
A Candidate Therapeutic Monoclonal Antibody Inhibits Both HRSV and HMPV Replication in Mice
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Human Samples
2.2. HMPV and HRSV Fusion Protein Production
2.3. Isolation of HMPV and RSV Specific B Cells
2.4. Monoclonal Antibody Production and Characterization
2.5. HMPV ELISA
2.6. HRSV ELISA
2.7. HRSV and HMPV Viruses
2.8. HRSV and HMPV Microneutralization
2.9. In Vivo Efficacy
2.10. Lung Viral Titers
2.11. Statistics
3. Results
3.1. Isolation of HRSV and HMPV Fusion Protein Specific B Cells
3.2. Production and Characterization of Binding Affinity against HMPV and HRSV
3.3. Neutralization Capacity of Antibodies with Strong Binding Affinity
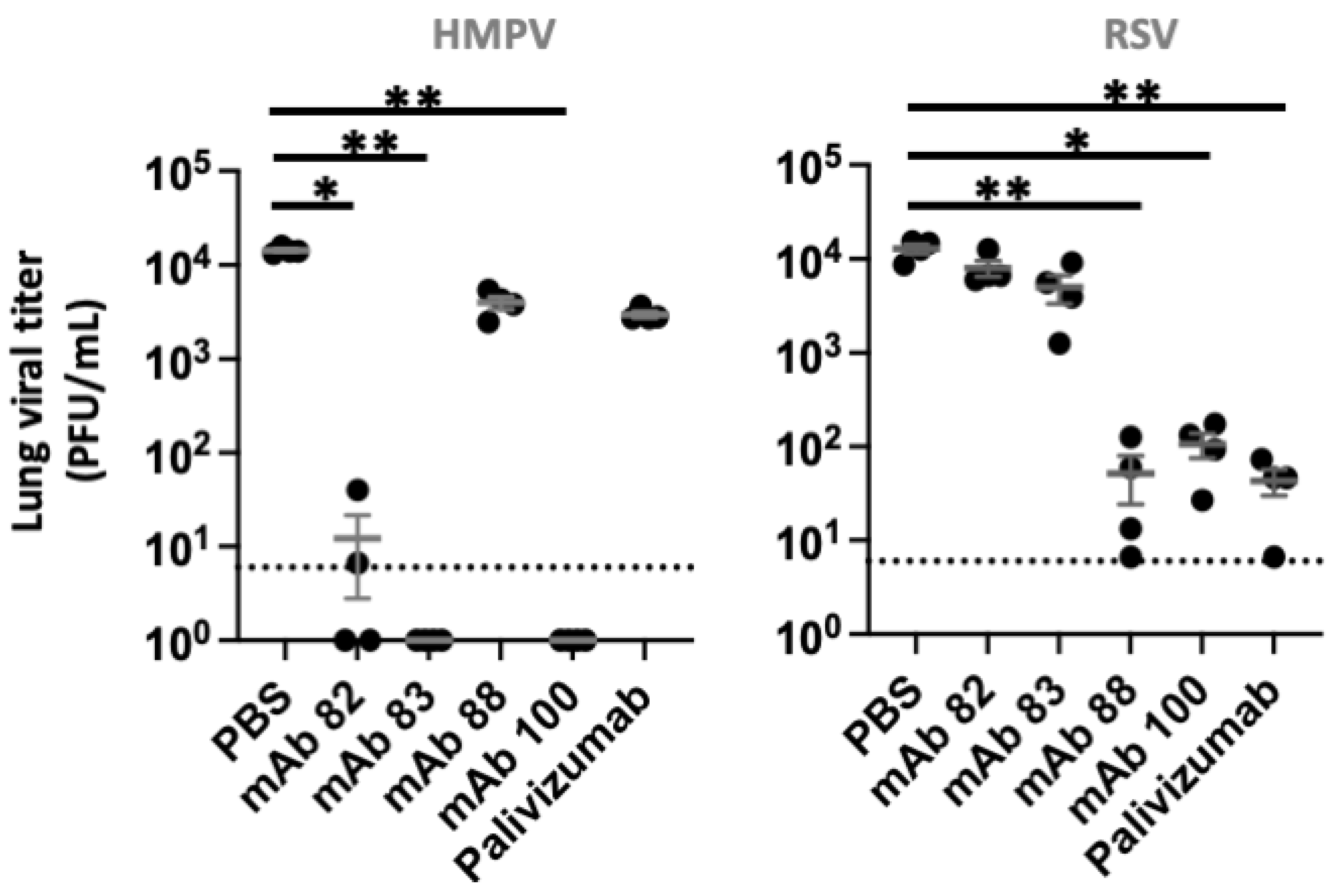
3.4. In Vivo Inhibition of HRSV and HMPV
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hamelin, M.; Boivin, G. Human metapneumovirus: A ubiquitous and long-standing respiratory pathogen. Pediatr. Infect. Dis. J. 2005, 24, S203–S207. [Google Scholar] [CrossRef] [PubMed]
- Rezaee, F.; Linfield, D.T.; Harford, T.; Piedimonte, G. Ongoing developments in RSV prophylaxis: A clinician’s analysis. Curr. Opin. Virol. 2017, 24, 70–78. [Google Scholar] [CrossRef]
- Van Den Hoogen, B.G.; De Jong, J.C.; Groen, J.; Kuiken, T.; De Groot, R.; Fouchier, R.A.; Osterhaus, A.D. A newly discovered human pneumovirus isolated from young children with respiratory tract disease. Nat. Med. 2001, 7, 719–724. [Google Scholar] [CrossRef]
- Leung, J.; Esper, F.; Weibel, C.; Kahn, J.S. Seroepidemiology of Human Metapneumovirus (hMPV) on the Basis of a Novel Enzyme-Linked Immunosorbent Assay Utilizing hMPV Fusion Protein Expressed in Recombinant Vesicular Stomatitis Virus. J. Clin. Microbiol. 2005, 43, 1213–1219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ebihara, T.; Endo, R.; Kikuta, H.; Ishiguro, N.; Yoshioka, M.; Ma, X.; Kobayashi, K. Seroprevalence of human metapneumovirus in Japan. J. Med. Virol. 2003, 70, 281–283. [Google Scholar] [CrossRef]
- Cusi, M.G.; Terrosi, C.; Kleines, M.; Schildgen, O. RSV and HMPV seroprevalence in Tuscany (Italy) and North-Rhine Westfalia (Germany) in the winter season 2009/2010. Influenza Other Respir. Viruses 2011, 5, 380–381. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyiro, J.U.; Kombe, I.K.; Sande, C.J.; Kipkoech, J.; Kiyuka, P.; Onyango, C.O.; Munywoki, P.K.; Kinyanjui, T.M.; Nokes, D.J. Defining the vaccination window for respiratory syncytial virus (RSV) using age-seroprevalence data for children in Kilifi, Kenya. PLoS ONE 2017, 12, e0177803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arankalle, V.A.; Kulkarni, R.; Malshe, N.; Palkar, S.; Lalwani, S.; Mishra, A.C. Seroepidemiology of respiratory syncytial virus in western India with special reference to appropriate age for infant vaccination. J. Med. Virol. 2019, 91, 1566–1570. [Google Scholar] [CrossRef]
- Boivin, G.; De Serres, G.; Côté, S.; Gilca, R.; Abed, Y.; Rochette, L.; Bergeron, M.G.; Déry, P. Human Metapneumovirus Infections in Hospitalized Children1. Emerg. Infect. Dis. 2003, 9, 634–640. [Google Scholar] [CrossRef]
- Zhao, Y.; Lu, R.; Shen, J.; Xie, Z.; Liu, G.; Tan, W. Comparison of viral and epidemiological profiles of hospitalized children with severe acute respiratory infection in Beijing and Shanghai, China. BMC Infect. Dis. 2019, 19, 1–8. [Google Scholar] [CrossRef]
- Freymuth, F.; Vabret, A.; Legrand, L.; Eterradossi, N.; Lafay-Delaire, F.; Brouard, J.; Guillois, B. Presence of the new human metapneumovirus in French children with bronchiolitis. Pediatr. Infect. Dis. J. 2003, 22, 92–94. [Google Scholar] [CrossRef] [PubMed]
- Venter, M.; Lassaunière, R.; Kresfelder, T.L.; Westerberg, Y.; Visser, A. Contribution of common and recently described respiratory viruses to annual hospitalizations in children in South Africa. J. Med. Virol. 2011, 83, 1458–1468. [Google Scholar] [CrossRef] [PubMed]
- Kotaniemi-Syrjänen, A.; Laatikainen, A.; Waris, M.; Reijonen, T.M.; Vainionpää, R.; Korppi, M. Respiratory syncytial virus infection in children hospitalized for wheezing: Virus-specific studies from infancy to preschool years. Acta Paediatr. 2007, 94, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Esper, F.; Martinello, R.A.; Boucher, D.; Weibel, C.; Ferguson, D.; Landry, M.L.; Kahn, J.S. A 1-Year Experience with Human Metapneumovirus in Children Aged <5 Years. J. Infect. Dis. 2004, 189, 1388–1396. [Google Scholar] [CrossRef]
- Ijpma, F.F.; Beekhuis, D.; Cotton, M.F.; Pieper, C.H.; Kimpen, J.L.; Hoogen, B.G.V.D.; van Doornum, G.J.; Osterhaus, D. Human metapneumovirus infection in hospital referred South African children. J. Med. Virol. 2004, 73, 486–493. [Google Scholar] [CrossRef]
- Williams, J.V.; Harris, P.A.; Tollefson, S.J.; Halburnt-Rush, L.L.; Pingsterhaus, J.M.; Edwards, K.M.; Wright, P.F.; Crowe, J.E., Jr. Human Metapneumovirus and Lower Respiratory Tract Disease in Otherwise Healthy Infants and Children. N. Engl. J. Med. 2004, 350, 443–450. [Google Scholar] [CrossRef] [Green Version]
- Acero-Bedoya, S.; Wozniak, P.S.; Sánchez, P.J.; Ramilo, O.; Mejias, A. Recent Trends in RSV Immunoprophylaxis: Clinical Implications for the Infant. Am. J. Perinatol. 2019, 36, S63–S67. [Google Scholar] [CrossRef] [Green Version]
- Storch, G.A. Humanized Monoclonal Antibody for Prevention of Respiratory Syncytial Virus Infection. Pediatrics 1998, 102, 648–651. [Google Scholar] [CrossRef]
- Griffin, M.P.; Yuan, Y.; Takas, T.; Domachowske, J.B.; Madhi, S.A.; Manzoni, P.; Simões, E.A.F.; Esser, M.T.; Khan, A.A.; Dubovsky, F.; et al. Single-Dose Nirsevimab for Prevention of RSV in Preterm Infants. N. Engl. J. Med. 2020, 383, 415–425. [Google Scholar] [CrossRef]
- Aliprantis, A.O.; Wolford, D.; Caro, L.; Maas, B.M.; Ma, H.; Montgomery, D.L.; Sterling, L.M.; Hunt, A.; Cox, K.S.; Vora, K.A.; et al. A Phase 1 Randomized, Double-Blind, Placebo-Controlled Trial to Assess the Safety, Tolerability, and Pharmacokinetics of a Respiratory Syncytial Virus Neutralizing Monoclonal Antibody MK-1654 in Healthy Adults. Clin. Pharmacol. Drug Dev. 2020, 10, 556–566. [Google Scholar] [CrossRef]
- Kumar, P.; Srivastava, M. Prophylactic and therapeutic approaches for human metapneumovirus. VirusDisease 2018, 29, 434–444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Principi, N.; Esposito, S. Paediatric human metapneumovirus infection: Epidemiology, prevention and therapy. J. Clin. Virol. 2014, 59, 141–147. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.I.; Park, S.; Lee, I.; Park, K.S.; Kwak, E.J.; Moon, K.M.; Lee, C.K.; Bae, J.-Y.; Park, M.-S.; Song, K.-J. Genome-Wide Analysis of Human Metapneumovirus Evolution. PLoS ONE 2016, 11, e0152962. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.-F.; Wang, C.K.; Tollefson, S.J.; Piyaratna, R.; Lintao, L.D.; Chu, M.; Liem, A.; Mark, M.; Spaete, R.R.; Crowe, J.E.; et al. Genetic diversity and evolution of human metapneumovirus fusion protein over twenty years. Virol. J. 2009, 6, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sullender, W.M. Respiratory Syncytial Virus Genetic and Antigenic Diversity. Clin. Microbiol. Rev. 2000, 13, 1–15. [Google Scholar] [CrossRef]
- McLellan, J.S.; Chen, M.; Joyce, M.G.; Sastry, M.; Stewart-Jones, G.B.E.; Yang, Y.; Zhang, B.; Chen, L.; Srivatsan, S.; Zheng, A.; et al. Structure-Based Design of a Fusion Glycoprotein Vaccine for Respiratory Syncytial Virus. Science 2013, 342, 592–598. [Google Scholar] [CrossRef] [Green Version]
- Smith, K.; Garman, L.; Wrammert, J.; Zheng, N.-Y.; Capra, J.D.; Ahmed, R.; Wilson, P.C. Rapid generation of fully human monoclonal antibodies specific to a vaccinating antigen. Nat. Protoc. 2009, 4, 372–384. [Google Scholar] [CrossRef] [Green Version]
- Vézina, L.-P.; Faye, L.; Lerouge, P.; D’Aoust, M.-A.; Marquet-Blouin, E.; Burel, C.; Lavoie, P.-O.; Bardor, M.; Gomord, V. Transient co-expression for fast and high-yield production of antibodies with human-like N-glycans in plants. Plant Biotechnol. J. 2009, 7, 442–455. [Google Scholar] [CrossRef]
- Lê, V.B.; Riteau, B.; Alessi, M.-C.; Couture, C.; Jandrot-Perrus, M.; Rhéaume, C.; Hamelin, M.; Boivin, G. Protease-activated receptor 1 inhibition protects mice against thrombin-dependent respiratory syncytial virus and human metapneumovirus infections. Br. J. Pharmacol. 2017, 175, 388–403. [Google Scholar] [CrossRef] [Green Version]
- Gunn, B.M.; Yu, W.-H.; Karim, M.M.; Brannan, J.M.; Herbert, A.S.; Wec, A.Z.; Halfmann, P.J.; Fusco, M.L.; Schendel, S.L.; Gangavarapu, K.; et al. A Role for Fc Function in Therapeutic Monoclonal Antibody-Mediated Protection against Ebola Virus. Cell Host Microbe 2018, 24, 221–233.e5. [Google Scholar] [CrossRef]
- Hamelin, M.-E.; Gagnon, C.; Prince, G.A.; Kiener, P.; Suzich, J.; Ulbrandt, N.; Boivin, G. Prophylactic and therapeutic benefits of a monoclonal antibody against the fusion protein of human metapneumovirus in a mouse model. Antivir. Res. 2010, 88, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Caidi, H.; Miao, C.; Thornburg, N.J.; Tripp, R.A.; Anderson, L.J.; Haynes, L.M. Anti-respiratory syncytial virus (RSV) G monoclonal antibodies reduce lung inflammation and viral lung titers when delivered therapeutically in a BALB/c mouse model. Antivir. Res. 2018, 154, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Cox, R.G.; Williams, J.V. Breaking In: Human Metapneumovirus Fusion and Entry. Viruses 2013, 5, 192–210. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Pan, Y.; Jiang, S.; Lu, L. Respiratory Syncytial Virus Entry Inhibitors Targeting the F Protein. Viruses 2013, 5, 211–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corti, D.; Bianchi, S.; Vanzetta, F.; Minola, A.; Perez, L.; Agatic, G.; Guarino, B.; Silacci, C.; Marcandalli, J.; Marsland, B.J.; et al. Cross-neutralization of four paramyxoviruses by a human monoclonal antibody. Nature 2013, 501, 439–443. [Google Scholar] [CrossRef] [PubMed]
- Schuster, J.E.; Cox, R.G.; Hastings, A.K.; Boyd, K.L.; Wadia, J.; Chen, Z.; Burton, D.R.; Williamson, R.A.; Williams, J.V. A Broadly Neutralizing Human Monoclonal Antibody Exhibits In Vivo Efficacy Against Both Human Metapneumovirus and Respiratory Syncytial Virus. J. Infect. Dis. 2014, 211, 216–225. [Google Scholar] [CrossRef] [Green Version]
- Wen, X.; Mousa, J.J.; Bates, J.T.; Lamb, R.A.; Crowe, J.E., Jr.; Jardetzky, T.S. Structural basis for antibody cross-neutralization of respiratory syncytial virus and human metapneumovirus. Nat. Microbiol. 2017, 2, 16272. [Google Scholar] [CrossRef] [Green Version]
- Xiao, X.; Tang, A.; Cox, K.S.; Wen, Z.; Callahan, C.; Sullivan, N.L.; Nahas, D.D.; Cosmi, S.; Galli, J.D.; Minnier, M.; et al. Characterization of potent RSV neutralizing antibodies isolated from human memory B cells and identification of diverse RSV/hMPV cross-neutralizing epitopes. mAbs 2019, 11, 1415–1427. [Google Scholar] [CrossRef] [Green Version]
- Ilinykh, P.A.; Huang, K.; Santos, R.I.; Gilchuk, P.; Gunn, B.M.; Karim, M.M.; Liang, J.; Fouch, M.E.; Davidson, E.; Parekh, D.V.; et al. Non-neutralizing Antibodies from a Marburg Infection Survivor Mediate Protection by Fc-Effector Functions and by Enhancing Efficacy of Other Antibodies. Cell Host Microbe 2020, 27, 976–991.e11. [Google Scholar] [CrossRef]
- Howell, K.A.; Brannan, J.M.; Bryan, C.; McNeal, A.; Davidson, E.; Turner, H.L.; Vu, H.; Shulenin, S.; He, S.; Kuehne, A.; et al. Cooperativity Enables Non-neutralizing Antibodies to Neutralize Ebolavirus. Cell Rep. 2017, 19, 413–424. [Google Scholar] [CrossRef]
- Lim, S.A.; Gramespacher, J.A.; Pance, K.; Rettko, N.J.; Solomon, P.; Jin, J.; Lui, I.; Elledge, S.K.; Liu, J.; Bracken, C.J.; et al. Bispecific VH/Fab antibodies targeting neutralizing and non-neutralizing Spike epitopes demonstrate enhanced potency against SARS-CoV-2. mAbs 2021, 13, 1893426. [Google Scholar] [CrossRef] [PubMed]
- Earnest, J.T.; Holmes, A.C.; Basore, K.; Mack, M.; Fremont, D.H.; Diamond, M.S. The mechanistic basis of protection by non-neutralizing anti-alphavirus antibodies. Cell Rep. 2021, 35, 108962. [Google Scholar] [CrossRef]
- Stoycheva, D.; Sandu, I.; Gräbnitz, F.; Amorim, A.; Borsa, M.; Weber, S.; Becher, B.; Oxenius, A. Non-neutralizing antibodies protect against chronic LCMV infection by promoting infection of inflammatory monocytes in mice. Eur. J. Immunol. 2021, 51, 1423–1435. [Google Scholar] [CrossRef] [PubMed]
- Excler, J.-L.; Ake, J.; Robb, M.L.; Kim, J.H.; Plotkin, S.A. Nonneutralizing Functional Antibodies: A New “Old” Paradigm for HIV Vaccines. Clin. Vaccine Immunol. 2014, 21, 1023–1036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahnan, W.; Wrighton, S.; Sundwall, M.; Bläckberg, A.; Larsson, O.; Höglund, U.; Khakzad, H.; Godzwon, M.; Walle, M.; Elder, E.; et al. Spike-Dependent Opsonization Indicates Both Dose-Dependent Inhibition of Phagocytosis and That Non-Neutralizing Antibodies Can Confer Protection to SARS-CoV-2. Front. Immunol. 2022, 12, 808932. [Google Scholar] [CrossRef] [PubMed]
- Dunand, C.J.H.; Leon, P.E.; Huang, M.; Choi, A.; Chromikova, V.; Ho, I.Y.; Tan, G.; Cruz, J.; Hirsh, A.; Zheng, N.-Y.; et al. Both Neutralizing and Non-Neutralizing Human H7N9 Influenza Vaccine-Induced Monoclonal Antibodies Confer Protection. Cell Host Microbe 2016, 19, 800–813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, G.S.; Leon, P.E.; Albrecht, R.A.; Margine, I.; Hirsh, A.; Bahl, J.; Krammer, F. Broadly-Reactive Neutralizing and Non-neutralizing Antibodies Directed against the H7 Influenza Virus Hemagglutinin Reveal Divergent Mechanisms of Protection. PLoS Pathog. 2016, 12, e1005578. [Google Scholar] [CrossRef]
- Bootz, A.; Karbach, A.; Spindler, J.; Kropff, B.; Reuter, N.; Sticht, H.; Winkler, T.H.; Britt, W.J.; Mach, M. Protective capacity of neutralizing and non-neutralizing antibodies against glycoprotein B of cytomegalovirus. PLoS Pathog. 2017, 13, e1006601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, W.; Pan, H.W.; Vllasaliu, D.; Lam, J.K.W. Pulmonary Delivery of Biological Drugs. Pharmaceutics 2020, 12, 1025. [Google Scholar] [CrossRef]
- Bittner, B.; Richter, W.; Schmidt, J. Subcutaneous Administration of Biotherapeutics: An Overview of Current Challenges and Opportunities. BioDrugs 2018, 32, 425–440. [Google Scholar] [CrossRef] [PubMed]
- Clarke, D.T.W.; McMillan, D.T.C.a.N.A. Targeted Drug Delivery to the Virus-Infected Airway; Complications and Remedies. Curr. Drug Deliv. 2015, 12, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Respaud, R.; Vecellio, L.; Diot, P.; Heuzé-Vourc’H, N. Nebulization as a delivery method for mAbs in respiratory diseases. Expert Opin. Drug Deliv. 2015, 12, 1027–1039. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| mAbs | HMPV A-16155 (µg/mL) | HMPV B-17480 (µg/mL) | RSV A-23094 (µg/mL) | RSV B-22909 (µg/mL) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 10 | 1 | 0,1 | 10 | 1 | 0,1 | 10 | 1 | 0,1 | 10 | 1 | 0,1 | |
| 29 | +++ | + | - | ++ | ++ | - | - | - | - | - | - | - |
| 82 | +++ | ++++ | +++ | ++++ | ++++ | +++ | - | - | - | - | - | - |
| 83 | ++++ | +++ | +++ | ++++ | +++ | ++ | - | - | - | - | - | - |
| 87 | +++ | +++ | - | +++ | + | - | - | - | - | - | - | - |
| 88 | - | - | - | - | - | - | ++++ | +++ | ++ | +++ | +++ | ++ |
| 89 | +++ | ++ | - | + | - | - | ++ | - | - | - | - | - |
| 90 | +++ | ++ | ++ | +++ | + | + | ++ | + | + | ++ | ++ | - |
| 92 | +++ | ++ | - | +++ | + | - | - | - | - | - | - | - |
| 100 | +++ | ++ | - | +++ | + | + | - | - | - | - | - | - |
| PZB(+) | - | - | - | - | - | - | +++ | ++ | - | ++++ | +++ | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fausther-Bovendo, H.; Hamelin, M.-E.; Carbonneau, J.; Venable, M.-C.; Checkmahomed, L.; Lavoie, P.-O.; Ouellet, M.-È.; Boivin, G.; D’Aoust, M.-A.; Kobinger, G.P. A Candidate Therapeutic Monoclonal Antibody Inhibits Both HRSV and HMPV Replication in Mice. Biomedicines 2022, 10, 2516. https://doi.org/10.3390/biomedicines10102516
Fausther-Bovendo H, Hamelin M-E, Carbonneau J, Venable M-C, Checkmahomed L, Lavoie P-O, Ouellet M-È, Boivin G, D’Aoust M-A, Kobinger GP. A Candidate Therapeutic Monoclonal Antibody Inhibits Both HRSV and HMPV Replication in Mice. Biomedicines. 2022; 10(10):2516. https://doi.org/10.3390/biomedicines10102516
Chicago/Turabian StyleFausther-Bovendo, Hugues, Marie-Eve Hamelin, Julie Carbonneau, Marie-Christine Venable, Liva Checkmahomed, Pierre-Olivier Lavoie, Marie-Ève Ouellet, Guy Boivin, Marc-André D’Aoust, and Gary P. Kobinger. 2022. "A Candidate Therapeutic Monoclonal Antibody Inhibits Both HRSV and HMPV Replication in Mice" Biomedicines 10, no. 10: 2516. https://doi.org/10.3390/biomedicines10102516