
Differential Affinity Chromatography Coupled to Mass Spectrometry: A Suitable Tool to Identify Common Binding Proteins of a Broad-Range Antimicrobial Peptide Derived from Leucinostatin

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Tissue Culture Media, Biochemicals and Compounds
2.2. In Vitro Culture, Collection of Parasite and Efficacy Assessments In Vitro
2.3. In Vivo Assessment of Peptide 6027 for the Treatment of Mice Experimentally Infected with T. gondii Oocysts
2.4. Assessment of Susceptibility of Murine Splenocytes to Peptide 6027
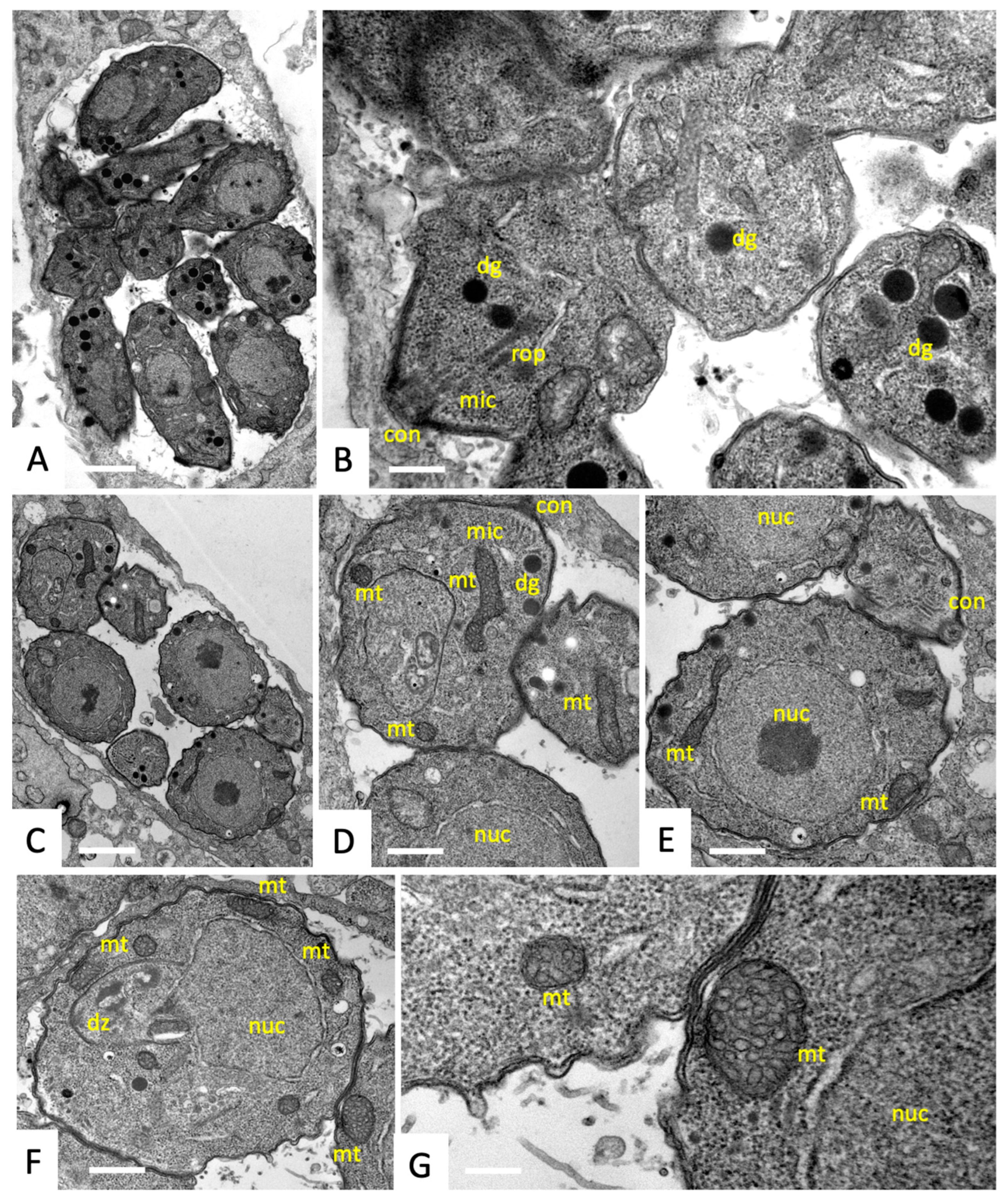
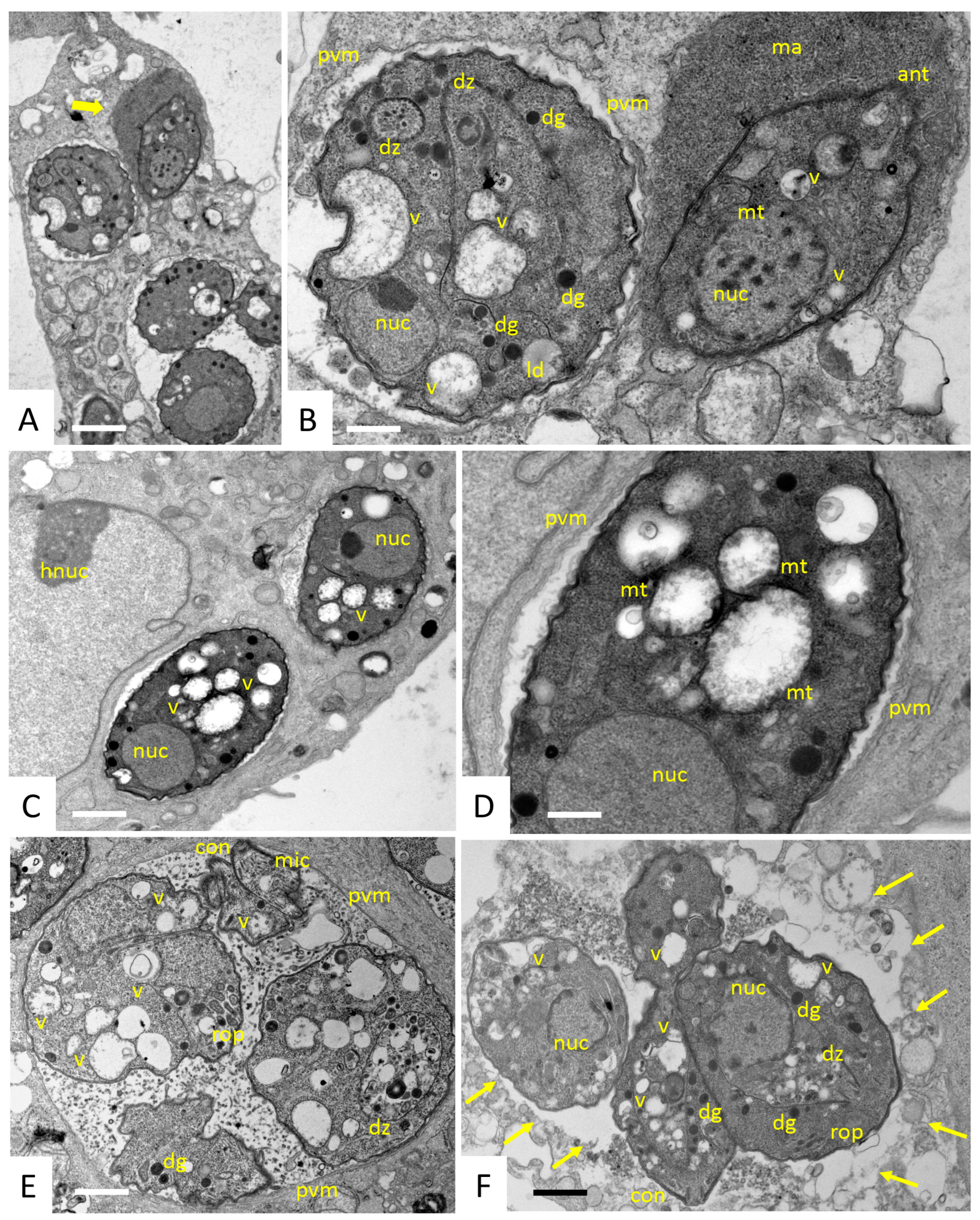
2.5. Transmission Electron Microscopy (TEM)
2.6. Protein Extraction and Differential Affinity Chromatography (DAC)
2.7. Proteomic Analysis of the Eluted Proteins by Mass Spectrometry
3. Results
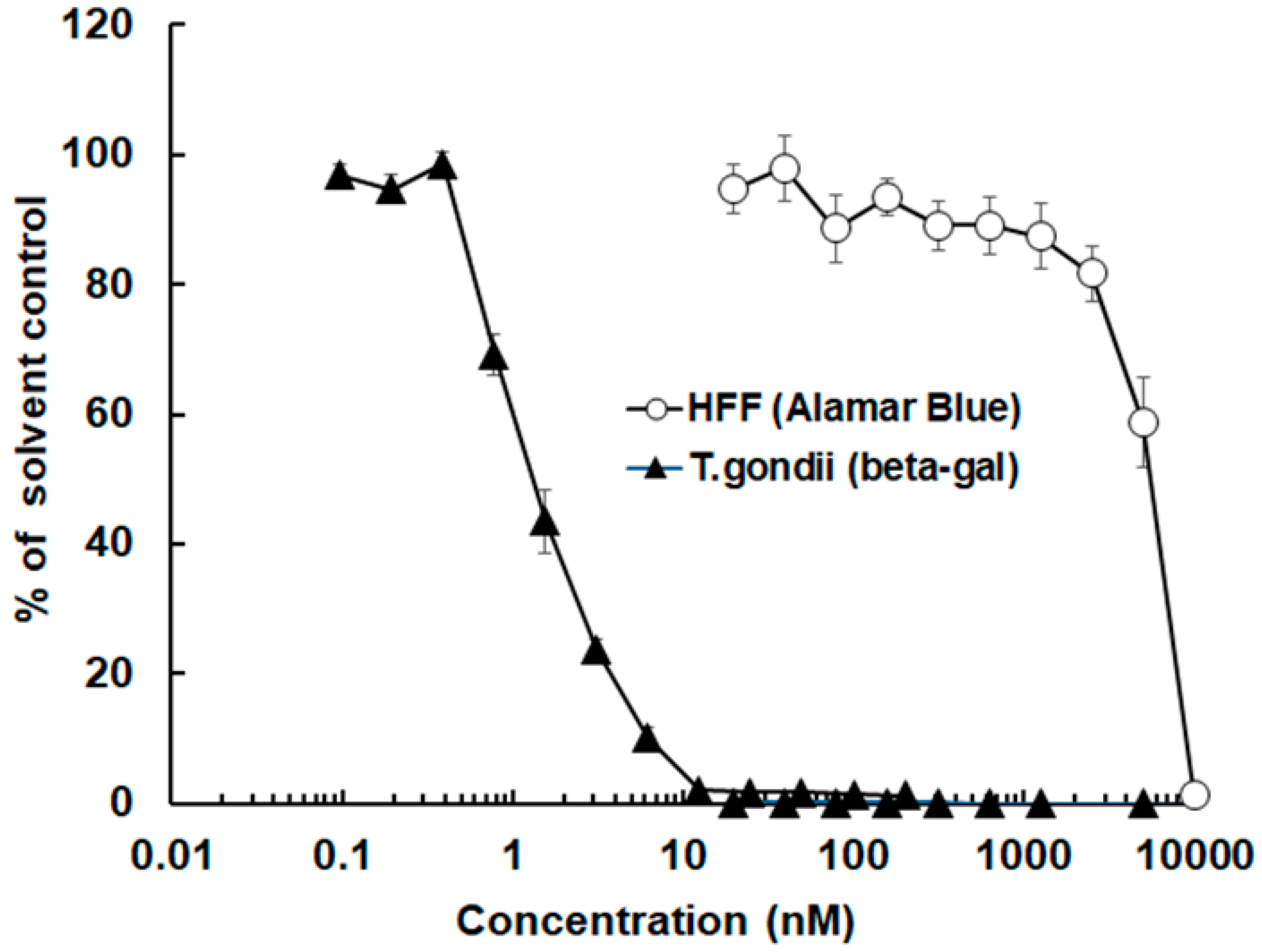
3.1. The Antimicrobial Peptide 6027 Selectively Impairs the Proliferation and Structural Integrity of T. gondii Tachyzoites In Vitro
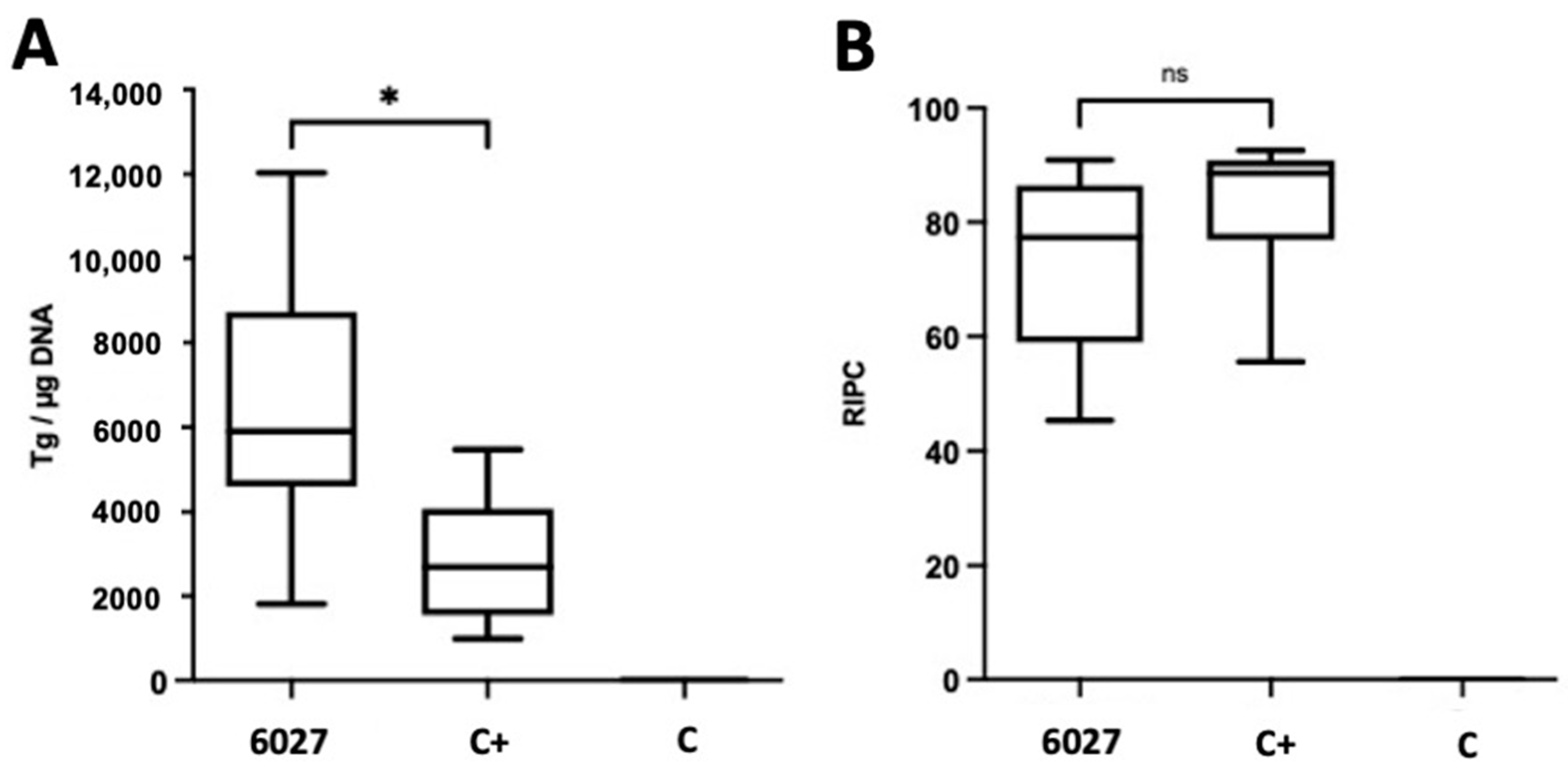
3.2. Peptide 6027 Has Detrimental Effects When Applied to CD1 Mice Orally Infected with T. gondii Oocysts
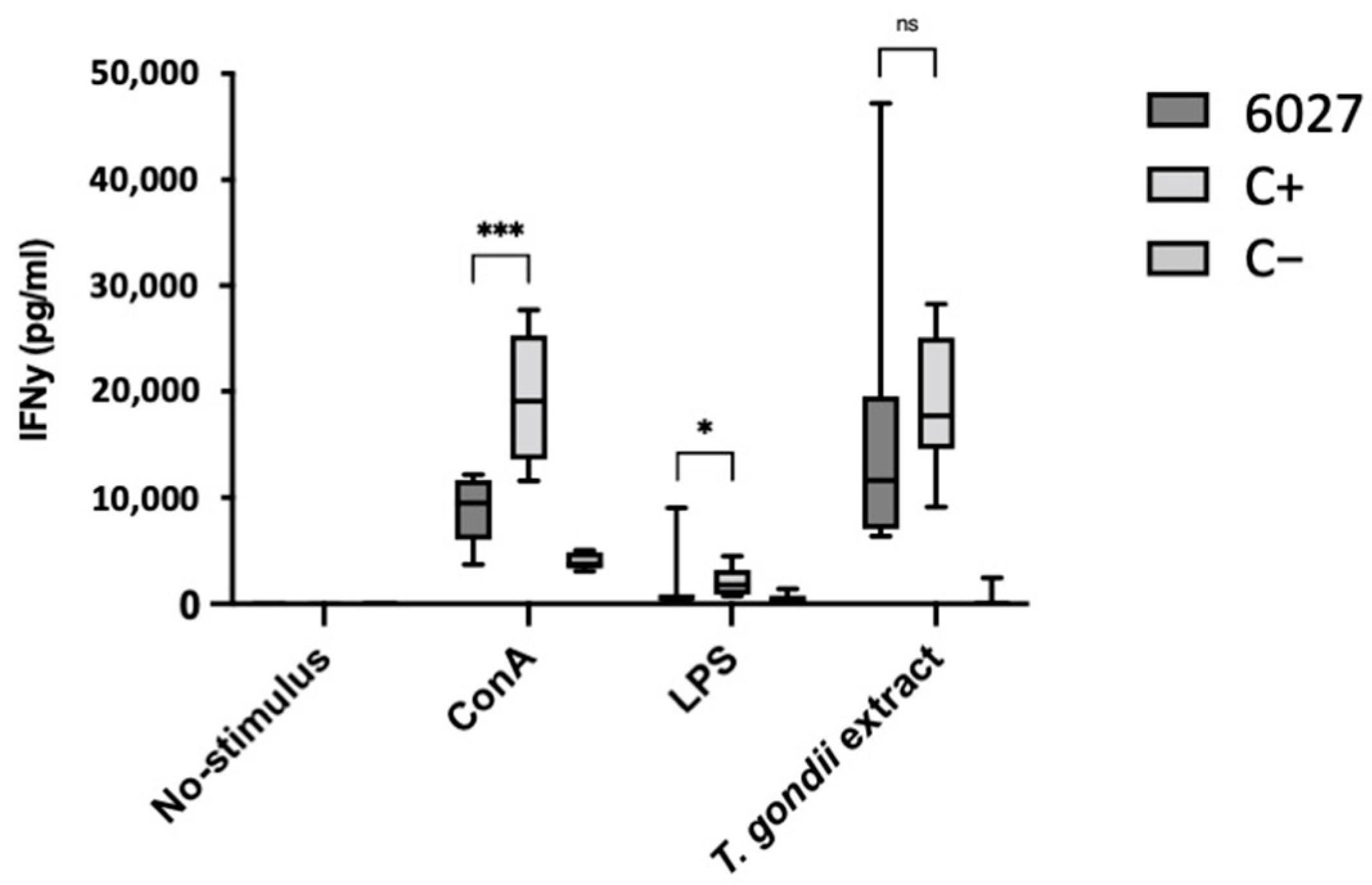
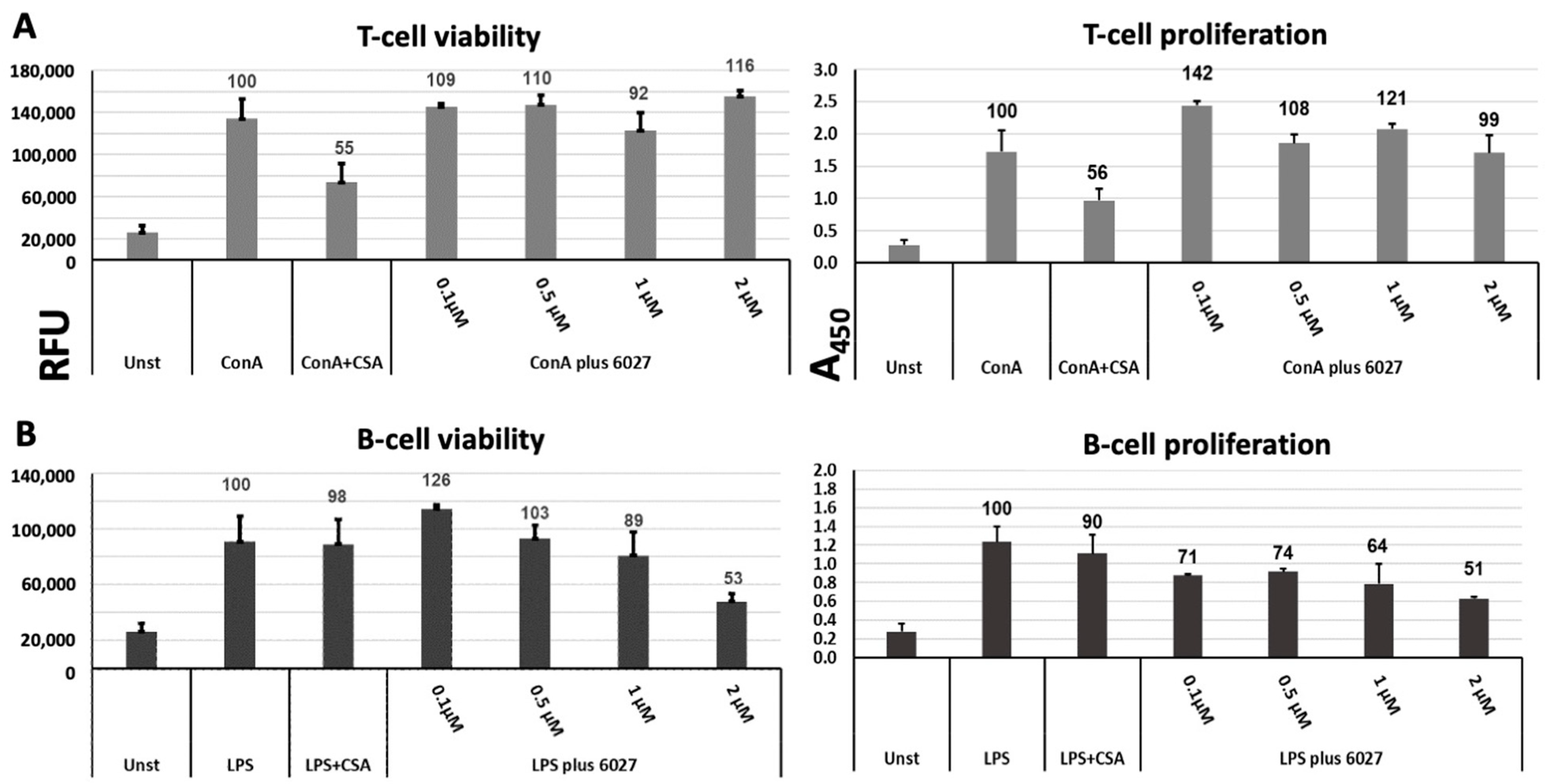
3.3. Peptide 6027 Affects the Proliferative Capacity and Viability of Murine B Cells
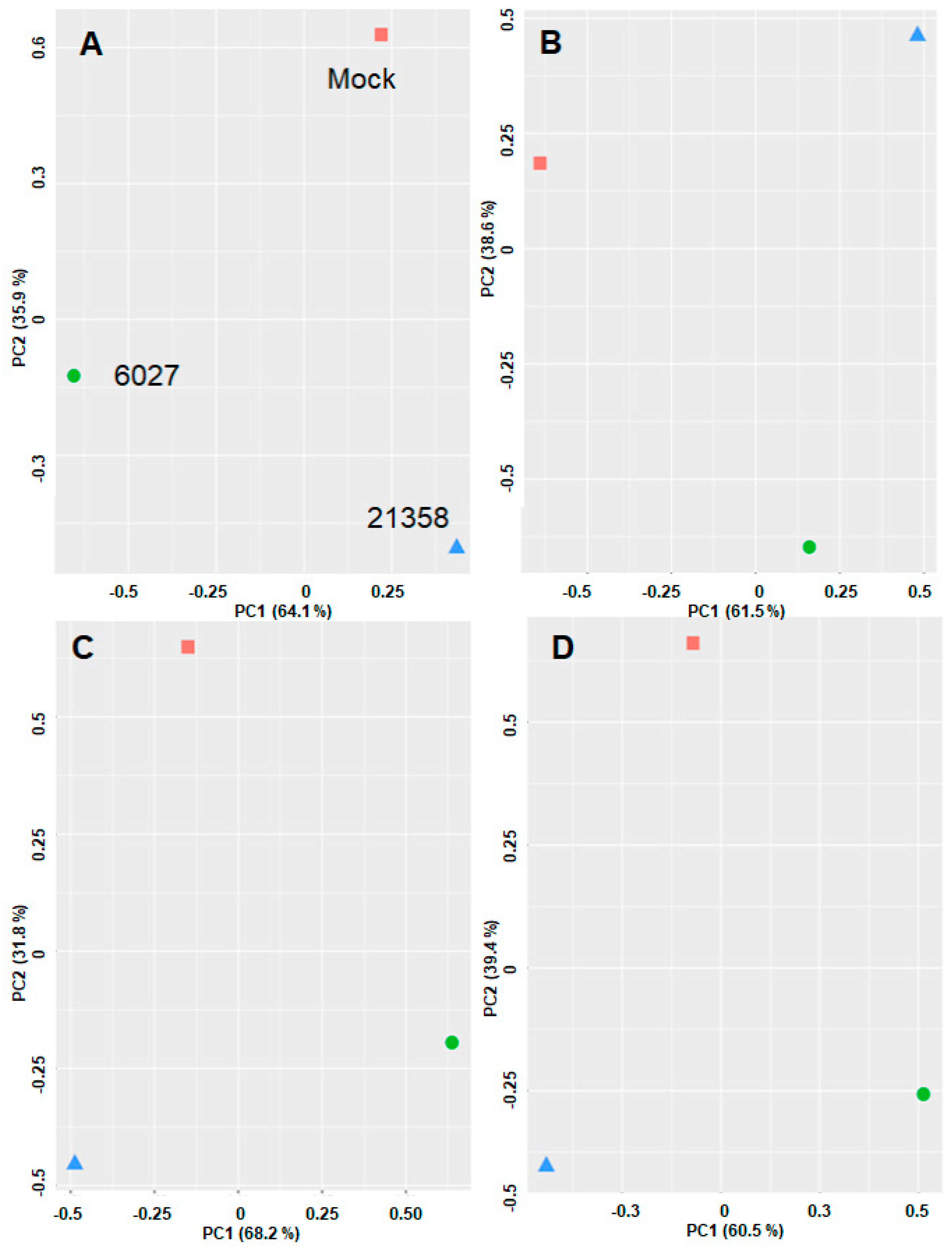
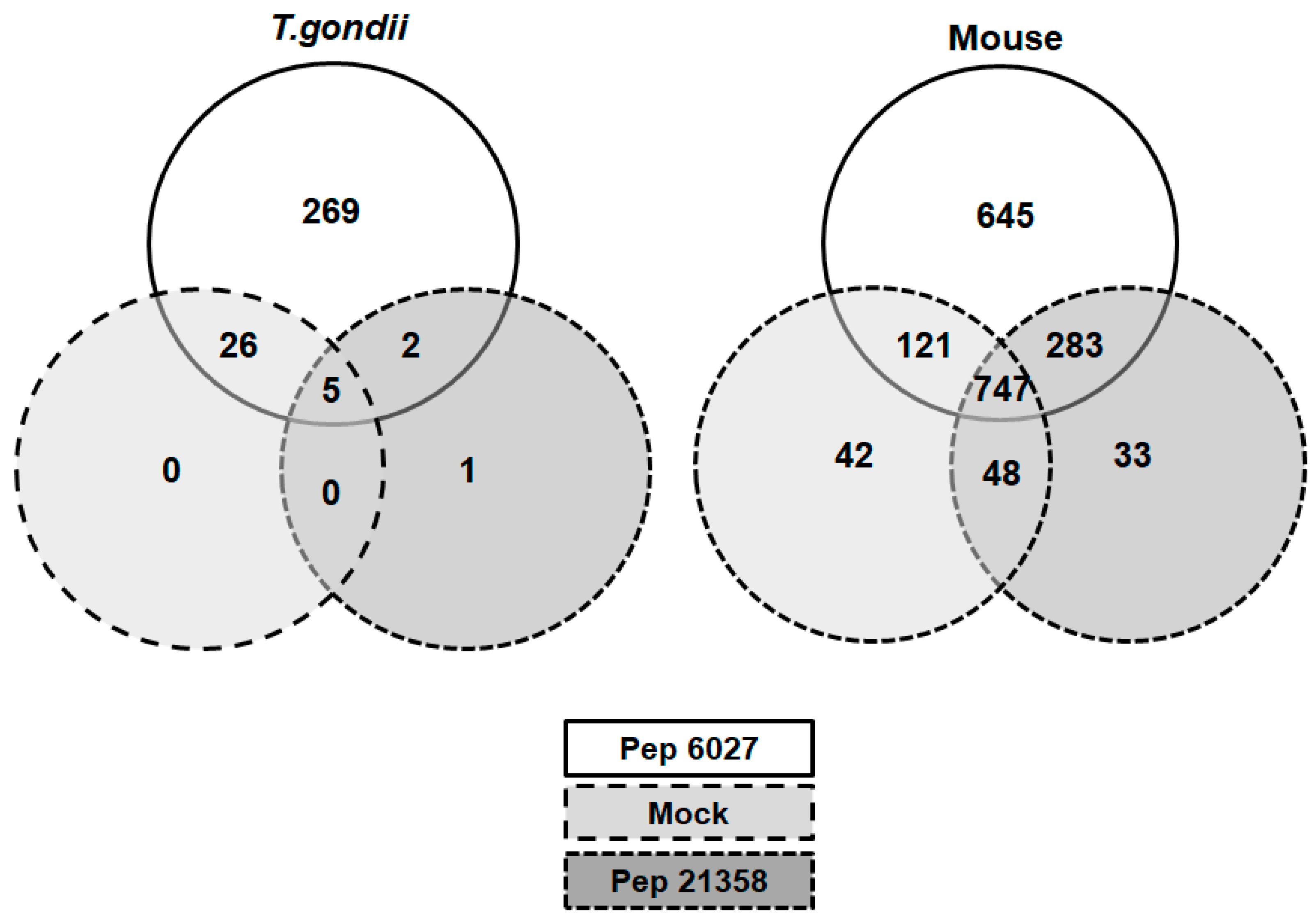
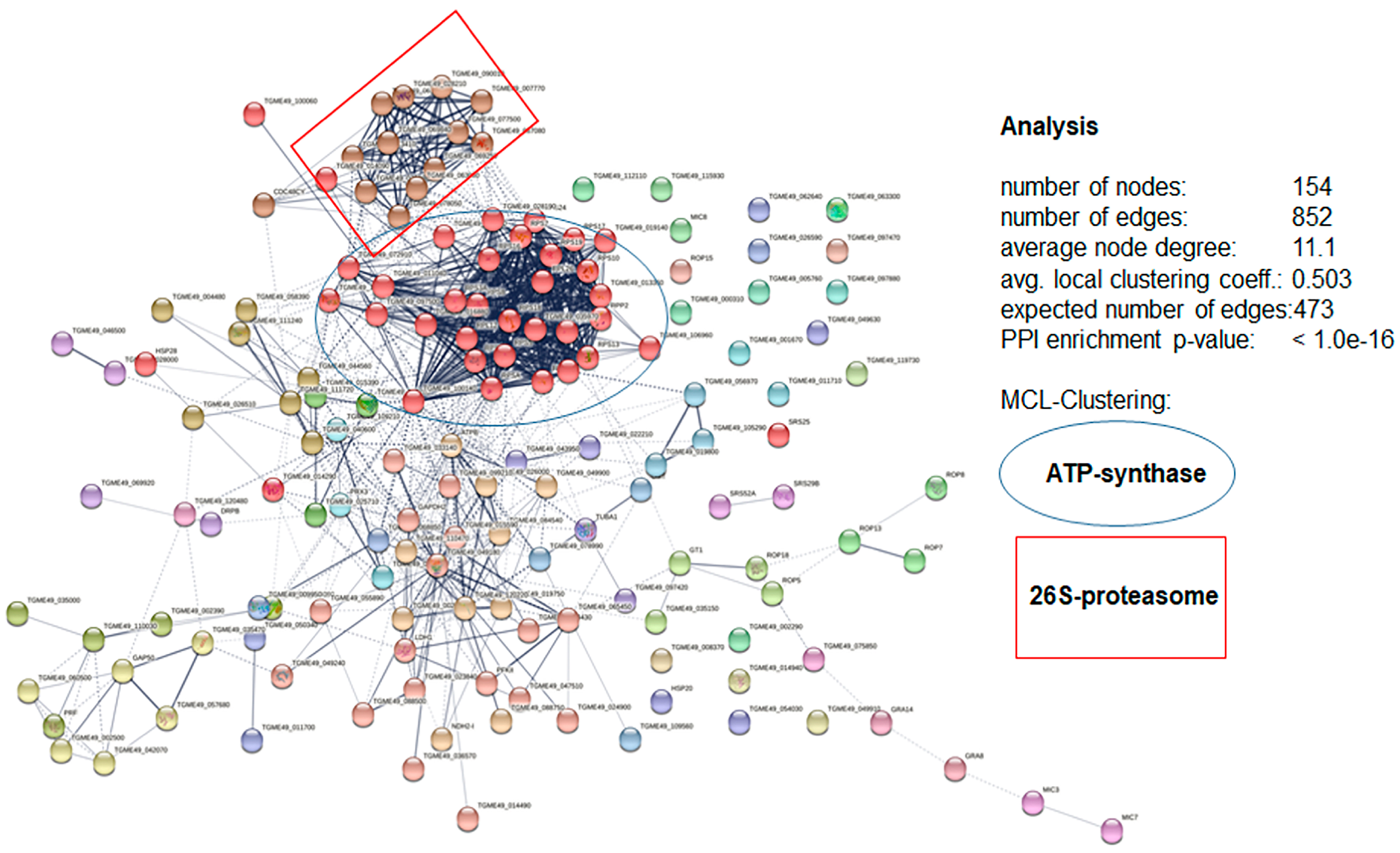
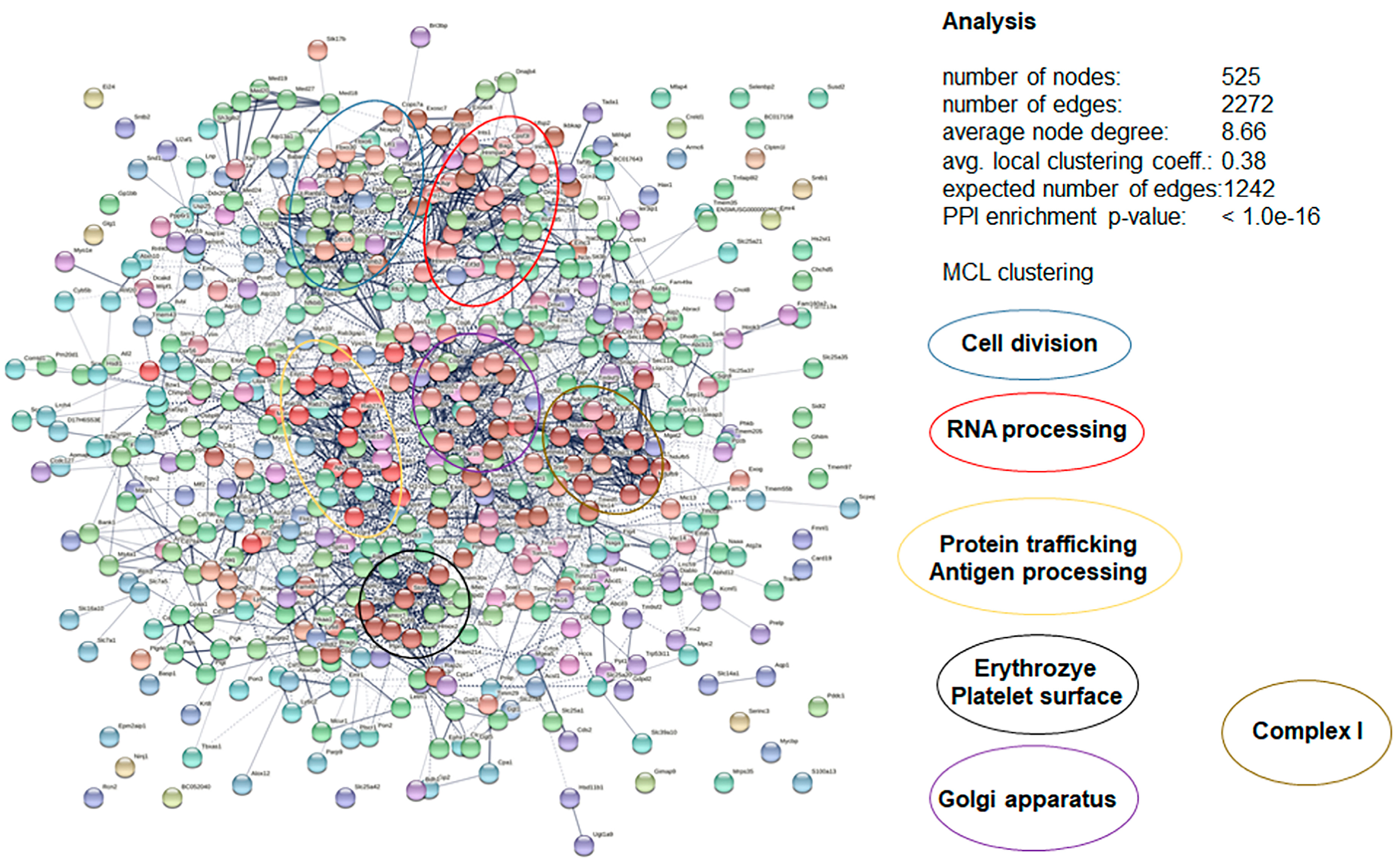
3.4. DAC Proteomes of Toxoplasma gondii Tachyzoites and Mouse Spleens
3.5. More Detailed Analysis of Proteome Data Reveals Putative Drug Targets of Peptide 6027 in T. gondii and Murine Spleen Cells
4. Discussion
5. Conclusions and Outlook
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dubey, J.P.; Hemphill, A.; Calero-Bernal, R.; Schares, G. Neosporosis in Animals; Taylor & Francis: Boca Raton, FL, USA, 2017; Volume xviii, 529p. [Google Scholar]
- Dubey, J.P. Toxoplasmosis of Animals and Humans, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2010; pp. 313–314. [Google Scholar]
- Adl, S.M.; Simpson, A.G.; Lane, C.E.; Lukes, J.; Bass, D.; Bowser, S.S.; Brown, M.W.; Burki, F.; Dunthorn, M.; Hampl, V.; et al. The revised classification of eukaryotes. J. Eukaryot. Microbiol. 2012, 59, 429–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, J.; Aguado-Martinez, A.; Balmer, V.; Maly, D.J.; Fan, E.; Ortega-Mora, L.M.; Ojo, K.K.; Van Voorhis, W.C.; Hemphill, A. Two novel calcium-dependent protein kinase 1 inhibitors interfere with vertical transmission in mice infected with Neospora caninum tachyzoites. Antimicrob. Agents Chemother. 2017, 61, e02324-16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basto, A.P.; Anghel, N.; Rubbiani, R.; Müller, J.; Stibal, D.; Giannini, F.; Suss-Fink, G.; Balmer, V.; Gasser, G.; Furrer, J.; et al. Targeting of the mitochondrion by dinuclear thiolato-bridged arene ruthenium complexes in cancer cells and in the apicomplexan parasite Neospora caninum. Metallomics 2019, 11, 462–474. [Google Scholar] [CrossRef] [Green Version]
- Anghel, N.; Winzer, P.A.; Imhof, D.; Müller, J.; Langa, X.; Rieder, J.; Barrett, L.K.; Vidadala, R.S.R.; Huang, W.; Choi, R.; et al. Comparative assessment of the effects of bumped kinase inhibitors on early zebrafish embryo development and pregnancy in mice. Int. J. Antimicrob. Agents 2020, 56, 106099. [Google Scholar] [CrossRef]
- Müller, J.; Hemphill, A. New approaches for the identification of drug targets in protozoan parasites. Int. Rev. Cell Mol. Biol. 2013, 301, 359–401. [Google Scholar] [CrossRef] [PubMed]
- Russell, W.M.S.; Burch, R.L. The Principles of Humane Experimental Technique; Methuen: London, UK, 1959; p. 238. [Google Scholar]
- Paunescu, E.; Boubaker, G.; Desiatkina, O.; Anghel, N.; Amdouni, Y.; Hemphill, A.; Furrer, J. The quest of the best—A SAR study of trithiolato-bridged dinuclear Ruthenium(II)-Arene compounds presenting antiparasitic properties. Eur. J. Med. Chem. 2021, 222, 113610. [Google Scholar] [CrossRef]
- Müller, J.; Naguleswaran, A.; Müller, N.; Hemphill, A. Neospora caninum: Functional inhibition of protein disulfide isomerase by the broad-spectrum anti-parasitic drug nitazoxanide and other thiazolides. Exp. Parasitol. 2008, 118, 80–88. [Google Scholar] [CrossRef]
- Müller, J.; Wastling, J.; Sanderson, S.; Müller, N.; Hemphill, A. A novel Giardia lamblia nitroreductase, GlNR1, interacts with nitazoxanide and other thiazolides. Antimicrob. Agents Chemother. 2007, 51, 1979–1986. [Google Scholar] [CrossRef] [Green Version]
- Müller, J.; Sidler, D.; Nachbur, U.; Wastling, J.; Brunner, T.; Hemphill, A. Thiazolides inhibit growth and induce glutathione-S-transferase Pi (GSTP1)-dependent cell death in human colon cancer cells. Int. J. Cancer 2008, 123, 1797–1806. [Google Scholar] [CrossRef]
- Müller, J.; Hemphill, A. Identification of a host cell target for the thiazolide class of broad-spectrum anti-parasitic drugs. Exp. Parasitol. 2011, 128, 145–150. [Google Scholar] [CrossRef]
- Anghel, N.; Müller, J.; Serricchio, M.; Jelk, J.; Butikofer, P.; Boubaker, G.; Imhof, D.; Ramseier, J.; Desiatkina, O.; Paunescu, E.; et al. Cellular and molecular targets of nucleotide-tagged trithiolato-bridged arene ruthenium complexes in the protozoan parasites Toxoplasma gondii and Trypanosoma brucei. Int. J. Mol. Sci. 2021, 22, 10787. [Google Scholar] [CrossRef] [PubMed]
- Müller, J.; Anghel, N.; Imhof, D.; Hänggeli, K.; Uldry, A.C.; Braga-Lagache, S.; Heller, M.; Ojo, K.K.; Ortega-Mora, L.M.; Van Voorhis, W.C.; et al. Common molecular targets of a quinoline based bumped kinase inhibitor in Neospora caninum and Danio rerio. Int. J. Mol. Sci. 2022, 23, 2381. [Google Scholar] [CrossRef] [PubMed]
- Arai, T.; Mikami, Y.; Fukushima, K.; Utsumi, T.; Yazawa, K. A new antibiotic, leucinostatin, derived from Penicillium lilacinum. J. Antibiot. 1973, 26, 157–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brand, M.; Wang, L.; Agnello, S.; Gazzola, S.; Gall, F.M.; Raguz, L.; Kaiser, M.; Schmidt, R.S.; Ritschl, A.; Jelk, J.; et al. Antiprotozoal structure-activity relationships of synthetic leucinostatin derivatives and elucidation of their mode of action. Angew. Chem. Int. Ed. Engl. 2021, 60, 15613–15621. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, S.; Müller, J.; Suana, A.; Hemphill, A. Vaccination with microneme protein NcMIC4 increases mortality in mice inoculated with Neospora caninum. J. Parasitol. 2007, 93, 1046–1055. [Google Scholar] [CrossRef]
- Sanchez-Sanchez, R.; Ferre, I.; Regidor-Cerrillo, J.; Gutierrez-Exposito, D.; Ferrer, L.M.; Arteche-Villasol, N.; Moreno-Gonzalo, J.; Muller, J.; Aguado-Martinez, A.; Perez, V.; et al. Virulence in mice of a Toxoplasma gondii type II isolate does not correlate with the outcome of experimental infection in pregnant sheep. Front. Cell. Infect. Microbiol. 2018, 8, 436. [Google Scholar] [CrossRef] [Green Version]
- Costa, J.M.; Pautas, C.; Ernault, P.; Foulet, F.; Cordonnier, C.; Bretagne, S. Real-time PCR for diagnosis and follow-up of Toxoplasma reactivation after allogeneic stem cell transplantation using fluorescence resonance energy transfer hybridization probes. J. Clin. Microbiol. 2000, 38, 2929–2932. [Google Scholar] [CrossRef] [Green Version]
- Imhof, D.; Anghel, N.; Winzer, P.; Balmer, V.; Ramseier, J.; Hanggeli, K.; Choi, R.; Hulverson, M.A.; Whitman, G.R.; Arnold, S.L.M.; et al. In vitro activity, safety and in vivo efficacy of the novel bumped kinase inhibitor BKI-1748 in non-pregnant and pregnant mice experimentally infected with Neospora caninum tachyzoites and Toxoplasma gondii oocysts. Int. J. Parasitol. Drugs Drug Resist. 2021, 16, 90–101. [Google Scholar] [CrossRef]
- Aguado-Martinez, A.; Basto, A.P.; Müller, J.; Balmer, V.; Manser, V.; Leitao, A.; Hemphill, A. N-terminal fusion of a toll-like receptor 2-ligand to a Neospora caninum chimeric antigen efficiently modifies the properties of the specific immune response. Parasitology 2016, 143, 606–616. [Google Scholar] [CrossRef]
- Müller, J.; Heller, M.; Uldry, A.C.; Braga, S.; Müller, N. Nitroreductase activites in Giardia lamblia: ORF 17150 encodes a quinone reductase with nitroreductase activity. Pathogens 2021, 10, 129. [Google Scholar] [CrossRef]
- Uldry, A.C.; Maciel-Dominguez, A.; Jornod, M.; Buchs, N.; Braga-Lagache, S.; Brodard, J.; Jankovic, J.; Bonadies, N.; Heller, M. Effect of sample transportation on the proteome of human circulating blood extracellular vesicles. Int. J. Mol. Sci. 2022, 23, 4515. [Google Scholar] [CrossRef]
- Esposito, M.; Stettler, R.; Moores, S.L.; Pidathala, C.; Müller, N.; Stachulski, A.; Berry, N.G.; Rossignol, J.F.; Hemphill, A. In vitro efficacies of nitazoxanide and other thiazolides against Neospora caninum tachyzoites reveal antiparasitic activity independent of the nitro group. Antimicrob. Agents Chemother. 2005, 49, 3715–3723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nishiyama, S.; Pradipta, A.; Ma, J.S.; Sasai, M.; Yamamoto, M. T cell-derived interferon-gamma is required for host defense to Toxoplasma gondii. Parasitol. Int. 2020, 75, 102049. [Google Scholar] [CrossRef]
- Ishiguro, K.; Arai, T. Action of the peptide antibiotic leucinostatin. Antimicrob. Agents Chemother. 1976, 9, 893–898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, Y.; Suzuki, M.; Fukushima, K.; Arai, T. Structure of leucinostatin B, an uncoupler on mitochondria. J. Antibiot. 1983, 36, 1084–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csermely, P.; Radics, L.; Rossi, C.; Szamel, M.; Ricci, M.; Mihaly, K.; Somogyi, J. The nonapeptide leucinostatin A acts as a weak ionophore and as an immunosuppressant on T lymphocytes. Biochim. Biophys. Acta 1994, 1221, 125–132. [Google Scholar] [CrossRef]
- Tsukihara, T.; Aoyama, H.; Yamashita, E.; Tomizaki, T.; Yamaguchi, H.; Shinzawa-Itoh, K.; Nakashima, R.; Yaono, R.; Yoshikawa, S. The whole structure of the 13-subunit oxidized cytochrome c oxidase at 2.8 A. Science 1996, 272, 1136–1144. [Google Scholar] [CrossRef]
- Crofts, A.R. The cytochrome bc1 complex: Function in the context of structure. Annu. Rev. Physiol. 2004, 66, 689–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seidi, A.; Muellner-Wong, L.S.; Rajendran, E.; Tjhin, E.T.; Dagley, L.F.; Aw, V.Y.; Faou, P.; Webb, A.I.; Tonkin, C.J.; van Dooren, G.G. Elucidating the mitochondrial proteome of Toxoplasma gondii reveals the presence of a divergent cytochrome c oxidase. Elife 2018, 7, e38131. [Google Scholar] [CrossRef] [PubMed]
- Mallo, N.; Ovciarikova, J.; Martins-Duarte, E.S.; Baehr, S.C.; Biddau, M.; Wilde, M.L.; Uboldi, A.D.; Lemgruber, L.; Tonkin, C.J.; Wideman, J.G.; et al. Depletion of a Toxoplasma porin leads to defects in mitochondrial morphology and contacts with the endoplasmic reticulum. J. Cell Sci. 2021, 134, jcs255299. [Google Scholar] [CrossRef]
- Shima, A.; Fukushima, K.; Arai, T.; Terada, H. Dual inhibitory effects of the peptide antibiotics leucinostatins on oxidative phosphorylation in mitochondria. Cell Struct. Funct. 1990, 15, 53–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandt, U. Energy converting NADH: Quinone oxidoreductase (complex I). Annu. Rev. Biochem. 2006, 75, 69–92. [Google Scholar] [CrossRef]
- Michelin, A.; Bittame, A.; Bordat, Y.; Travier, L.; Mercier, C.; Dubremetz, J.F.; Lebrun, M. GRA12, a Toxoplasma dense granule protein associated with the intravacuolar membranous nanotubular network. Int. J. Parasitol. 2009, 39, 299–306. [Google Scholar] [CrossRef] [PubMed]
- Fox, B.A.; Guevara, R.B.; Rommereim, L.M.; Falla, A.; Bellini, V.; Petre, G.; Rak, C.; Cantillana, V.; Dubremetz, J.F.; Cesbron-Delauw, M.F.; et al. Toxoplasma gondii parasitophorous vacuole membrane-associated dense granule proteins orchestrate chronic infection and GRA12 underpins resistance to host gamma interferon. MBio 2019, 10, e00589-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercier, C.; Cesbron-Delauw, M.F. Toxoplasma secretory granules: One population or more? Trends Parasitol. 2015, 31, 604. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Martin, R.D.; Mercier, C.; Gomez de Leon, C.T.; Gonzalez, R.M.; Pozos, S.G.; Rios-Castro, E.; Garcia, R.A.; Fox, B.A.; Bzik, D.J.; Flores, R.M. The dense granule protein 8 (GRA8) is a component of the sub-pellicular cytoskeleton in Toxoplasma gondii. Parasitol. Res. 2019, 118, 1899–1918. [Google Scholar] [CrossRef] [PubMed]
- Osborne, A.R.; Rapoport, T.A.; van den Berg, B. Protein translocation by the Sec61/SecY channel. Annu. Rev. Cell Dev. Biol. 2005, 21, 529–550. [Google Scholar] [CrossRef]
- Liaci, A.M.; Steigenberger, B.; Telles de Souza, P.C.; Tamara, S.; Grollers-Mulderij, M.; Ogrissek, P.; Marrink, S.J.; Scheltema, R.A.; Forster, F. Structure of the human signal peptidase complex reveals the determinants for signal peptide cleavage. Mol. Cell 2021, 81, 3934–3948.e3911. [Google Scholar] [CrossRef]
- Zhao, X.; Yang, H.; Liu, W.; Duan, X.; Shang, W.; Xia, D.; Tong, C. Sec22 regulates endoplasmic reticulum morphology but not autophagy and is required for eye development in Drosophila. J. Biol. Chem. 2015, 290, 7943–7951. [Google Scholar] [CrossRef] [Green Version]
- Shaw, M.K.; He, C.Y.; Roos, D.S.; Tilney, L.G. Proteasome inhibitors block intracellular growth and replication of Toxoplasma gondii. Parasitology 2000, 121 Pt 1, 35–47. [Google Scholar] [CrossRef]
- Kozlova, V.; Ledererova, A.; Ladungova, A.; Peschelova, H.; Janovska, P.; Slusarczyk, A.; Domagala, J.; Kopcil, P.; Vakulova, V.; Oppelt, J.; et al. CD20 is dispensable for B-cell receptor signaling but is required for proper actin polymerization, adhesion and migration of malignant B cells. PLoS ONE 2020, 15, e0229170. [Google Scholar] [CrossRef]
- Novi, G.; Bovis, F.; Fabbri, S.; Tazza, F.; Gazzola, P.; Maietta, I.; Curro, D.; Bruschi, N.; Roccatagliata, L.; Boffa, G.; et al. Tailoring B cell depletion therapy in MS according to memory B cell monitoring. Neurol. Neuroimmunol. Neuroinflamm. 2020, 7, e845. [Google Scholar] [CrossRef] [PubMed]
- Menard, L.C.; Minns, L.A.; Darche, S.; Mielcarz, D.W.; Foureau, D.M.; Roos, D.; Dzierszinski, F.; Kasper, L.H.; Buzoni-Gatel, D. B cells amplify IFN-gamma production by T cells via a TNF-alpha-mediated mechanism. J. Immunol. 2007, 179, 4857–4866. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ORF | Annotation | Relative Abundance (×103) (%) | |
|---|---|---|---|
| TGRH88_008910 | Sec61beta family protein | 33.2 | 9.0 |
| TGRH88_013490 | Dense granule protein GRA12 | 22.8 | 6.2 |
| TGRH88_067710 | Eukaryotic porin protein | 20.0 | 5.4 |
| TGRH88_002590 | Unspecified product (TgApiCox30) | 14.5 | 3.9 |
| TGRH88_000580 | Unspecified product (TgApiCox18) | 11.7 | 3.2 |
| TGRH88_067940 | 14-3-3 protein | 11.3 | 3.1 |
| TGRH88_061860 | Unspecified product (TgApiCox19) | 7.5 | 2.0 |
| TGRH88_004320 | Zinc finger CDGSH-type domain-containing protein | 6.9 | 1.9 |
| TGRH88_006890 | Putative ubiquinol cytochrome c oxidoreductase | 6.8 | 1.8 |
| TGRH88_018120 | Unspecified product (TgApiCox26) | 6.6 | 1.8 |
| TGRH88_074430 | Unspecified product (TgApiCox35) | 6.5 | 1.8 |
| TGRH88_067370 | Unspecified product | 6.5 | 1.8 |
| TGRH88_081780 | Putative calmodulin | 6.2 | 1.7 |
| TGRH88_040180 | Facilitative glucose transporter GT1 | 6.0 | 1.6 |
| TGRH88_051090 | Chaperonin protein BiP | 5.9 | 1.6 |
| TGRH88_003600 | Dense granule protein GRA8 | 5.8 | 1.6 |
| TGRH88_050010 | Putative cytochrome C oxidase subunit IIb (TgCox2b) | 5.2 | 1.4 |
| TGRH88_022800 | Ribosomal protein RPS17 | 4.9 | 1.3 |
| TGRH88_012110 | Unspecified product (TgApiCox16) | 4.6 | 1.3 |
| TGRH88_068380 | Cg8 family protein (TgApiCox23) | 4.6 | 1.2 |
| IDs | Annotation | Relative Abundance (rAbu) (%) | |
|---|---|---|---|
| Q9CQP3 | Coiled-coil-helix-coiled-coil-helix domain-containing protein 5 | 1290 | 2.1 |
| Q9D958 | Signal peptidase complex subunit 1 | 1231 | 2.0 |
| O08547 | Vesicle-trafficking protein SEC22b | 1186 | 1.9 |
| Q8R5J9 | PRA1 family protein 3 | 1162 | 1.9 |
| P17665 | Cytochrome c oxidase subunit 7C, mitochondrial | 1143 | 1.8 |
| P35459 | Lymphocyte antigen 6D | 1141 | 1.8 |
| Q08857 | Platelet glycoprotein 4 | 1078 | 1.7 |
| Q9D8V7 | Signal peptidase complex catalytic subunit SEC11C | 1023 | 1.6 |
| P20491 | High affinity immunoglobulin epsilon receptor subunit gamma | 1004 | 1.6 |
| Q8BXV2 | BRI3-binding protein | 945 | 1.5 |
| P19437 | B-lymphocyte antigen CD20 | 935 | 1.5 |
| P97370 | Sodium/potassium-transporting ATPase subunit beta-3 | 907 | 1.4 |
| Q9DCF9-2 | Isoform 2 of Translocon-associated protein subunit gamma | 835 | 1.3 |
| P61166 | Transmembrane protein 258 | 821 | 1.3 |
| Q9JKW0 | ADP-ribosylation factor-like protein 6-interacting protein | 821 | 1.3 |
| Q9R0P6 | Signal peptidase complex catalytic subunit SEC11A | 819 | 1.3 |
| P54116 | Stomatin | 680 | 1.1 |
| Q9D3P8 | Plasminogen receptor (KT) | 601 | 1.0 |
| Q9R0Q3 | Transmembrane emp24 domain-containing protein 2 | 600 | 1.0 |
| P46638 | Ras-related protein Rab-11B | 573 | 0.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Müller, J.; Boubaker, G.; Imhof, D.; Hänggeli, K.; Haudenschild, N.; Uldry, A.-C.; Braga-Lagache, S.; Heller, M.; Ortega-Mora, L.-M.; Hemphill, A. Differential Affinity Chromatography Coupled to Mass Spectrometry: A Suitable Tool to Identify Common Binding Proteins of a Broad-Range Antimicrobial Peptide Derived from Leucinostatin. Biomedicines 2022, 10, 2675. https://doi.org/10.3390/biomedicines10112675
Müller J, Boubaker G, Imhof D, Hänggeli K, Haudenschild N, Uldry A-C, Braga-Lagache S, Heller M, Ortega-Mora L-M, Hemphill A. Differential Affinity Chromatography Coupled to Mass Spectrometry: A Suitable Tool to Identify Common Binding Proteins of a Broad-Range Antimicrobial Peptide Derived from Leucinostatin. Biomedicines. 2022; 10(11):2675. https://doi.org/10.3390/biomedicines10112675
Chicago/Turabian StyleMüller, Joachim, Ghalia Boubaker, Dennis Imhof, Kai Hänggeli, Noé Haudenschild, Anne-Christine Uldry, Sophie Braga-Lagache, Manfred Heller, Luis-Miguel Ortega-Mora, and Andrew Hemphill. 2022. "Differential Affinity Chromatography Coupled to Mass Spectrometry: A Suitable Tool to Identify Common Binding Proteins of a Broad-Range Antimicrobial Peptide Derived from Leucinostatin" Biomedicines 10, no. 11: 2675. https://doi.org/10.3390/biomedicines10112675