
Relationships between Sex and Adaptation to Physical Exercise in Young Athletes: A Pilot Study

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Design
2.2. Participants
2.3. Training Protocol
2.4. Materials
2.5. Procedures
2.5.1. Anthropometric Assessment
2.5.2. Biological Samples
2.6. Plasma Oxidative Stress Measurements
2.7. Adiponectin Western Blot Analysis
2.8. Salivary Cortisol Measurement
2.9. Gas Chromatography–MS (GC–MS) Analysis of Plasma Metabolites
2.10. Extraction of Steroid Hormones and Standard Preparation
2.11. Transesterification of Fatty Acid in Plasma Samples
2.12. Gas Chromatography–Mass Spectrometry (GC–MS) Analysis of Plasma Fatty Acid Methyl Esters (FAMEs)
2.13. Statistical Analysis
3. Results
3.1. Participants’ Characteristics
3.2. Plasma Oxidative Stress and Salivary Cortisol Measurements
3.3. Plasma Adiponectin Determination
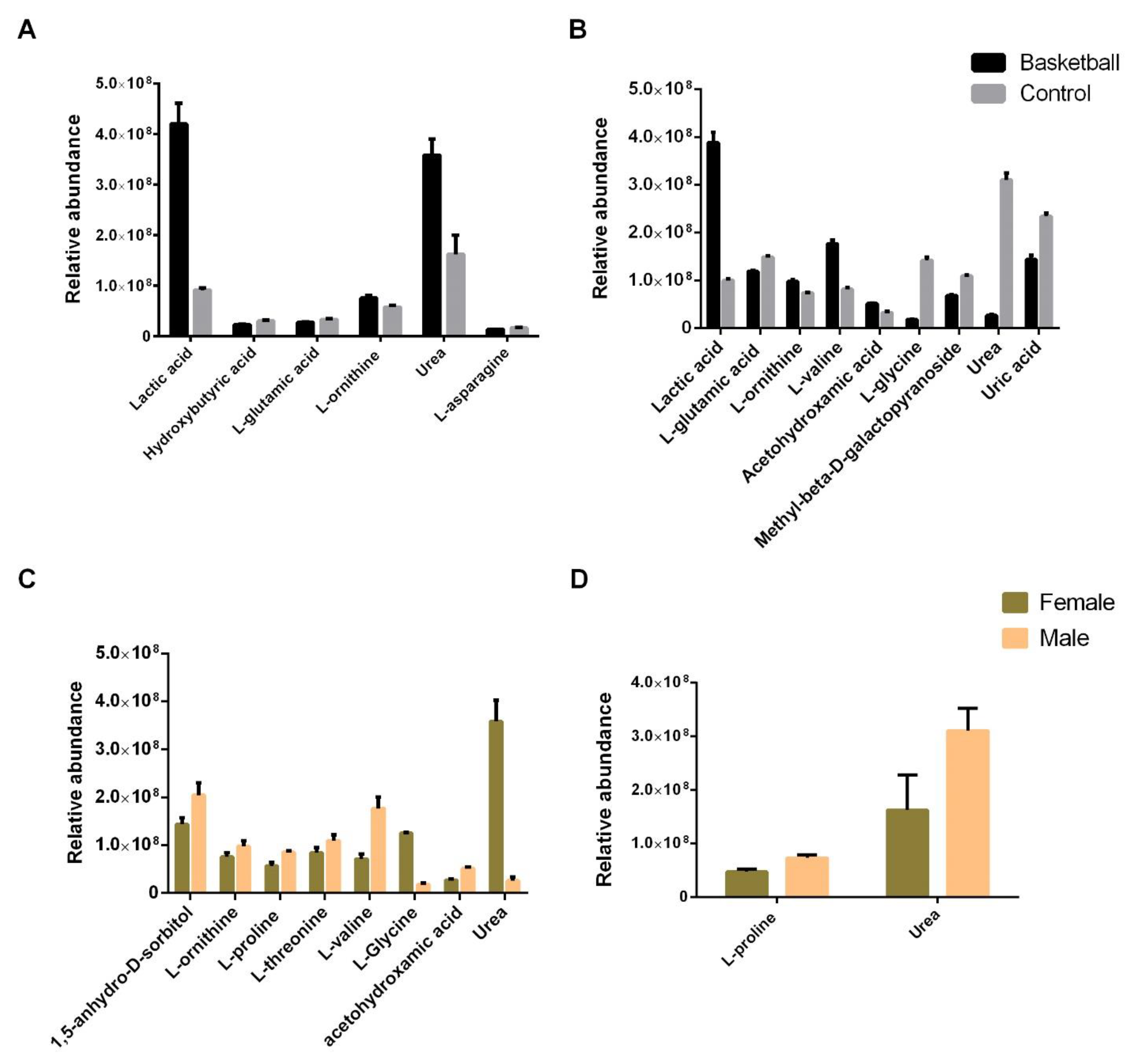
3.4. GC–MS Metabolomic Analysis of Plasma Samples
3.5. Steroid Hormones Evaluation in Plasma Samples
3.6. Fatty Acids Evaluation in Plasma Samples
3.7. Metabolic Pathways Analysis
4. Discussion
5. Limitation
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hawley, J.A.; Lundby, C.; Cotter, J.D.; Burke, L.M. Maximizing Cellular Adaptation to Endurance Exercise in Skeletal Muscle. Cell Metab. 2018, 27, 962–976. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luti, S.; Modesti, A.; Modesti, P.A. Inflammation, Peripheral Signals and Redox Homeostasis in Athletes Who Practice Different Sports. Antioxidants 2020, 9, 1065. [Google Scholar] [CrossRef] [PubMed]
- Hadžović-Džuvo, A.; Valjevac, A.; Lepara, O.; Pjanić, S.; Hadžimuratović, A.; Mekić, A. Oxidative stress status in elite athletes engaged in different sport disciplines. Bosn. J. Basic Med. Sci. 2014, 14, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mangine, G.T.; Hoffman, J.R.; Gonzalez, A.M.; Townsend, J.R.; Wells, A.J.; Jajtner, A.R.; Beyer, K.S.; Boone, C.H.; Wang, R.; Miramonti, A.A.; et al. Exercise-Induced Hormone Elevations Are Related to Muscle Growth. J. Strength Cond. Res. 2017, 31, 45–53. [Google Scholar] [CrossRef]
- Magherini, F.; Fiaschi, T.; Marzocchini, R.; Mannelli, M.; Gamberi, T.; Modesti, P.A.; Modesti, A. Oxidative stress in exercise training: The involvement of inflammation and peripheral signals. Free Radic. Res. 2019, 53, 1155–1165. [Google Scholar] [CrossRef]
- Martinez-Huenchullan, S.F.; Tam, C.S.; Ban, L.A.; Ehrenfeld-Slater, P.; Mclennan, S.V.; Twigg, S.M. Skeletal muscle adiponectin induction in obesity and exercise. Metabolism 2020, 102, 154008. [Google Scholar] [CrossRef] [Green Version]
- Ren, Y.; Li, Y.; Yan, J.; Ma, M.; Zhou, D.; Xue, Z.; Zhang, Z.; Liu, H.; Yang, H.; Jia, L.; et al. Adiponectin modulates oxidative stress-induced mitophagy and protects C2C12 myoblasts against apoptosis. Sci. Rep. 2017, 7, 3209. [Google Scholar] [CrossRef] [Green Version]
- Cnop, M.; Havel, P.J.; Utzschneider, K.M.; Carr, D.B.; Sinha, M.K.; Boyko, E.J.; Retzlaff, B.M.; Knopp, R.H.; Brunzell, J.D.; Kahn, S.E. Relationship of adiponectin to body fat distribution, insulin sensitivity and plasma lipoproteins: Evidence for independent roles of age and sex. Diabetologia 2003, 46, 459–469. [Google Scholar] [CrossRef] [Green Version]
- Böttner, A.; Kratzsch, J.; Müller, G.; Kapellen, T.M.; Blüher, S.; Keller, E.; Blüher, M.; Kiess, W. Gender differences of adiponectin levels develop during the progression of puberty and are related to serum androgen levels. J. Clin. Endocrinol. Metab. 2004, 89, 4053–4061. [Google Scholar] [CrossRef] [Green Version]
- Gamberi, T.; Magherini, F.; Fiaschi, T. Adiponectin in Myopathies. Int. J. Mol. Sci. 2019, 20, 1544. [Google Scholar] [CrossRef] [Green Version]
- Agostinis-Sobrinho, C.; Santos, R.; Moreira, C.; Abreu, S.; Lopes, L.; Oliveira-Santos, J.; Rosário, R.; Póvoas, S.; Mota, J. Association between serum adiponectin levels and muscular fitness in Portuguese adolescents: LabMed Physical Activity Study. Nutr. Metab. Cardiovasc. Dis. 2016, 26, 517–524. [Google Scholar] [CrossRef] [Green Version]
- Senefeld, J.W.; Clayburn, A.J.; Baker, S.E.; Carter, R.E.; Johnson, P.W.; Joyner, M.J. Sex differences in youth elite swimming. PLoS ONE 2019, 14, e0225724. [Google Scholar] [CrossRef]
- Handelsman, D.J.; Hirschberg, A.L.; Bermon, S. Circulating Testosterone as the Hormonal Basis of Sex Differences in Athletic Performance. Endocr. Rev. 2018, 39, 803–829. [Google Scholar] [CrossRef] [Green Version]
- Vesper, H.W.; Wang, Y.; Vidal, M.; Botelho, J.C.; Caudill, S.P. Serum Total Testosterone Concentrations in the US Household Population from the NHANES 2011–2012 Study Population. Clin. Chem. 2015, 61, 1495–1504. [Google Scholar] [CrossRef] [Green Version]
- Handelsman, D.J.; Sikaris, K.; Ly, L.P. Estimating age-specific trends in circulating testosterone and sex hormone-binding globulin in males and females across the lifespan. Ann. Clin. Biochem. 2016, 53, 377–384. [Google Scholar] [CrossRef] [Green Version]
- Handelsman, D.J. Sex differences in athletic performance emerge coinciding with the onset of male puberty. Clin. Endocrinol. 2017, 87, 68–72. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, J.R.; Bar-Eli, M.; Tenenbaum, G. An examination of mood changes and performance in a professional basketball team. J. Sports Med. Phys. Fit. 1999, 39, 74–79. [Google Scholar]
- McInnes, S.E.; Carlson, J.S.; Jones, C.J.; McKenna, M.J. The physiological load imposed on basketball players during competition. J. Sports Sci. 1995, 13, 387–397. [Google Scholar] [CrossRef]
- Ben Abdelkrim, N.; El Fazaa, S.; El Ati, J. Time-motion analysis and physiological data of elite under-19-year-old basketball players during competition. Br. J. Sports Med. 2007, 41, 69–75; discussion 75. [Google Scholar] [CrossRef]
- Scanlan, A.; Dascombe, B.; Reaburn, P. A comparison of the activity demands of elite and sub-elite Australian men’s basketball competition. J. Sports Sci. 2011, 29, 1153–1160. [Google Scholar] [CrossRef]
- Hagströmer, M.; Oja, P.; Sjöström, M. The International Physical Activity Questionnaire (IPAQ): A study of concurrent and construct validity. Public Health Nutr. 2006, 9, 755–762. [Google Scholar] [CrossRef]
- Foster, C.; Florhaug, J.A.; Franklin, J.; Gottschall, L.; Hrovatin, L.A.; Parker, S.; Doleshal, P.; Dodge, C. A new approach to monitoring exercise training. J. Strength Cond. Res. 2001, 15, 109–115. [Google Scholar]
- Militello, R.; Luti, S.; Parri, M.; Marzocchini, R.; Soldaini, R.; Modesti, A.; Modesti, P.A. Redox Homeostasis and Metabolic Profile in Young Female Basketball Players during in-Season Training. Healthcare 2021, 9, 368. [Google Scholar] [CrossRef]
- Luti, S.; Fiaschi, T.; Magherini, F.; Modesti, P.A.; Piomboni, P.; Governini, L.; Luddi, A.; Amoresano, A.; Illiano, A.; Pinto, G.; et al. Relationship between the metabolic and lipid profile in follicular fluid of women undergoing in vitro fertilization. Mol. Reprod. Dev. 2020, 87, 986–997. [Google Scholar] [CrossRef]
- McGuigan, M.R.; Egan, A.D.; Foster, C. Salivary Cortisol Responses and Perceived Exertion during High Intensity and Low Intensity Bouts of Resistance Exercise. J. Sports Sci. Med. 2004, 3, 8–15. [Google Scholar]
- Gao, J.; Tarcea, V.G.; Karnovsky, A.; Mirel, B.R.; Weymouth, T.E.; Beecher, C.W.; Cavalcoli, J.D.; Athey, B.D.; Omenn, G.S.; Burant, C.F.; et al. Metscape: A Cytoscape plug-in for visualizing and interpreting metabolomic data in the context of human metabolic networks. Bioinformatics 2010, 26, 971–973. [Google Scholar] [CrossRef] [Green Version]
- Wiecek, M.; Maciejczyk, M.; Szymura, J.; Szygula, Z. Changes in Oxidative Stress and Acid-Base Balance in Men and Women Following Maximal-Intensity Physical Exercise. Physiol. Res. 2015, 64, 93–102. [Google Scholar] [CrossRef]
- Spanidis, Y.; Goutzourelas, N.; Stagos, D.; Mpesios, A.; Priftis, A.; Bar-Or, D.; Spandidos, D.A.; Tsatsakis, A.M.; Leon, G.; Kouretas, D. Variations in oxidative stress markers in elite basketball players at the beginning and end of a season. Exp. Ther. Med. 2016, 11, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Yilmaz, N.; Erel, Ö.; Hazer, M.; Bagci, C.; Namiduru, E.; Giil, E. Biochemical assessments of retinol, α-tocopherol, pyridoxal—5-phosphate oxidative stress index and total antioxidant status in adolescent professional basketball players and sedentary controls. Int. J. Adolesc. Med. Health 2007, 19, 177–186. [Google Scholar] [CrossRef]
- Kraemer, W.J.; Ratamess, N.A. Hormonal responses and adaptations to resistance exercise and training. Sports Med. 2005, 35, 339–361. [Google Scholar] [CrossRef]
- Casanova, N.; Palmeira-DE-Oliveira, A.; Pereira, A.; Crisóstomo, L.; Travassos, B.; Costa, A.M. Cortisol, testosterone and mood state variation during an official female football competition. J. Sports Med. Phys. Fit. 2016, 56, 775–781. [Google Scholar]
- Tanner, A.V.; Nielsen, B.V.; Allgrove, J. Salivary and plasma cortisol and testosterone responses to interval and tempo runs and a bodyweight-only circuit session in endurance-trained men. J. Sports Sci. 2014, 32, 680–689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwinn, A.-C.; Sauer, F.J.; Gerber, V.; Bruckmaier, R.M.; Gross, J.J. Free and bound cortisol in plasma and saliva during ACTH challenge in dairy cows and horses. J. Anim. Sci. 2018, 96, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Jurimae, J.; Purge, P.; Jurimae, T. Adiponectin is altered after maximal exercise in highly trained male rowers. Eur. J. Appl. Physiol. 2005, 93, 502–505. [Google Scholar] [CrossRef] [PubMed]
- Plinta, R.; Olszanecka-Glinianowicz, M.; Drosdzol-Cop, A.; Chudek, J.; Skrzypulec-Plinta, V. The effect of three-month pre-season preparatory period and short-term exercise on plasma leptin, adiponectin, visfatin, and ghrelin levels in young female handball and basketball players. J. Endocrinol. Investig. 2012, 35, 595–601. [Google Scholar] [CrossRef]
- Waki, H.; Yamauchi, T.; Kamon, J.; Ito, Y.; Uchida, S.; Kita, S.; Hara, K.; Hada, Y.; Vasseur, F.; Froguel, P.; et al. Impaired multimerization of human adiponectin mutants associated with diabetes. Molecular structure and multimer formation of adiponectin. J. Biol. Chem. 2003, 278, 40352–40363. [Google Scholar] [CrossRef] [Green Version]
- Høeg, L.D.; Sjøberg, K.A.; Lundsgaard, A.-M.; Jordy, A.B.; Hiscock, N.; Wojtaszewski, J.F.P.; Richter, E.A.; Kiens, B. Adiponectin concentration is associated with muscle insulin sensitivity, AMPK phosphorylation, and ceramide content in skeletal muscles of men but not women. J. Appl. Physiol. 2013, 114, 592–601. [Google Scholar] [CrossRef] [Green Version]
- Senefeld, J.; Joyner, M.J.; Stevens, A.; Hunter, S.K. Sex differences in elite swimming with advanced age are less than marathon running. Scand. J. Med. Sci. Sports 2016, 26, 17–28. [Google Scholar] [CrossRef] [Green Version]
- Vingren, J.L.; Kraemer, W.J.; Ratamess, N.A.; Anderson, J.M.; Volek, J.S.; Maresh, C.M. Testosterone physiology in resistance exercise and training: The up-stream regulatory elements. Sports Med. 2010, 40, 1037–1053. [Google Scholar] [CrossRef]
- Heaney, J.L.J.; Carroll, D.; Phillips, A.C. DHEA, DHEA-S and cortisol responses to acute exercise in older adults in relation to exercise training status and sex. Age 2013, 35, 395–405. [Google Scholar] [CrossRef] [Green Version]
- Ennour-Idrissi, K.; Maunsell, E.; Diorio, C. Effect of physical activity on sex hormones in women: A systematic review and meta-analysis of randomized controlled trials. Breast Cancer Res. BCR 2015, 17, 139. [Google Scholar] [CrossRef] [Green Version]
- Mika, A.; Macaluso, F.; Barone, R.; Di Felice, V.; Sledzinski, T. Effect of Exercise on Fatty Acid Metabolism and Adipokine Secretion in Adipose Tissue. Front. Physiol. 2019, 10, 26. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Fu, X.; Chen, Q.; Patra, J.K.; Wang, D.; Wang, Z.; Gai, Z. Arachidonic Acid Metabolism and Kidney Inflammation. Int. J. Mol. Sci. 2019, 20, 3683. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Minegishi, Y.; Hasumura, T.; Shimotoyodome, A.; Ota, N. Involvement of ammonia metabolism in the improvement of endurance performance by tea catechins in mice. Sci. Rep. 2020, 10, 6065. [Google Scholar] [CrossRef] [Green Version]
- Shimomura, Y.; Murakami, T.; Nakai, N.; Nagasaki, M.; Harris, R.A. Exercise promotes BCAA catabolism: Effects of BCAA supplementation on skeletal muscle during exercise. J. Nutr. 2004, 134, 1583S–1587S. [Google Scholar] [CrossRef] [Green Version]
- Shou, J.; Chen, P.-J.; Xiao, W.-H. The Effects of BCAAs on Insulin Resistance in Athletes. J. Nutr. Sci. Vitaminol. 2019, 65, 383–389. [Google Scholar] [CrossRef] [Green Version]
- Comitato, R.; Saba, A.; Turrini, A.; Arganini, C.; Virgili, F. Sex Hormones and Macronutrient Metabolism. Crit. Rev. Food Sci. Nutr. 2015, 55, 227–241. [Google Scholar] [CrossRef] [Green Version]
- Carter, S.L.; Rennie, C.; Tarnopolsky, M.A. Substrate utilization during endurance exercise in men and women after endurance training. Am. J. Physiol. Endocrinol. Metab. 2001, 280, E898–E907. [Google Scholar] [CrossRef] [Green Version]
- Roepstorff, C.; Steffensen, C.H.; Madsen, M.; Stallknecht, B.; Kanstrup, I.-L.; Richter, E.A.; Kiens, B. Gender differences in substrate utilization during submaximal exercise in endurance-trained subjects. Am. J. Physiol. Endocrinol. Metab. 2002, 282, E435–E447. [Google Scholar] [CrossRef] [Green Version]
- Roepstorff, C.; Thiele, M.; Hillig, T.; Pilegaard, H.; Richter, E.A.; Wojtaszewski, J.F.P.; Kiens, B. Higher skeletal muscle alpha2AMPK activation and lower energy charge and fat oxidation in men than in women during submaximal exercise. J. Physiol. 2006, 574, 125–138. [Google Scholar] [CrossRef]
- Brooks, G.A. The Science and Translation of Lactate Shuttle Theory. Cell Metab. 2018, 27, 757–785. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, B.S.; Rogatzki, M.J.; Goodwin, M.L.; Kane, D.A.; Rightmire, Z.; Gladden, L.B. Lactate metabolism: Historical context, prior misinterpretations, and current understanding. Eur. J. Appl. Physiol. 2018, 118, 691–728. [Google Scholar] [CrossRef]
- Kitaoka, Y.; Takeda, K.; Tamura, Y.; Hatta, H. Lactate administration increases mRNA expression of PGC-1α and UCP3 in mouse skeletal muscle. Appl. Physiol. Nutr. Metab. 2016, 41, 695–698. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, T.; Hussien, R.; Oommen, S.; Gohil, K.; Brooks, G.A. Lactate sensitive transcription factor network in L6 cells: Activation of MCT1 and mitochondrial biogenesis. FASEB J. 2007, 21, 2602–2612. [Google Scholar] [CrossRef]
- McNulty, K.L.; Elliott-Sale, K.J.; Dolan, E.; Swinton, P.A.; Ansdell, P.; Goodall, S.; Thomas, K.; Hicks, K.M. The Effects of Menstrual Cycle Phase on Exercise Performance in Eumenorrheic Women: A Systematic Review and Meta-Analysis. Sports Med. 2020, 50, 1813–1827. [Google Scholar] [CrossRef]
- Pallavi, L.C.; Souza, U.J.D.; Shivaprakash, G. Assessment of Musculoskeletal Strength and Levels of Fatigue during Different Phases of Menstrual Cycle in Young Adults. J. Clin. Diagn. Res. JCDR 2017, 11, CC11–CC13. [Google Scholar] [CrossRef]
- Bambaeichi, E.; Reilly, T.; Cable, N.T.; Giacomoni, M. The isolated and combined effects of menstrual cycle phase and time-of-day on muscle strength of eumenorrheic females. Chronobiol. Int. 2004, 21, 645–660. [Google Scholar] [CrossRef]
- Oosthuyse, T.; Bosch, A.N.; Jackson, S. Cycling time trial performance during different phases of the menstrual cycle. Eur. J. Appl. Physiol. 2005, 94, 268–276. [Google Scholar] [CrossRef]
- McLay, R.T.; Thomson, C.D.; Williams, S.M.; Rehrer, N.J. Carbohydrate loading and female endurance athletes: Effect of menstrual-cycle phase. Int. J. Sport Nutr. Exerc. Metab. 2007, 17, 189–205. [Google Scholar] [CrossRef]
- Janse DE Jonge, X.A.K.; Thompson, M.W.; Chuter, V.H.; Silk, L.N.; Thom, J.M. Exercise performance over the menstrual cycle in temperate and hot, humid conditions. Med. Sci. Sports Exerc. 2012, 44, 2190–2198. [Google Scholar] [CrossRef]
- Vaiksaar, S.; Jürimäe, J.; Mäestu, J.; Purge, P.; Kalytka, S.; Shakhlina, L.; Jürimäe, T. No effect of menstrual cycle phase and oral contraceptive use on endurance performance in rowers. J. Strength Cond. Res. 2011, 25, 1571–1578. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rate (°C/min) | Value (°C) | Hold Time (min) | Run Time (min) | |
|---|---|---|---|---|
| (initial) | 90 | 1 | 1 | |
| Ramp 1 | 10 | 140 | 2 | 8 |
| Ramp 2 | 5 | 180 | 0 | 16 |
| Ramp 3 | 10 | 280 | 5 | 31 |
| Characteristics | Mean (SD) | Tukey’s Test a | ||||||
|---|---|---|---|---|---|---|---|---|
| Basket Male | Control Male | Basket Female | Control Female | BM vs. CM | BF vs. CF | BM vs. BF | CM vs. CF | |
| Age (year) | 21 ± 2.2 | 26.1 ± 4.1 | 25.1 ± 5.5 | 26.9 ± 2.2 | 0.01 * | 0.77 | 0.08 | 0.97 |
| Weight (kg) | 81.5 ± 10.2 | 73 ± 8.7 | 68.7 ± 11.9 | 58.7 ± 5.8 | 0.17 | 0.17 | 0.03 * | 0.02 * |
| Height (cm) | 186 ± 0.06 | 178.7 ± 0.06 | 175.6 ± 0.08 | 163.4 ± 0.06 | 0.06 | 0.003 ** | 0.006 ** | <0.0001 **** |
| BMI (kg/m2) | 23.6 ± 2.7 | 22.9 ± 2.9 | 22.1 ± 2.05 | 22 ± 2.3 | 0.64 | 1.00 | 0.18 | 0.72 |
| Metabolic Pathways Involved in Females and Males |
|---|
| Androgen and estrogen biosynthesis and metabolism |
| Arachidonic acid metabolism |
| Bile acid biosynthesis |
| C21-steroid hormone biosynthesis and metabolism |
| De novo fatty acid biosynthesis |
| Di-unsaturated fatty acid beta-oxidation |
| Glycolysis and gluconeogenesis |
| Histidine metabolism |
| Leukotriene metabolism |
| Linoleate metabolism |
| Omega-6 fatty acid metabolism |
| Purine metabolism |
| Urea cycle and metabolism of arginine, proline, glutamate, aspartate, and asparagine |
| Vitamin B9 (folate) metabolism |
| Metabolic pathways involved only in females |
| Butanoate metabolism |
| De novo fatty acid biosynthesis |
| Omega-6 fatty acid metabolism |
| Squalene and cholesterol biosynthesis |
| Metabolic pathways involved only in males |
| Glycine, serine, alanine, and threonine metabolism |
| Lysine metabolism |
| Porphyrin metabolism |
| Prostaglandin formation from arachidonate |
| Saturated fatty acids beta-oxidation |
| Valine, leucine, and isoleucine degradation |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pinto, G.; Militello, R.; Amoresano, A.; Modesti, P.A.; Modesti, A.; Luti, S. Relationships between Sex and Adaptation to Physical Exercise in Young Athletes: A Pilot Study. Healthcare 2022, 10, 358. https://doi.org/10.3390/healthcare10020358
Pinto G, Militello R, Amoresano A, Modesti PA, Modesti A, Luti S. Relationships between Sex and Adaptation to Physical Exercise in Young Athletes: A Pilot Study. Healthcare. 2022; 10(2):358. https://doi.org/10.3390/healthcare10020358
Chicago/Turabian StylePinto, Gabriella, Rosamaria Militello, Angela Amoresano, Pietro Amedeo Modesti, Alessandra Modesti, and Simone Luti. 2022. "Relationships between Sex and Adaptation to Physical Exercise in Young Athletes: A Pilot Study" Healthcare 10, no. 2: 358. https://doi.org/10.3390/healthcare10020358