.; Yeatts, K.A.; Kobes, S.; Pearson, J.V.; Lee, A.M.; Knowler, W.C.; Nelson, R.G.; Wolford, J.K. Identification of PVT1 as a Candidate Gene for End-Stage Renal Disease in Type 2 Diabetes Using a Pooling-Based Genome-Wide Single Nucleotide Polymorphism Association Study. Diabetes 2007, 56, 975–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millis, M.P.; Bowen, D.; Kingsley, C.; Watanabe, R.M.; Wolford, J.K. Variants in the Plasmacytoma Variant Translocation Gene (PVT1) Are Associated With End-Stage Renal Disease Attributed to Type 1 Diabetes. Diabetes 2007, 56, 3027–3032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, G.-C.; Pan, H.-F.; Leng, R.-X.; Wang, D.-G.; Li, X.-P.; Li, X.-M.; Ye, D.-Q. Emerging role of long noncoding RNAs in autoimmune diseases. Autoimmun. Rev. 2015, 14, 798–805. [Google Scholar] [CrossRef]
- Srivastava, S.; Goodwin, J.; Tripathi, P.; Kanasaki, K.; Koya, D. Interactions among Long Non-Coding RNAs and microRNAs Influence Disease Phenotype in Diabetes and Diabetic Kidney Disease. Int. J. Mol. Sci. 2021, 22, 6027. [Google Scholar] [CrossRef] [PubMed]
- Onagoruwa, O.T.; Pal, G.; Ochu, C.; Ogunwobi, O.O. Oncogenic Role of PVT1 and Therapeutic Implications. Front. Oncol. 2020, 10, 17. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.-W.; Zhang, J.-H.; Liu, F.-X.; Wang, X.-T.; Pan, S.-K.; Jiang, D.-K.; Zhao, Z.-H.; Liu, Z.-S. Silencing of long noncoding RNA PVT1 inhibits podocyte damage and apoptosis in diabetic nephropathy by upregulating FOXA1. Exp. Mol. Med. 2019, 51, 1–15. [Google Scholar] [CrossRef]
- Zhong, W.; Zeng, J.; Xue, J.; Du, A.; Xu, Y. Knockdown of lncRNA PVT1 alleviates high glucose-induced proliferation and fibrosis in human mesangial cells by miR-23b-3p/WT1 axis. Diabetol. Metab. Syndr. 2020, 12, 33. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, M.L.; DiStefano, J.K. Functional Characterization of the Plasmacytoma Variant Translocation 1 Gene (PVT1) in Diabetic Nephropathy. PLoS ONE 2011, 6, e18671. [Google Scholar] [CrossRef] [Green Version]
- Alvarez, M.L.; Khosroheidari, M.; Eddy, E.; Kiefer, J. Role of MicroRNA 1207-5P and Its Host Gene, the Long Non-Coding RNA Pvt1, as Mediators of Extracellular Matrix Accumulation in the Kidney: Implications for Diabetic Nephropathy. PLoS ONE 2013, 8, e77468. [Google Scholar] [CrossRef] [Green Version]
- Yu, D.; Yang, X.; Zhu, Y.; Xu, F.; Zhang, H.; Qiu, Z. Knockdown of plasmacytoma variant translocation 1 (PVT1) inhibits high glucose-induced proliferation and renal fibrosis in HRMCs by regulating miR-23b-3p/early growth response factor 1 (EGR1). Endocr. J. 2021, 68, 519–529. [Google Scholar] [CrossRef]
- King, A.J. The use of animal models in diabetes research. J. Cereb. Blood Flow Metab. 2012, 166, 877–894. [Google Scholar] [CrossRef] [Green Version]
- Weston, B.S.; Wahab, N.A.; Mason, R.M. CTGF Mediates TGF-β–Induced Fibronectin Matrix Deposition by Upregulating Active α5β1 Integrin in Human Mesangial Cells. J. Am. Soc. Nephrol. 2003, 14, 601–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, K.; Ziyadeh, F.N. Renal hypertrophy is associated with upregulation of TGF-beta 1 gene expression in diabetic BB rat and NOD mouse. Am. J. Physiol. Physiol. 1994, 267, F1001–F1094. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Jiang, S.; Shang, J.; Jiang, Y.; Dai, Y.; Xu, B.; Yu, Y.; Liang, Z.; Yang, Y. LncRNA: Shedding light on mechanisms and opportunities in fibrosis and aging. Ageing Res. Rev. 2019, 52, 17–31. [Google Scholar] [CrossRef]
- Khorkova, O.; Hsiao, J.; Wahlestedt, C. Basic biology and therapeutic implications of lncRNA. Adv. Drug Deliv. Rev. 2015, 87, 15–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicholas, S.B.; Aguiniga, E.; Ren, Y.; Kim, J.; Wong, J.; Govindarajan, N.; Noda, M.; Wang, W.; Kawano, Y.; Collins, A.; et al. Plasminogen activator inhibitor-1 deficiency retards diabetic nephropathy. Kidney Int. 2005, 67, 1297–1307. [Google Scholar] [CrossRef] [Green Version]
- Samarakoon, R.; Overstreet, J.M.; Higgins, P.J. TGF-β signaling in tissue fibrosis: Redox controls, target genes and therapeutic opportunities. Cell. Signal. 2013, 25, 264–268. [Google Scholar] [CrossRef] [Green Version]
- Böttinger, E.P. TGF-β in renal injury and disease. In Seminars in Nephrology; Elsevier: Amsterdam, The Netherlands, 2007. [Google Scholar]
- Patel, S.R.; Dressler, G.R. BMP7 signaling in renal development and disease. Trends Mol. Med. 2005, 11, 512–518. [Google Scholar] [CrossRef]
- Yokoi, H.; Mukoyama, M.; Sugawara, A.; Mori, K.; Nagae, T.; Makino, H.; Suganami, T.; Yahata, K.; Fujinaga, Y.; Tanaka, I.; et al. Role of connective tissue growth factor in fibronectin expression and tubulointerstitial fibrosis. Am. J. Physiol. Physiol. 2002, 282, F933–F942. [Google Scholar] [CrossRef] [Green Version]
- Cao, L.; Qin, P.; Zhang, J.; Qiao, H.; Shi, P.; Huo, H. LncRNA PVT1 Suppresses the Progression of Renal Fibrosis via Inactivation of TGF-β Signaling Pathway. Drug Des. Dev. Ther. 2020, 14, 3547–3557. [Google Scholar] [CrossRef]
- Wong, M.G.; Perkovic, V.; Woodward, M.; Chalmers, J.; Li, Q.; Hillis, G.S.; Azari, D.Y.; Jun, M.; Poulter, N.; Hamet, P.; et al. Circulating bone morphogenetic protein-7 and transforming growth factor-β1 are better predictors of renal end points in patients with type 2 diabetes mellitus. Kidney Int. 2013, 83, 278–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Niu, Q.; Liang, K.; Li, X.; Jiang, J.; Bian, C. Effect of LncPVT1/miR-20a-5p on Lipid Metabolism and Insulin Resistance in NAFLD. Diabetes Metab. Syndr. Obes. Targets Ther. 2021, 14, 4599. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, D.; Qin, Z.; Li, J.; Shen, Z. The role and possible mechanism of long noncoding RNA PVT1 in modulating 3T3-L1 preadipocyte proliferation and differentiation. IUBMB Life 2020, 72, 1460–1467. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, M.L.; DiStefano, J.K. The role of non-coding RNAs in diabetic nephropathy: Potential applications as biomarkers for disease development and progression. Diabetes Res. Clin. Pract. 2013, 99, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Bilous, R. KDOQI Clinical Practice Guideline for Diabetes and CKD: 2012 Update. Am. J. Kidney Dis. 2012, 60, 850–886. [Google Scholar] [CrossRef]
- Abrass, C.K.; Spicer, D.; Raugi, G.J. Insulin induces a change in extracellular matrix glycoproteins synthesized by rat mesangial cells in culture. Kidney Int. 1994, 46, 613–620. [Google Scholar] [CrossRef] [Green Version]
- Alpers, C.E.; Hudkins, K.L. Mouse models of diabetic nephropathy. Curr. Opin. Nephrol. Hypertens. 2011, 20, 278. [Google Scholar] [CrossRef]
- Azushima, K.; Gurley, S.B.; Coffman, T.M. Modelling diabetic nephropathy in mice. Nat. Rev. Nephrol. 2018, 14, 48–56. [Google Scholar] [CrossRef]
- Giralt-López, A.; Bosch, M.M.-V.D.; Vergara, A.; García-Carro, C.; Seron, D.; Jacobs-Cacha, C.; Soler, M.J. Revisiting Experimental Models of Diabetic Nephropathy. Int. J. Mol. Sci. 2020, 21, 3587. [Google Scholar] [CrossRef]
- Tesch, G.H.; Allen, T. Rodent models of streptozotocin-induced diabetic nephropathy (Methods in Renal Research). Nephrology 2007, 12, 261–266. [Google Scholar] [CrossRef]
- Magee, C.; Grieve, D.J.; Watson, C.J.; Brazil, D. Diabetic Nephropathy: A Tangled Web to Unweave. Cardiovasc. Drugs Ther. 2017, 31, 579–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juin, S.K.; Pushpakumar, S.; Sen, U. GYY4137 Regulates Extracellular Matrix Turnover in the Diabetic Kidney by Modulating Retinoid X Receptor Signaling. Biomolecules 2021, 11, 1477. [Google Scholar] [CrossRef] [PubMed]
- Qin, B.; Cao, X. LncRNA PVT1 Regulates High Glucose-Induced Viability, Oxidative Stress, Fibrosis, and Inflammation in Diabetic Nephropathy via miR-325-3p/Snail1 Axis. Diabetes, Metab. Syndr. Obes. Targets Ther. 2021, 14, 1741–1750. [Google Scholar] [CrossRef] [PubMed]
- Du, H.; Zhang, H.; Yang, R.; Qiao, L.; Shao, H.; Zhang, X. Small interfering RNA-induced silencing lncRNA PVT1 inhibits atherosclerosis via inactivating the MAPK/NF-κB pathway. Aging 2021, 13, 24449. [Google Scholar] [CrossRef]
- Watts, J.K.; Corey, D.R. Clinical status of duplex RNA. Bioorganic Med. Chem. Lett. 2010, 20, 3203–3207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burnett, J.C.; Rossi, J.J.; Tiemann, K. Current progress of siRNA/shRNA therapeutics in clinical trials. Biotechnol. J. 2011, 6, 1130–1146. [Google Scholar] [CrossRef] [Green Version]
- Demirjian, S.; Ailawadi, G.; Polinsky, M.; Bitran, D.; Silberman, S.; Shernan, S.K.; Burnier, M.; Hamilton, M.; Squiers, E.; Erlich, S.; et al. Safety and Tolerability Study of an Intravenously Administered Small Interfering Ribonucleic Acid (siRNA) Post On-Pump Cardiothoracic Surgery in Patients at Risk of Acute Kidney Injury. Kidney Int. Rep. 2017, 2, 836–843. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Young (9 Weeks) | Middle-Aged (16 Weeks) | Old (24 Weeks) | ||||
|---|---|---|---|---|---|---|
| Control | Diabetic | Control | Diabetic | Control | Diabetic | |
| Body weight (g) | 26.090 ± 0.580 | 28.030 ± 0.560 * | 28.530 ± 0.460 | 30.410 ± 0.600 * | 31.850 ± 0.900 | 34.120 ± 1.530 |
| Body weight change (%) | 32.990 ± 3.270 | 46.200 ± 3.830 * | 48.590 ± 4.560 | 66.360 ± 5.760 * | 67.850 ± 4.900 | 80.040 ± 10.470 |
| Blood glucose (mmol/l) | 10.83 ± 0.41 | 25.61 ± 0.42 *** | 12.44 ± 0.41 | 28.37 ± 0.90 *** | 11.58 ± 0.26 | 27.07 ± 1.79 *** |
| Kidney weight (g) | 0.228 ± 0.011 | 0.268 ± 0.022 | 0.234 ± 0.009 | 0.285 ± 0.018 * | 0.238 ± 0.008 | 0.275 ± 0.015 * |
| Kidney to body weight ratio | 0.009 ± 0.000 | 0.001 ± 0.000 | 0.008 ± 0.000 | 0.009 ± 0.000 * | 0.008 ± 0.000 | 0.008 ± 0.000 |
| Serum creatinine (mg/dl) | 0.53 ± 0.03 | 0.81 ± 0.09 * | 0.57 ± 0.04 | 0.93 ± 0.12 * | 0.58 ± 0.04 | 0.95 ± 0.14 * |
| Creatinine clearance (ml/min) | 0.022 ± 0.002 | 0.020 ± 0.003 | 0.0220 ± 0.002 | 0.014 ± 0.002 * | 0.021 ± 0.003 | 0.012 ± 0.001 *, # |
| UAE (μg) | 9.680 ± 1.200 | 19.940 ± 2.670 ** | 11.100 ± 2.670 | 33.230 ± 2.940 ***, # | 15.360 ± 2.470 | 50.550 ± 4.910 ***, ### |
| UACR | 0.064 ± 0.010 | 0.098 ± 0.011 * | 0.073 ± 0.0047 | 0.152 ± 0.011 ***, # | 0.088 ± 0.005 | 0.219 ± 0.018 ***, ### |
| UPE (mg) | 0.527 ± 0.055 | 0.799 ± 0.133 | 0.520 ± 0.123 | 0.968 ± 0.125 * | 0.530 ± 0.069 | 1.074 ± 0.134 * |
| UPCR | 3.093 ± 0.090 | 4.072 ± 0.421 * | 3.051 ± 0.343 | 4.288 ± 0.325 * | 2.911 ± 0.308 | 4.420 ± 0.306 ** |
| Young (9 Weeks) | Middle-Aged (16 Weeks) | Old (24 Weeks) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Control | DM + siRNA (Scramble) | DM + siRNA (PVT1) | Control | DM + siRNA (Scramble) | DM + siRNA (PVT1) | Control | DM + siRNA (Scramble) | DM + siRNA (PVT1) | |
| Body weight (g) | 22.930 ± 0.230 | 26.600 ± 0.860 * | 26.230 ± 0.470 * | 27.630 ± 0.760 | 30.300 ± 0.460 * | 28.000 ± 0.530 ^ | 28.370 ± 0.410 | 29.820 ± 0.500 | 30.170 ± 0.310 |
| Body weight change (%) | 25.350 ± 1.310 | 40.490 ± 1.400 *** | 34.580 ± 1.210 **, ^ | 47.460 ± 3.600 | 70.670 ± 2.620 ** | 50.590 ± 3.810 ^^ | 92.390 ± 4.180 | 101.730 ± 9.520 | 96.130 ± 2.500 |
| Blood glucose (mmol/l) | 12.37 ± 0.67 | 25.42 ± 0.77 *** | 23.47 ± 0.42 *** | 12.10 ± 0.35 | 30.27 ± 0.35 *** | 28.82 ± 0.36 ***, ^ | 11.90 ± 0.44 | 28.38 ± 0.52 *** | 26.74 ± 0.50 *** |
| Kidney weight (g) | 0.252 ± 0.002 | 0.258 ± 0.008 | 0.240 ± 0.005 | 0.236 ± 0.012 | 0.266 ± 0.012 | 0.250 ± 0.009 | 0.240 ± 0.011 | 0.269 ± 0.005 | 0.262 ± 0.004 |
| Kidney to body weight ratio | 0.010 ± 0.000 | 0.010 ± 0.001 | 0.009 ± 0.000 | 0.009 ± 0.000 | 0.011 ± 0.000 ** | 0.0090 ± 0.000 ^^ | 0.009 ± 0.000 | 0.011 ± 0.000 ** | 0.008 ± 0.000 ^^^ |
| Serum TGF-β1 (ng/ml) | 72.95 ± 2.58 | 90.69 ± 3.58 ** | 76.31 ± 2.17 ^ | 78.61 ± 4.38 | 96.70 ± 2.72 * | 83.34 ± 2.76 ^ | 78.61 ± 4.38 | 96.70 ± 2.72 **, # | 83.34 ± 2.76 ^, @ |
| Serum insulin (ng/ml) | 1.386 ± 0.202 | 0.241 ± 0.140 ** | 0.750 ± 0.193 | 1.396 ± 0.249 | 0.299 ± 0.086 ** | 0.626 ± 0.204 * | 1.148 ± 0.357 | 0.120 ± 0.026 ** | 0.274 ± 0.121 * |
| Serum creatinine (mg/dl) | 0.667 ± 0.037 | 1.015 ± 0.001 *** | 0.836 ± 0.031 ^ | 0.702 ± 0.062 | 1.318 ± 0.102 *** | 0.991 ± 0.063 ^ | 0.700 ± 0.094 | 1.168 ± 0.069 ** | 0.936 ± 0.087 |
| Creatinine clearance (ml/min) | 0.032 ± 0.002 | 0.028 ± 0.002 | 0.031 ± 0.004 | 0.032 ± 0.002 | 0.020 ± 0.002 *, # | 0.028 ± 0.003 | 0.031 ± 0.004 | 0.016 ± 0.001 **, ## | 0.025 ± 0.002 ^ |
| UAE (μg) | 11.760 ± 0.290 | 25.923 ± 1.043 *** | 16.253 ± 1.644 ^^^ | 13.071 ± 0.397 | 38.817 ± 3.094 ***, # | 22.272 ± 2.911 ^^ | 14.518 ± 0.481 | 44.337 ± 4.255 ***, ## | 24.632 ± 2.278 ^^ |
| UACR | 0.067 ± 0.009 | 0.135 ± 0.015 ** | 0.069 ± 0.008 ^^ | 0.081 ± 0.006 | 0.162 ± 0.014 **, # | 0.110 ± 0.017 ^ | 0.094 ± 0.016 | 0.233 ± 0.021 ***, ## | 0.132 ± 0.011 ^^, @@ |
| UPE (mg) | 0.808 ± 0.051 | 0.989 ± 0.068 | 0.834 ± 0.041 | 0.913 ± 0.090 | 1.679 ± 0.084 **, # | 1.015 ± 0.150 ^^ | 1.046 ± 0.089 | 2.067 ± 0.198 **, ### | 1.433 ± 0.175 ^, @ |
| UPCR | 2.845 ± 0.184 | 3.331 ± 0.165 | 2.997 ± 0.219 | 2.833 ± 0.160 | 3.569 ± 0.217 * | 3.138 ± 0.099 | 2.873 ± 0.249 | 3.976 ± 0.315 * | 3.127 ± 0.177 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mok, H.; Al-Jumaily, A.; Lu, J. Plasmacytoma Variant Translocation 1 (PVT1) Gene as a Potential Novel Target for the Treatment of Diabetic Nephropathy. Biomedicines 2022, 10, 2711. https://doi.org/10.3390/biomedicines10112711
Mok H, Al-Jumaily A, Lu J. Plasmacytoma Variant Translocation 1 (PVT1) Gene as a Potential Novel Target for the Treatment of Diabetic Nephropathy. Biomedicines. 2022; 10(11):2711. https://doi.org/10.3390/biomedicines10112711
Chicago/Turabian StyleMok, Helen, Ahmed Al-Jumaily, and Jun Lu. 2022. "Plasmacytoma Variant Translocation 1 (PVT1) Gene as a Potential Novel Target for the Treatment of Diabetic Nephropathy" Biomedicines 10, no. 11: 2711. https://doi.org/10.3390/biomedicines10112711