

The Transcriptional Regulator Prdm1 Is Essential for the Early Development of the Sensory Whisker Follicle and Is Linked to the Beta-Catenin First Dermal Signal
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mouse Strains
2.2. In Situ Hybridization
2.3. In Vivo Lineage Tracing
2.4. Proliferation Experiments
2.5. Histology and Immunostaining
2.6. Imaging
2.7. RT–PCR
2.8. Quantitative PCR
2.9. Interspecies Sequence Comparison
2.10. 4C-Seq
2.11. Statistical Analysis
2.12. CUT&Tag
3. Results
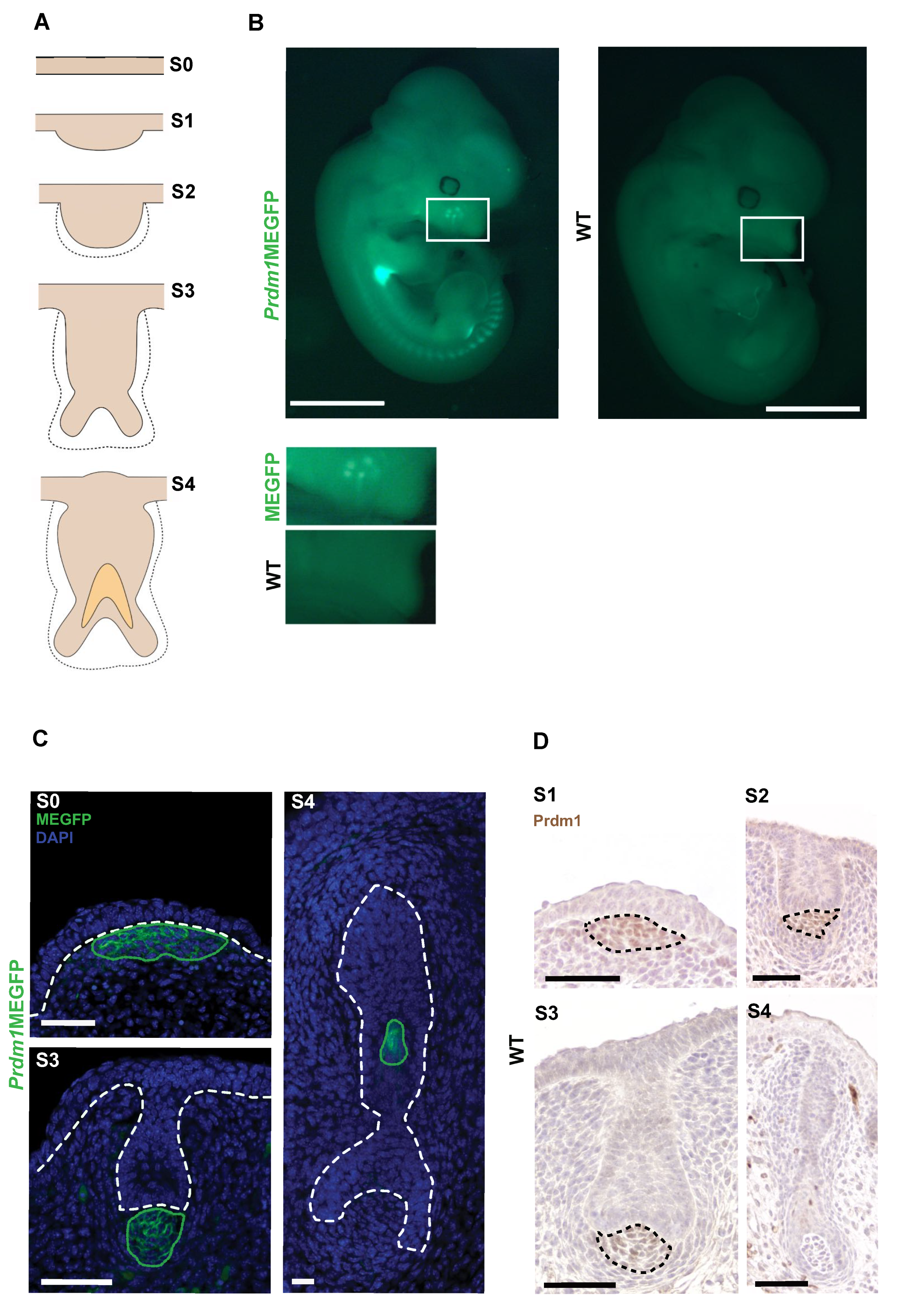
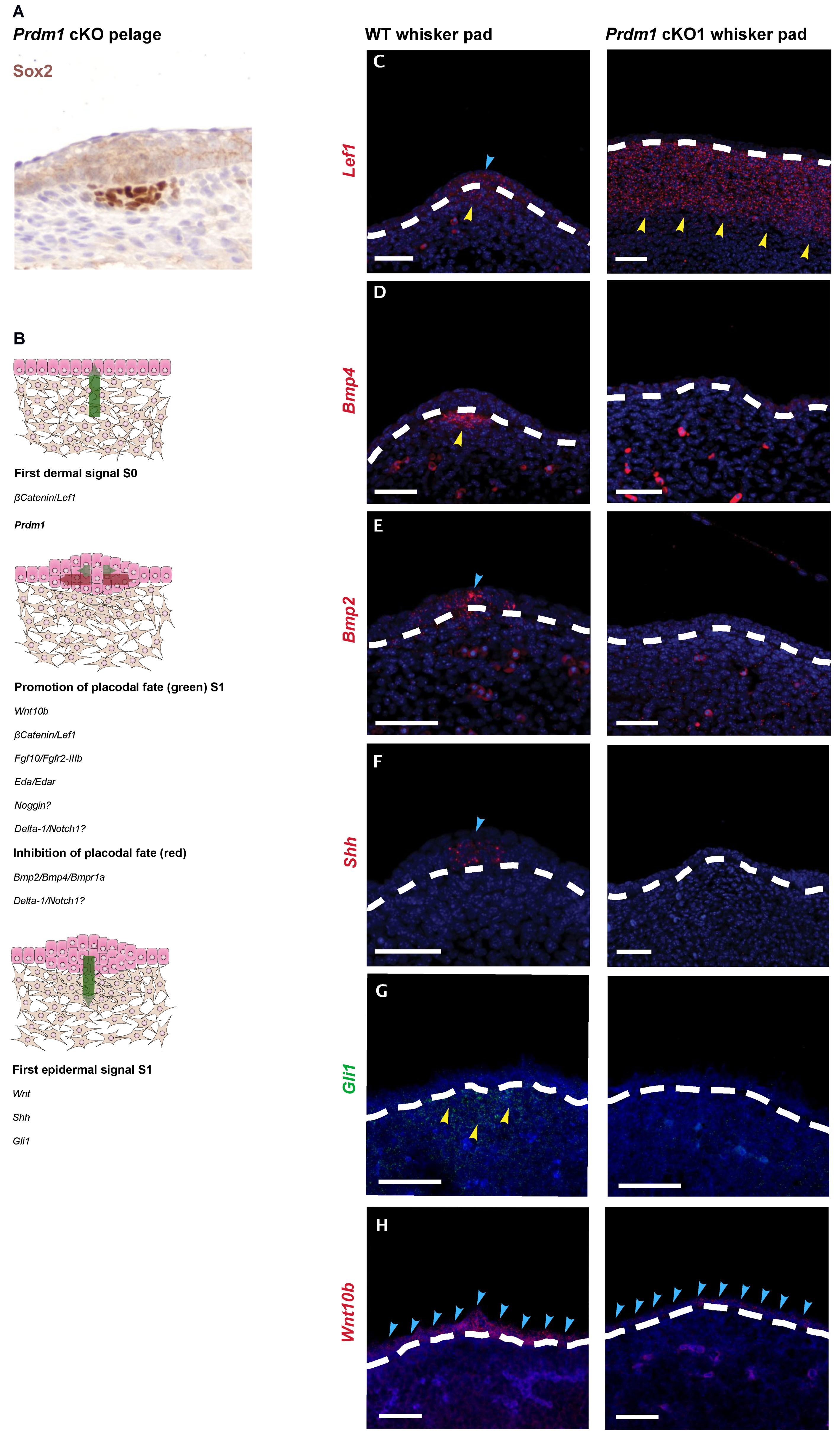
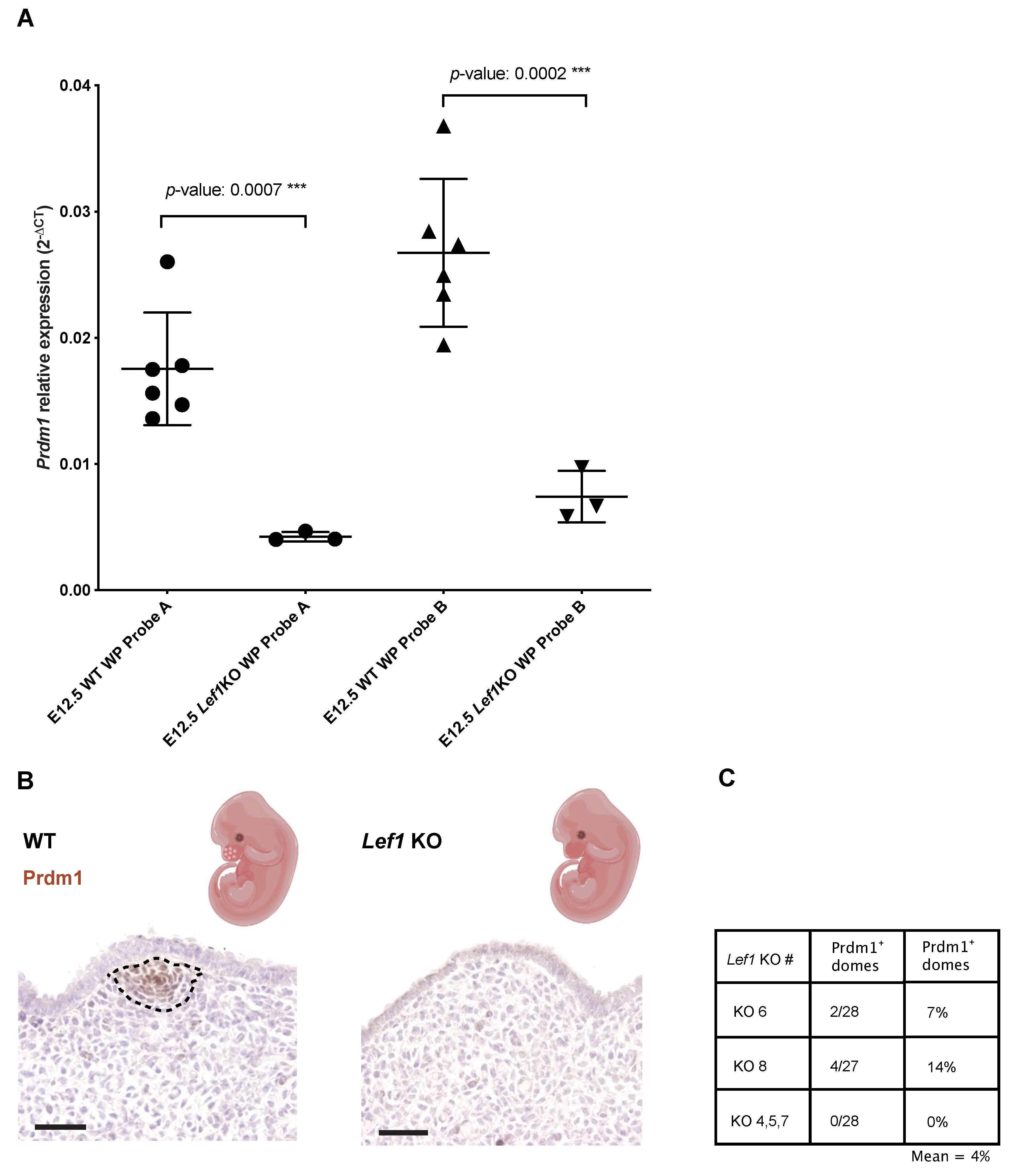
3.1. Prdm1 Is an Essential Gene for Whisker Follicle Development

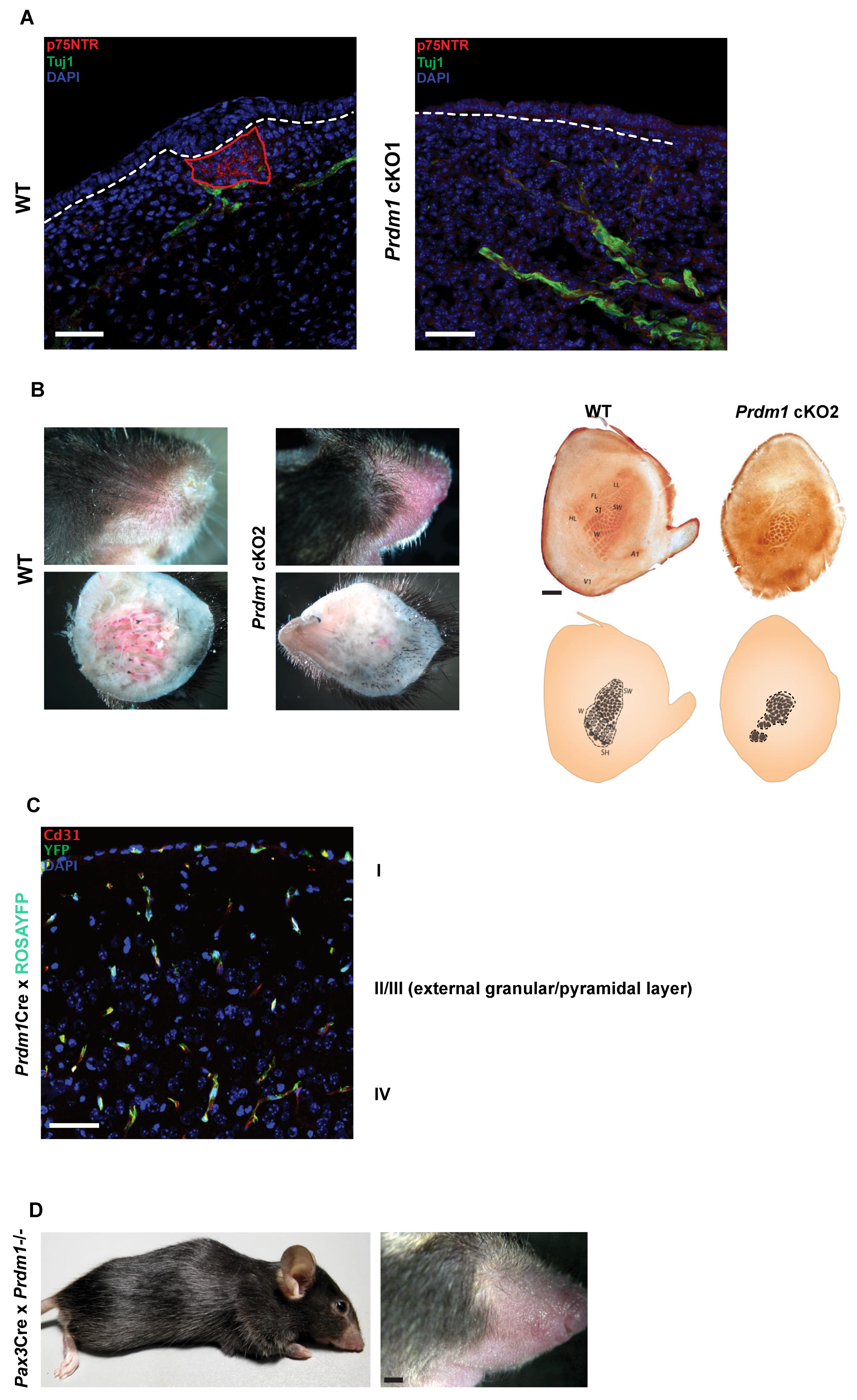
3.2. The Whisker Inducing Mesenchyme Contributes to Several Lineages of the Adult Whisker
3.3. Prdm1 Genetic Ablation Leads to the Reorganization of the Rodent Barrel Cortex
3.4. Prdm1 Knockouts Lead to Disrupted Trigeminal Nerve Wiring and Major Reorganization of the Barrel Cortex
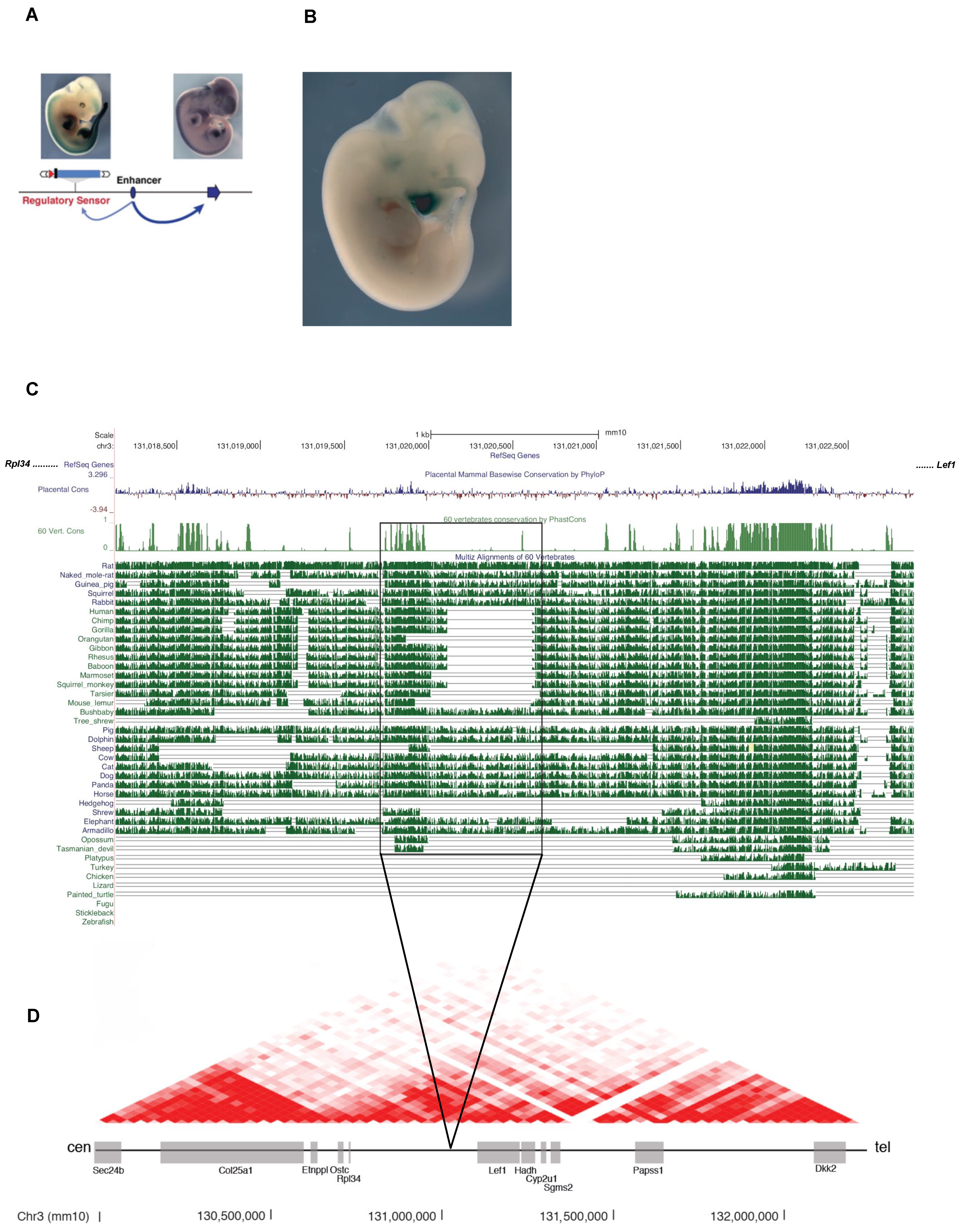
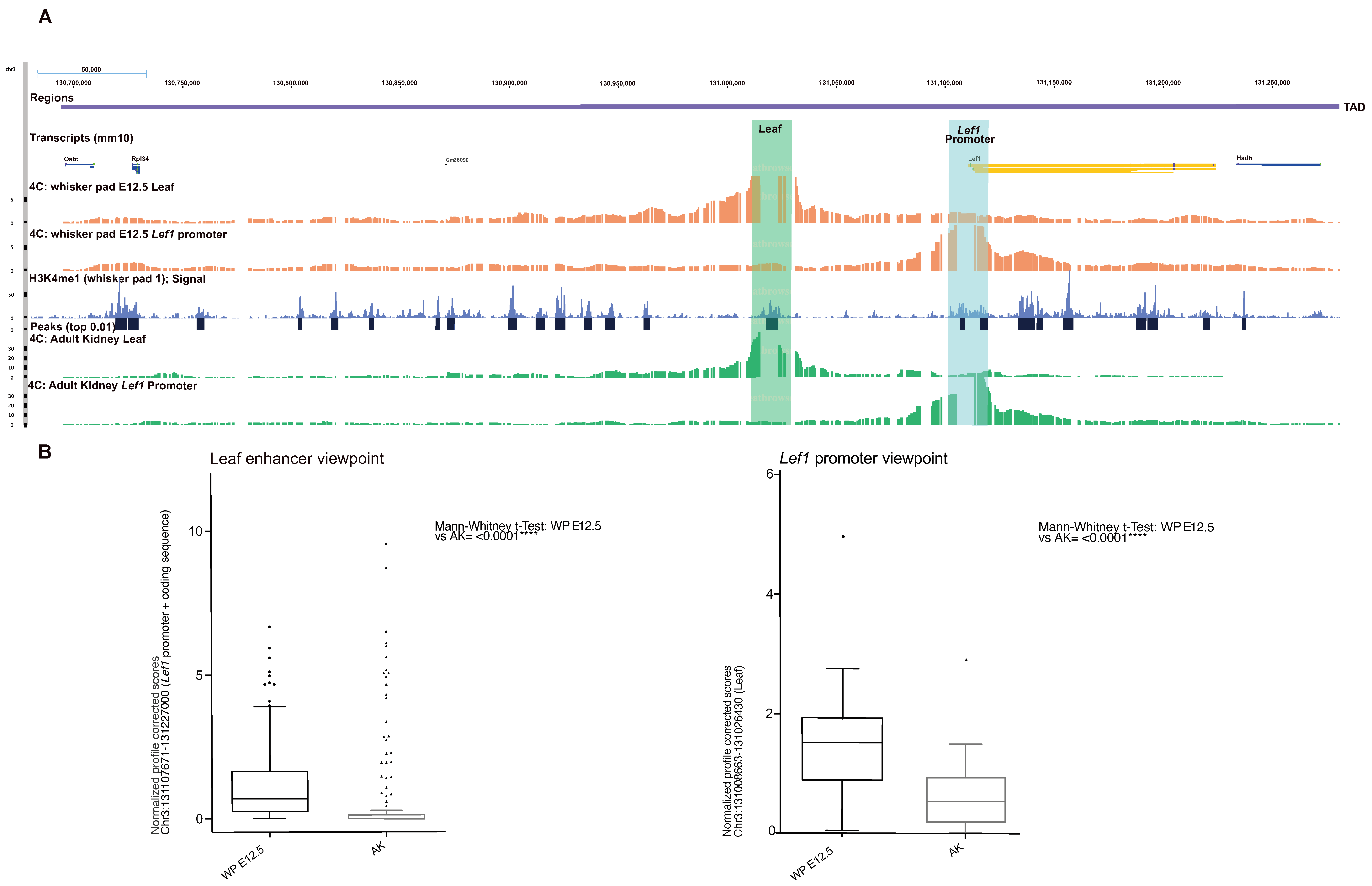
3.5. Identification of Regulatory Elements Lost during Evolution in Whisker Morphogenesis

3.6. Expression of an Ar Regulatory Enhancer Specifically Lost in Humans in the Early Steps of Whisker Development
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oshima, H.; Rochat, A.; Kedzia, C.; Kobayashi, K.; Barrandon, Y. Morphogenesis and renewal of hair follicles from adult multipotent stem cells. Cell 2001, 104, 233–245. [Google Scholar] [CrossRef] [Green Version]
- Claudinot, S.; Nicolas, M.; Oshima, H.; Rochat, A.; Barrandon, Y. Long-term renewal of hair follicles from clonogenic multipotent stem cells. Proc. Natl. Acad. Sci. USA 2005, 102, 14677–14682. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petersen, C.C.H. The functional organization of the barrel cortex. Neuron 2007, 56, 339–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brecht, M. Barrel cortex and whisker-mediated behaviors. Curr. Opin. Neurobiol. 2007, 17, 408–416. [Google Scholar] [CrossRef] [PubMed]
- Diamond, M.E.; von Heimendahl, M.; Knutsen, P.M.; Kleinfeld, D.; Ahissar, E. “Where” and “what” in the whisker sensorimotor system. Nat. Rev. Neurosci. 2008, 9, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Van Horn, R.N. Vibrissae structure in the rhesus monkey. Folia Primatol. 1970, 13, 241–285. [Google Scholar] [CrossRef] [PubMed]
- Tamatsu, Y.; Tsukahara, K.; Hotta, M.; Shimada, K. Vestiges of vibrissal capsular muscles exist in the human upper lip. Clin. Anat. 2007, 20, 628–631. [Google Scholar] [CrossRef]
- Woolsey, T.A.; Van der Loos, H. The structural organization of layer IV in the somatosensory region (SI) of mouse cerebral cortex. The description of a cortical field composed of discrete cytoarchitectonic units. Brain Res. 1970, 17, 205–242. [Google Scholar] [CrossRef]
- Erzurumlu, R.S.; Gaspar, P. Development and critical period plasticity of the barrel cortex. Eur. J. Neurosci. 2012, 35, 1540–1553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrés, F.L.; Van der Loos, H. From sensory periphery to cortex: The architecture of the barrelfield as modified by various early manipulations of the mouse whiskerpad. Anat. Embryol. 1985, 172, 11–20. [Google Scholar] [CrossRef] [PubMed]
- Keller, A.D.; Maniatis, T. Identification and characterization of a novel repressor of beta-interferon gene expression. Genes Dev. 1991, 5, 868–879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Angelin-Duclos, C.; Cattoretti, G.; Lin, K.I.; Calame, K. Commitment of B lymphocytes to a plasma cell fate is associated with Blimp-1 expression in vivo. J. Immunol. 2000, 165, 5462–5471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shaffer, A.L.; Lin, K.I.; Kuo, T.C.; Yu, X.; Hurt, E.M.; Rosenwald, A.; Giltnane, J.M.; Yang, L.; Zhao, H.; Calame, K.; et al. Blimp-1 orchestrates plasma cell differentiation by extinguishing the mature B cell gene expression program. Immunity 2002, 17, 51–62. [Google Scholar] [CrossRef] [Green Version]
- Shapiro-Shelef, M.; Lin, K.-I.; McHeyzer-Williams, L.J.; Liao, J.; McHeyzer-Williams, M.G.; Calame, K. Blimp-1 is required for the formation of immunoglobulin secreting plasma cells and pre-plasma memory B cells. Immunity 2003, 19, 607–620. [Google Scholar] [CrossRef] [Green Version]
- Martins, G.A.; Cimmino, L.; Shapiro-Shelef, M.; Szabolcs, M.; Herron, A.; Magnusdottir, E.; Calame, K. Transcriptional repressor Blimp-1 regulates T cell homeostasis and function. Nat. Immunol. 2006, 7, 457–465. [Google Scholar] [CrossRef] [PubMed]
- Vincent, S.D.; Dunn, N.R.; Sciammas, R.; Shapiro-Shalef, M.; Davis, M.M.; Calame, K.; Bikoff, E.K.; Robertson, E.J. The zinc finger transcriptional repressor Blimp1/Prdm1 is dispensable for early axis formation but is required for specification of primordial germ cells in the mouse. Development 2005, 132, 1315–1325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohinata, Y.; Payer, B.; O’Carroll, D.; Ancelin, K.; Ono, Y.; Sano, M.; Barton, S.C.; Obukhanych, T.; Nussenzweig, M.; Tarakhovsky, A.; et al. Blimp1 is a critical determinant of the germ cell lineage in mice. Nature 2005, 436, 207–213. [Google Scholar] [CrossRef]
- Magnúsdóttir, E.; Kalachikov, S.; Mizukoshi, K.; Savitsky, D.; Ishida-Yamamoto, A.; Panteleyev, A.A.; Calame, K. Epidermal terminal differentiation depends on B lymphocyte-induced maturation protein-1. Proc. Natl. Acad. Sci. USA 2007, 104, 14988–14993. [Google Scholar] [CrossRef] [Green Version]
- Horsley, V.; O’Carroll, D.; Tooze, R.; Ohinata, Y.; Saitou, M.; Obukhanych, T.; Nussenzweig, M.; Tarakhovsky, A.; Fuchs, E. Blimp1 defines a progenitor population that governs cellular input to the sebaceous gland. Cell 2006, 126, 597–609. [Google Scholar] [CrossRef] [Green Version]
- Robertson, E.J.; Charatsi, I.; Joyner, C.J.; Koonce, C.H.; Morgan, M.; Islam, A.; Paterson, C.; Lejsek, E.; Arnold, S.J.; Kallies, A.; et al. Blimp1 regulates development of the posterior forelimb, caudal pharyngeal arches, heart and sensory vibrissae in mice. Development 2007, 134, 4335–4345. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Dixon, J.R.; Selvaraj, S.; Yue, F.; Kim, A.; Li, Y.; Shen, Y.; Hu, M.; Liu, J.S.; Ren, B. Topological domains in mammalian genomes identified by analysis of chromatin interactions. Nature 2012, 485, 376–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Song, F.; Zhang, B.; Zhang, L.; Xu, J.; Kuang, D.; Li, D.; Choudhary, M.N.K.; Li, Y.; Hu, M.; et al. The 3D Genome Browser: A web-based browser for visualizing 3D genome organization and long-range chromatin interactions. Genome Biol. 2018, 19, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- David, F.P.A.; Delafontaine, J.; Carat, S.; Ross, F.J.; Lefebvre, G.; Jarosz, Y.; Sinclair, L.; Noordermeer, D.; Rougemont, J.; Leleu, M. HTSstation: A web application and open-access libraries for high-throughput sequencing data analysis. PLoS ONE 2014, 9, e85879. [Google Scholar] [CrossRef]
- Noordermeer, D.; Leleu, M.; Splinter, E.; Rougemont, J.; De Laat, W.; Duboule, D. The dynamic architecture of Hox gene clusters. Science 2011, 334, 222–225. [Google Scholar] [CrossRef] [Green Version]
- Tolhuis, B.; Blom, M.; Kerkhoven, R.M.; Pagie, L.; Teunissen, H.; Nieuwland, M.; Simonis, M.; de Laat, W.; van Lohuizen, M.; van Steensel, B. Interactions among Polycomb domains are guided by chromosome architecture. PLoS Genet. 2011, 7, e1001343. [Google Scholar] [CrossRef] [Green Version]
- Lieberman-Aiden, E.; van Berkum, N.L.; Williams, L.; Imakaev, M.; Ragoczy, T.; Telling, A.; Amit, I.; Lajoie, B.R.; Sabo, P.J.; Dorschner, M.O.; et al. Comprehensive mapping of long-range interactions reveals folding principles of the human genome. Science 2009, 326, 289–293. [Google Scholar] [CrossRef] [Green Version]
- Meers, M.P.; Tenenbaum, D.; Henikoff, S. Peak calling by Sparse Enrichment Analysis for CUTnRUN chromatin profiling. Epigenet. Chromatin. 2019, 12, 42. [Google Scholar] [CrossRef] [Green Version]
- Hardy, M.H. The secret life of the hair follicle. Trends Genet. 1992, 8, 55–61. [Google Scholar] [CrossRef]
- Hayashi, S.; Tenzen, T.; McMahon, A.P. Maternal inheritance of Cre activity in a Sox2Cre deleter strain. Genesis 2003, 37, 51–53. [Google Scholar] [CrossRef]
- Noramly, S.; Freeman, A.; Morgan, B.A. beta-catenin signaling can initiate feather bud development. Development 1999, 126, 3509–3521. [Google Scholar] [CrossRef] [PubMed]
- Van Genderen, C.; Okamura, R.M.; Fariñas, I.; Quo, R.G.; Parslow, T.G.; Bruhn, L.; Grosschedl, R. Development of several organs that require inductive epithelial-mesenchymal interactions is impaired in LEF-1-deficient mice. Genes Dev. 1994, 8, 2691–2703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kratochwil, K.; Dull, M.; Farinas, I.; Galceran, J.; Grosschedl, R. Lef1 expression is activated by BMP-4 and regulates inductive tissue interactions in tooth and hair development. Genes Dev. 1996, 10, 1382–1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGowan, K.M.; Coulombe, P.A. Onset of keratin 17 expression coincides with the definition of major epithelial lineages during skin development. J. Cell Biol. 1998, 143, 469–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millar, S.E. Molecular mechanisms regulating hair follicle development. J. Investig. Dermatol. 2002, 118, 216–225. [Google Scholar] [CrossRef] [PubMed]
- Madisen, L.; Zwingman, T.A.; Sunkin, S.M.; Oh, S.W.; Zariwala, H.A.; Gu, H.; Ng, L.L.; Palmiter, R.D.; Hawrylycz, M.J.; Jones, A.R.; et al. A robust and high-throughput Cre reporting and characterization system for the whole mouse brain. Nat. Neurosci. 2010, 13, 133–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, K.; Sarkar, A.; Yram, M.A.; Polo, J.M.; Bronson, R.; Sengupta, S.; Seandel, M.; Geijsen, N.; Hochedlinger, K. Sox2(+) adult stem and progenitor cells are important for tissue regeneration and survival of mice. Cell Stem. Cell 2011, 9, 317–329. [Google Scholar] [CrossRef] [Green Version]
- Danielian, P.S.; Muccino, D.; Rowitch, D.H.; Michael, S.K.; McMahon, A.P. Modification of gene activity in mouse embryos in utero by a tamoxifen-inducible form of Cre recombinase. Curr. Biol. 1998, 8, 1323–1326. [Google Scholar] [CrossRef] [Green Version]
- Lang, D.; Lu, M.M.; Huang, L.; Engleka, K.A.; Zhang, M.; Chu, E.Y.; Lipner, S.; Skoultchi, A.; Millar, S.E.; Epstein, J.A. Pax3 functions at a nodal point in melanocyte stem cell differentiation. Nature 2005, 433, 884–887. [Google Scholar] [CrossRef]
- Chen, C.-K.; Symmons, O.; Uslu, V.V.; Tsujimura, T.; Ruf, S.; Smedley, D.; Spitz, F. TRACER: A resource to study the regulatory architecture of the mouse genome. BMC Genom. 2013, 14, 215. [Google Scholar] [CrossRef]
- Darbellay, F.; Necsulea, A. Comparative Transcriptomics Analyses across Species, Organs, and Developmental Stages Reveal Functionally Constrained lncRNAs. Mol. Biol. Evol. 2020, 37, 240–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henikoff, S.; Henikoff, J.G.; Kaya-Okur, H.S.; Ahmad, K. Efficient chromatin accessibility mapping in situ by nucleosome-tethered tagmentation. eLife 2020, 9, e63274. [Google Scholar] [CrossRef] [PubMed]
- McLean, C.Y.; Reno, P.L.; Pollen, A.A.; Bassan, A.I.; Capellini, T.D.; Guenther, C.; Indjeian, V.B.; Lim, X.; Menke, D.B.; Schaar, B.T.; et al. Human-specific loss of regulatory DNA and the evolution of human-specific traits. Nature 2011, 471, 216–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reno, P.L.; McLean, C.Y.; Hines, J.E.; Capellini, T.D.; Bejerano, G.; Kingsley, D.M. A penile spine/vibrissa enhancer sequence is missing in modern and extinct humans but is retained in multiple primates with penile spines and sensory vibrissae. PLoS ONE 2013, 8, e84258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, C.E.; Paratore, C.; Dours-Zimmermann, M.T.; Rochat, A.; Pietri, T.; Suter, U.; Zimmermann, D.R.; Dufour, S.; Thiery, J.P.; Meijer, D.; et al. Neural crest-derived cells with stem cell features can be traced back to multiple lineages in the adult skin. J. Cell Biol. 2006, 175, 1005–1015. [Google Scholar] [CrossRef] [PubMed]
- Yeh, S.; Tsai, M.-Y.; Xu, Q.; Mu, X.-M.; Lardy, H.; Huang, K.-E.; Lin, H.; Yeh, S.-D.; Altuwaijri, S.; Zhou, X.; et al. Generation and characterization of androgen receptor knockout (ARKO) mice: An in vivo model for the study of androgen functions in selective tissues. Proc. Natl. Acad. Sci. USA 2002, 99, 13498–13503. [Google Scholar] [CrossRef] [Green Version]
- Telerman, S.B.; Rognoni, E.; Sequeira, I.; Pisco, A.O.; Lichtenberger, B.M.; Culley, O.J.; Viswanathan, P.; Driskell, R.R.; Watt, F.M. Dermal Blimp1 Acts Downstream of Epidermal TGFβ and Wnt/β-Catenin to Regulate Hair Follicle Formation and Growth. J. Investig. Dermatol. 2017, 137, 2270–2281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sellheyer, K.; Krahl, D. Blimp-1: A marker of terminal differentiation but not of sebocytic progenitor cells. J. Cutan. Pathol. 2010, 37, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Oliver, R.F. Whisker growth after removal of the dermal papilla and lengths of follicle in the hooded rat. J. Embryol. Exp. Morphol. 1966, 15, 331–347. [Google Scholar] [CrossRef] [PubMed]
- Jahoda, C.A.; Oliver, R.F. Histological studies of the effects of wounding vibrissa follicles in the hooded rat. J. Embryol. Exp. Morphol. 1984, 83, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Attanasio, C.; Nord, A.S.; Zhu, Y.; Blow, M.J.; Li, Z.; Liberton, D.K.; Morrison, H.; Plajzer-Frick, I.; Holt, A.; Hosseini, R.; et al. Fine tuning of craniofacial morphology by distant-acting enhancers. Science 2013, 342, 1241006. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain Name | Provider |
|---|---|
| B6.Cg-Tg(Prdm1-cre)1Masu/J | The Jackson Laboratory |
| B6.129-Prdm1tm1Clme/J | The Jackson Laboratory |
| Prdm1MEGFP | Mitinori Saitou, MTA |
| B6.Cg-Gt(ROSA)26Sortm3(CAG-EYFP)Hze/J | The Jackson Laboratory |
| Tg(Sox2-cre)1Amc | The Jackson Laboratory |
| B6;129S-Sox2tm1(cre/ERT2)Hoch/J | The Jackson Laboratory |
| STOCK Tg(Wnt1-cre)11Rth Tg(Wnt1-GAL4)11Rth/J | The Jackson Laboratory |
| Lef1tm1Rug | Rudolf Grosschedl and Werner Held, MTA |
| Primer Name | Primer Sequences |
|---|---|
| EGFP * | Primer Fw: CCTACGGCGTGCAGTGCTTCAGC |
| Primer Rv: CGGCGAGCTGCACGCTGCGTCCT | |
| Generic Cre | Primer Fw: CTAGAGCCTGTTTTGCACGTTC |
| Primer Rv: GTTCGCAAGAACCTGATGGACA | |
| Prdm1 Cre | Primer Fw: GCCGAGGTGCGCGTCAGTAC |
| Primer Rv: CTGAACATGTCCATCAGGTTCTTG | |
| Lef1KotmGro | Primer 24: CCGTTTCAGTGGCACGCCCTCTCC |
| Primer 25: TGTCTCTCTTTCCGTGCTAGTTC | |
| Primer 26: ATGGCGATGCCTGCTTGCCGAATA | |
| ROSAYFP | Primer 1: AAGGGAGCTGCAGTGGAGTA |
| Primer2: CCGAAAATCTGTGGGAAGTC | |
| Primer3: ACATGGTCCTGCTGGAGTTC | |
| Primer4: GGCATTAAAGCAGCGTATCC | |
| Prdm1lox/lox | Primer common A: CCTGGTTAGTAGTTGAATGGGAGC |
| Primer F1A: GTGCTTTTCTTGTGTTGGGAGG | |
| Primer F2A: AGCAGTGTTTCTGAGTGGGTGG |
| Antibody | Species | Dilution | Clone | Company |
|---|---|---|---|---|
| Prdm1 | Mouse | 1:500 | 3H2-E8 | Abcam |
| Sox2 | Mouse | 1:2000 | 9-9-3 | Abcam |
| p75 | Mouse | 1:1000 | 9G395 | UsBiologicals |
| GFP | Goat | 1:1000 | 6673 | Abcam |
| CD31 | Rat | 1:500 | MEC13.3 | BD Biopharmingen |
| Ng2 | Rabbit | 1:300 | AB5320 | Millipore |
| AR | Rabbit | 1:5000 | PG-21 | Millipore |
| Tuj1 | Goat | 1:300 | MMS-435P | Covance |
| Tuj1 | Rabbit | 1:300 | MRB-435P | Covance |
| Pdgfrb | Rabbit | 1:300 | 28E1 | Cell Signaling |
| GFP | Goat | 1:1000 | NB100-NB1770 | Novus |
| Viewpoint | Name | Sequence (5′->3′) |
|---|---|---|
| Lef1 promoter | PromNla3Illumina | AATGATACGGCGACCACCGAACACTCTTTCCCTACACGA CGCTCTTCCGATCTTTAAACAGGGCTACCCTTAAAACCA |
| Lef1 promoter | PromDpn5Illumina | CAAGCAGAAGACGGCATACGAAGGCTCAGTCTTCATCC ACACC |
| Leaf | EnNla2Illumina | AATGATACGGCGACCACCGAACACTCTTTCCCTACACG ACGCTCTTCCGATCTCCGGAAGCGGCTGTTCTC |
| Leaf | EnDpn1Illumina | CAAGCAGAAGACGGCATACGAGGTGGAGAACGGAACC CAAG |
| Data Type | Website | Accession |
|---|---|---|
| 4C-Seq | https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE193356, accessed on 12 January 2022 | GSE193356 |
| CUT&Tag | https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE192851, accessed on 12 January 2022 | GSE192851 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manti, P.G.; Darbellay, F.; Leleu, M.; Coughlan, A.Y.; Moret, B.; Cuennet, J.; Droux, F.; Stoudmann, M.; Mancini, G.-F.; Hautier, A.; et al. The Transcriptional Regulator Prdm1 Is Essential for the Early Development of the Sensory Whisker Follicle and Is Linked to the Beta-Catenin First Dermal Signal. Biomedicines 2022, 10, 2647. https://doi.org/10.3390/biomedicines10102647
Manti PG, Darbellay F, Leleu M, Coughlan AY, Moret B, Cuennet J, Droux F, Stoudmann M, Mancini G-F, Hautier A, et al. The Transcriptional Regulator Prdm1 Is Essential for the Early Development of the Sensory Whisker Follicle and Is Linked to the Beta-Catenin First Dermal Signal. Biomedicines. 2022; 10(10):2647. https://doi.org/10.3390/biomedicines10102647
Chicago/Turabian StyleManti, Pierluigi G., Fabrice Darbellay, Marion Leleu, Aisling Y. Coughlan, Bernard Moret, Julien Cuennet, Frederic Droux, Magali Stoudmann, Gian-Filippo Mancini, Agnès Hautier, and et al. 2022. "The Transcriptional Regulator Prdm1 Is Essential for the Early Development of the Sensory Whisker Follicle and Is Linked to the Beta-Catenin First Dermal Signal" Biomedicines 10, no. 10: 2647. https://doi.org/10.3390/biomedicines10102647