
Pan-Tissue and -Cancer Analysis of ROR1 and ROR2 Transcript Variants Identify Novel Functional Significance for an Alternative Splice Variant of ROR1

Abstract
:1. Introduction
2. Materials and Methods
2.1. Sequence Analysis of Transcript Variants of Human ROR1 and ROR2 Genes
2.2. Analysis of ROR1 and ROR2 Gene and Transcript Variant Expression in Healthy Human Tissues
2.3. Analysis of ROR1 and ROR2 Transcript Variant Expression in Human Tumour Tissues
3. Results
3.1. Sequence Analysis of Transcript Variants of Human ROR1 and ROR2 Genes
3.2. Analysis of ROR1 Gene and Transcript Variant Expression in Healthy Human Tissues
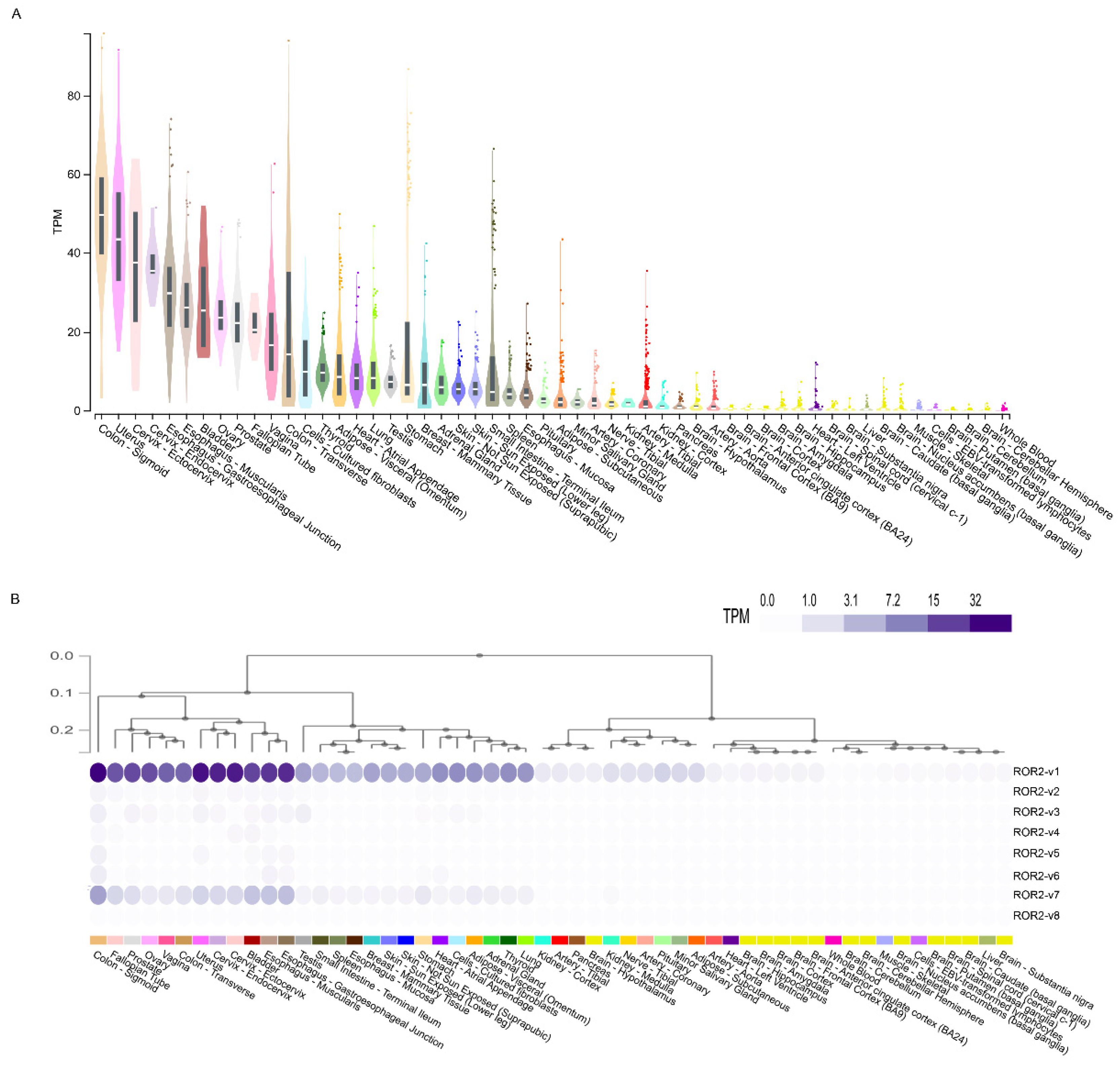
3.3. Analysis of ROR2 Gene and Transcript Variant Expression in Healthy Human Tissues
3.4. Analysis of ROR1 and ROR2 Transcript Variant Expression in Human Tumour Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Katoh, M.; Katoh, M. Comparative genomics on ROR1 and ROR2 orthologs. Oncol. Rep. 2005, 14, 1381–1384. [Google Scholar] [CrossRef] [PubMed]
- Kipps, T.J. ROR1—An Orphan Becomes Apparent. Blood 2022, 140, 1583–1591. [Google Scholar] [CrossRef] [PubMed]
- Menck, K.; Heinrichs, S.; Baden, C.; Bleckmann, A. The WNT/ROR Pathway in Cancer: From Signaling to Therapeutic Intervention. Cells 2021, 10, 142. [Google Scholar] [CrossRef]
- Zhang, S.; Zhang, H.; Ghia, E.M.; Huang, J.; Wu, L.; Zhang, J.; Lam, S.; Lei, Y.; He, J.; Cui, B.; et al. Inhibition of chemotherapy resistant breast cancer stem cells by a ROR1 specific antibody. Proc. Natl. Acad. Sci. USA 2019, 116, 1370–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bicocca, V.T.; Chang, B.H.; Masouleh, B.K.; Muschen, M.; Loriaux, M.M.; Druker, B.J.; Tyner, J.W. Crosstalk between ROR1 and the Pre-B cell receptor promotes survival of t(1;19) acute lymphoblastic leukemia. Cancer Cell 2012, 22, 656–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuda, T.; Chen, L.; Endo, T.; Tang, L.; Lu, D.; Castro, J.E.; Widhopf, G.F.; Rassenti, L.Z.; Cantwell, M.J.; Prussak, C.E.; et al. Antisera induced by infusions of autologous Ad-CD154-leukemia B cells identify ROR1 as an oncofetal antigen and receptor for Wnt5a. Proc. Natl. Acad. Sci. USA 2008, 105, 3047–3052. [Google Scholar] [CrossRef] [Green Version]
- Henry, C.; Llamosas, E.; Knipprath-Meszaros, A.; Schoetzau, A.; Obermann, E.; Fuenfschilling, M.; Caduff, R.; Fink, D.; Hacker, N.; Ward, R.; et al. Targeting the ROR1 and ROR2 receptors in epithelial ovarian cancer inhibits cell migration and invasion. Oncotarget 2015, 6, 40310–40326. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, T.; Yanagisawa, K.; Sugiyama, R.; Hosono, Y.; Shimada, Y.; Arima, C.; Kato, S.; Tomida, S.; Suzuki, M.; Osada, H.; et al. NKX2-1/TITF1/TTF-1-Induced ROR1 is required to sustain EGFR survival signaling in lung adenocarcinoma. Cancer Cell 2012, 21, 348–361. [Google Scholar] [CrossRef] [Green Version]
- Yu, J.; Chen, Y.; Chen, L.; Zhang, L.; Rassenti, L.Z.; Widhopf, G.F., 2nd; Kipps, T.J. Cirmtuzumab inhibits ibrutinib-resistant, Wnt5a-induced Rac1 activation and proliferation in mantle cell lymphoma. Oncotarget 2018, 9, 24731–24736. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Gunther, K.; Enriquez, L.A.; Daniels, B.; O’Mara, T.A.; Tang, K.; Spurdle, A.B.; Ford, C.E. ROR1 is upregulated in endometrial cancer and represents a novel therapeutic target. Sci. Rep. 2020, 10, 13906. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, D.; Guo, Y.; Lu, B.; Zhao, Z.J.; Xu, X.; Chen, Y. Tyrosine Kinase ROR1 as a Target for Anti-Cancer Therapies. Front. Oncol. 2021, 11, 680834. [Google Scholar] [CrossRef] [PubMed]
- Shirangi, A.; Mottaghitalab, F.; Dinarvand, S.; Atyabi, F. Theranostic silk sericin/SPION nanoparticles for targeted delivery of ROR1 siRNA: Synthesis, characterization, diagnosis and anticancer effect on triple-negative breast cancer. Int. J. Biol. Macromol. 2022, 221, 604–612. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Nerreter, T.; Mestermann, K.; Wachter, J.; Chang, J.; Hudecek, M.; Rader, C. ROR1-targeting switchable CAR-T cells for cancer therapy. Oncogene 2022, 41, 4104–4114. [Google Scholar] [CrossRef] [PubMed]
- Jiang, V.C.; Liu, Y.; Jordan, A.; McIntosh, J.; Li, Y.; Che, Y.; Jessen, K.A.; Lannutti, B.J.; Wang, M. The antibody drug conjugate VLS-101 targeting ROR1 is effective in CAR T-resistant mantle cell lymphoma. J. Hematol. Oncol. 2021, 14, 132. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Chen, L.; Cui, B.; Chuang, H.Y.; Yu, J.; Wang-Rodriguez, J.; Tang, L.; Chen, G.; Basak, G.W.; Kipps, T.J. ROR1 is expressed in human breast cancer and associated with enhanced tumor-cell growth. PLoS ONE 2012, 7, e31127. [Google Scholar] [CrossRef] [Green Version]
- Berger, C.; Sommermeyer, D.; Hudecek, M.; Berger, M.; Balakrishnan, A.; Paszkiewicz, P.J.; Kosasih, P.L.; Rader, C.; Riddell, C.R. Safety of targeting ROR1 in primates with chimeric antigen receptor-modified T cells. Cancer Immunol. Res. 2015, 3, 206–216. [Google Scholar] [CrossRef] [Green Version]
- Chang, H.; Jung, W.Y.; Kang, Y.; Lee, H.; Kim, A.; Kim, B.H. Expression of ROR1, pAkt, and pCREB in gastric adenocarcinoma. Ann. Diagn. Pathol. 2015, 19, 330–334. [Google Scholar] [CrossRef]
- Chien, H.P.; Ueng, S.H.; Chen, S.C.; Chang, Y.S.; Lin, Y.C.; Lo, Y.F.; Chang, H.K.; Chuang, W.Y.; Huang, Y.T.; Cheung, Y.C.; et al. Expression of ROR1 has prognostic significance in triple negative breast cancer. Virchows Arch. 2016, 468, 589–595. [Google Scholar] [CrossRef]
- Liu, Y.; Yang, H.; Chen, T.; Luo, Y.; Xu, Z.; Li, Y.; Yang, J. Silencing of Receptor Tyrosine Kinase ROR1 Inhibits Tumor-Cell Proliferation via PI3K/AKT/mTOR Signaling Pathway in Lung Adenocarcinoma. PLoS ONE 2015, 10, e0127092. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Qiu, J.; Ye, C.; Yang, D.; Gao, L.; Su, Y.; Tang, X.; Xu, N.; Zhang, D.; Xiong, L.; et al. ROR1 expression correlated with poor clinical outcome in human ovarian cancer. Sci. Rep. 2014, 4, 5811. [Google Scholar] [CrossRef]
- Liu, D.; Sharbeen, G.; Phillips, P.; Johns, A.L.; Gill, A.J.; Chantrill, L.A.; Timpson, P.; Chou, A.; Pajic, M.; Dwarte, T.; et al. ROR1 and ROR2 expression in pancreatic cancer. BMC Cancer 2021, 21, 1199. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, A.; Goodpaster, T.; Randolph-Habecker, J.; Hoffstrom, B.G.; Jalikis, F.G.; Koch, L.K.; Berger, C.; Kosasih, P.L.; Rajan, A.; Sommermeyer, D.; et al. Analysis of ROR1 Protein Expression in Human Cancer and Normal Tissues. Clin. Cancer Res. 2017, 23, 3061–3071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lara, E.; Calvanese, V.; Huidobro, C.; Fernandez, A.F.; Moncada-Pazos, A.; Obaya, A.J.; Aguilera, O.; Gonzalez-Sancho, J.M.; Sanchez, L.; Astudillo, A.; et al. Epigenetic repression of ROR2 has a Wnt-mediated, pro-tumourigenic role in colon cancer. Mol. Cancer 2010, 9, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, S.S.; Henry, C.E.; Llamosas, E.; Higgins, R.; Daniels, B.; Hesson, L.B.; Hawkins, N.J.; Ward, R.L.; Ford, C.E. Validation of specificity of antibodies for immunohistochemistry: The case of ROR2. Virchows Arch. 2017, 470, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Ma, S.S.; Srivastava, S.; Llamosas, E.; Hawkins, N.J.; Hesson, L.B.; Ward, R.L.; Ford, C.E. ROR2 is epigenetically inactivated in the early stages of colorectal neoplasia and is associated with proliferation and migration. BMC Cancer 2016, 16, 508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mei, H.; Lian, S.; Zhang, S.; Wang, W.; Mao, Q.; Wang, H. High expression of ROR2 in cancer cell correlates with unfavorable prognosis in colorectal cancer. Biochem. Biophys. Res. Commun. 2014, 453, 703–709. [Google Scholar] [CrossRef]
- Li, R.; Liu, T.; Shi, J.; Luan, W.; Wei, X.; Yu, J.; Mao, H.; Liu, P. ROR2 induces cell apoptosis via activating IRE1alpha/JNK/CHOP pathway in high-grade serous ovarian carcinoma in vitro and in vivo. J. Transl. Med. 2019, 17, 428. [Google Scholar] [CrossRef]
- Pan, Q.; Shai, O.; Lee, L.J.; Frey, B.J.; Blencowe, B.J. Deep surveying of alternative splicing complexity in the human transcriptome by high-throughput sequencing. Nat. Genet. 2008, 40, 1413–1415. [Google Scholar] [CrossRef]
- Tress, M.L.; Martelli, P.L.; Frankish, A.; Reeves, G.A.; Wesselink, J.J.; Yeats, C.; Olason, P.I.; Albrecht, M.; Hegyi, H.; Giorgetti, A.; et al. The implications of alternative splicing in the ENCODE protein complement. Proc. Natl. Acad. Sci. USA 2007, 104, 5495–5500. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez, J.M.; Rodriguez-Rivas, J.; Di Domenico, T.; Vazquez, J.; Valencia, A.; Tress, M.L. APPRIS 2017: Principal isoforms for multiple gene sets. Nucleic Acids Res. 2018, 46, D213–D217. [Google Scholar] [CrossRef]
- Almagro Armenteros, J.J.; Tsirigos, K.D.; Sonderby, C.K.; Petersen, T.N.; Winther, O.; Brunak, S.; von Heijne, G.; Nielsen, H. SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 2019, 37, 420–423. [Google Scholar]
- Consortium, G.T. Human genomics. The Genotype-Tissue Expression (GTEx) pilot analysis: Multitissue gene regulation in humans. Science 2015, 348, 648–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, Y.; Liu, C.; Zhou, J.; Rong, X.; Wang, H. Molecular, functional, and gene expression analysis of zebrafish Ror1 receptor. Fish Physiol. Biochem. 2019, 45, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.Y.; Widhopf, G.F., 2nd; Ghia, E.M.; Kidwell, R.L.; Hasan, M.K.; Yu, J.; Rassenti, L.Z.; Chen, L.; Chen, Y.; Pittman, E.; et al. Phase I Trial: Cirmtuzumab Inhibits ROR1 Signaling and Stemness Signatures in Patients with Chronic Lymphocytic Leukemia. Cell Stem Cell 2018, 22, 951–959.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tachon, G.; Cortes, U.; Richard, S.; Martin, S.; Milin, S.; Evrard, C.; Lamour, C.; Karayan-Tapon, L. Targeted RNA-sequencing assays: A step forward compared to FISH and IHC techniques? Cancer Med. 2019, 8, 7556–7566. [Google Scholar] [CrossRef] [Green Version]
- Manzoni, C.; Kia, D.A.; Vandrovcova, J.; Hardy, J.; Wood, N.W.; Lewis, P.A.; Ferrari, R. Genome, transcriptome and proteome: The rise of omics data and their integration in biomedical sciences. Brief. Bioinform. 2018, 19, 286–302. [Google Scholar] [CrossRef] [Green Version]
- Dave, H.; Anver, M.R.; Butcher, D.O.; Brown, P.; Khan, J.; Wayne, A.S.; Baskar, S.; Rader, C. Restricted cell surface expression of receptor tyrosine kinase ROR1 in pediatric B-lineage acute lymphoblastic leukemia suggests targetability with therapeutic monoclonal antibodies. PLoS ONE 2012, 7, e52655. [Google Scholar] [CrossRef] [Green Version]
- Baskar, S.; Kwong, K.Y.; Hofer, T.; Levy, J.M.; Kennedy, M.G.; Lee, E.; Staudt, L.M.; Wilson, W.H.; Wiestner, A.; Rader, C. Unique cell surface expression of receptor tyrosine kinase ROR1 in human B-cell chronic lymphocytic leukemia. Clin. Cancer Res. 2008, 14, 396–404. [Google Scholar] [CrossRef] [Green Version]
- Henry, C.; Quadir, A.; Hawkins, N.J.; Jary, E.; Llamosas, E.; Kumar, D.; Daniels, B.; Ward, R.L.; Ford, C.E. Expression of the novel Wnt receptor ROR2 is increased in breast cancer and may regulate both beta-catenin dependent and independent Wnt signalling. J. Cancer Res. Clin. Oncol. 2015, 141, 243–254. [Google Scholar] [CrossRef]
- Masiakowski, P.; Carroll, R.D. A novel family of cell surface receptors with tyrosine kinase-like domain. J. Biol. Chem. 1992, 267, 26181–26190. [Google Scholar] [CrossRef]
- Liu, Y.; Rubin, B.; Bodine, P.V.; Billiard, J. Wnt5a induces homodimerization and activation of Ror2 receptor tyrosine kinase. J. Cell Biochem. 2008, 105, 497–502. [Google Scholar] [CrossRef] [PubMed]
- Oishi, I.; Takeuchi, S.; Hashimoto, R.; Nagabukuro, A.; Ueda, T.; Liu, Z.J.; Hatta, T.; Akira, S.; Matsuda, Y.; Yamamura, H.; et al. Spatio-temporally regulated expression of receptor tyrosine kinases, mRor1, mRor2, during mouse development: Implications in development and function of the nervous system. Genes Cells 1999, 4, 41–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bainbridge, T.W.; DeAlmeida, V.I.; Izrael-Tomasevic, A.; Chalouni, C.; Pan, B.; Goldsmith, J.; Schoen, A.P.; Quinones, G.A.; Kelly, R.; Lill, J.R.; et al. Evolutionary divergence in the catalytic activity of the CAM-1, ROR1 and ROR2 kinase domains. PLoS ONE 2014, 9, e102695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, J.M.; Zhang, Q.; Young, S.N.; Reese, M.L.; Bailey, F.P.; Eyers, P.A.; Ungureanu, D.; Hammaren, H.; Silvennoinen, O.; Varghese, L.N.; et al. A robust methodology to subclassify pseudokinases based on their nucleotide-binding properties. Biochem. J. 2014, 457, 323–334. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transcript Name | Transcript ID | Transcript Length (bp) | Protein Length (aa) | Transcript Classification | Annotation | ||||
|---|---|---|---|---|---|---|---|---|---|
| GENCODE | Ensemble | APPRIS | MANE Select | Transcript Support Level | |||||
| ROR1-v1 | ENST00000371079.6 | 5858 | 937 | Protein coding | basic | Canonical | P1 | MANE Select v0.93 | TSL 1 |
| ROR1-v2 | ENST00000371080.5 | 2305 | 393 | Protein coding | basic | TSL 1 | |||
| ROR1-v3 | ENST00000545203.2 | 5362 | 882 | Protein coding | basic | TSL 5 | |||
| ROR1-v4 | ENST00000482426.1 | 900 | No protein | Processed transcript | TSL 5 | ||||
| ROR2-v1 | ENST00000375708.4 | 4158 | 943 | Protein coding | basic | Canonical | P1 | MANE Select v0.93 | TSL 1 |
| ROR2-v2 | ENST00000375715.5 | 2993 | 704 | Protein coding | basic | TSL 1 | |||
| ROR2-v3 | ENST00000550066.5 | 4360 | No protein | Processed transcript | TSL 2 | ||||
| ROR2-v4 | ENST00000495386.5 | 664 | No protein | Processed transcript | TSL 3 | ||||
| ROR2-v5 | ENST00000546883.1 | 501 | No protein | Processed transcript | TSL 3 | ||||
| ROR2-v6 | ENST00000493846.1 | 370 | No protein | Processed transcript | TSL 3 | ||||
| ROR2-v7 | ENST00000476440.1 | 316 | No protein | Processed transcript | TSL 5 | ||||
| ROR2-v8 | ENST00000548585.2 | 334 | No protein | Retained intron | TSL 5 | ||||
| Gene | Transcript Name | Transcript ID | Prediction of Signal Peptide | ||||
|---|---|---|---|---|---|---|---|
| SignalP 5.0 | DeepSig | Philius | Phobius | Predisi | |||
| ROR1 | ROR1-v1 | ENST00000371079.6 | SP (1–29) | SP (1–29) | SP (1–29) | SP (1–29) | SP (1–29) |
| ROR1-v2 | ENST00000371080.5 | SP (1–29) | SP (1–29) | SP (1–29) | SP (1–29) | SP (1–29) | |
| ROR1-v3 | ENST00000545203.2 | - | - | - | - | - | |
| ROR2 | ROR2-v1 | ENST00000375708.4 | SP (1–33) | SP (1–22) | SP (1–33) | SP (1–33) | SP (1–33) |
| ROR2-v2 | ENST00000375715.5 | - | - | - | - | - | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
John, M.; Ford, C.E. Pan-Tissue and -Cancer Analysis of ROR1 and ROR2 Transcript Variants Identify Novel Functional Significance for an Alternative Splice Variant of ROR1. Biomedicines 2022, 10, 2559. https://doi.org/10.3390/biomedicines10102559
John M, Ford CE. Pan-Tissue and -Cancer Analysis of ROR1 and ROR2 Transcript Variants Identify Novel Functional Significance for an Alternative Splice Variant of ROR1. Biomedicines. 2022; 10(10):2559. https://doi.org/10.3390/biomedicines10102559
Chicago/Turabian StyleJohn, Miya, and Caroline E. Ford. 2022. "Pan-Tissue and -Cancer Analysis of ROR1 and ROR2 Transcript Variants Identify Novel Functional Significance for an Alternative Splice Variant of ROR1" Biomedicines 10, no. 10: 2559. https://doi.org/10.3390/biomedicines10102559