
Study of Reversible Platelet Aggregation Model by Nonlinear Dynamics



Abstract
:1. Introduction
2. Materials and Methods
2.1. Computational Methods
2.2. Reagents
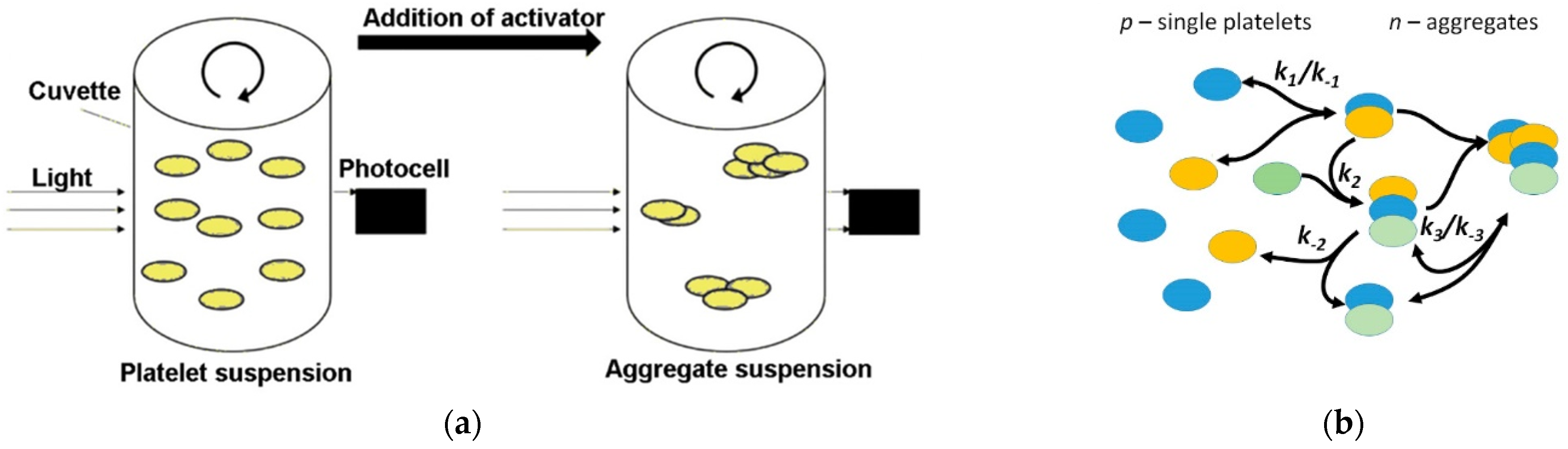
2.3. Blood Collection and Aggregometry
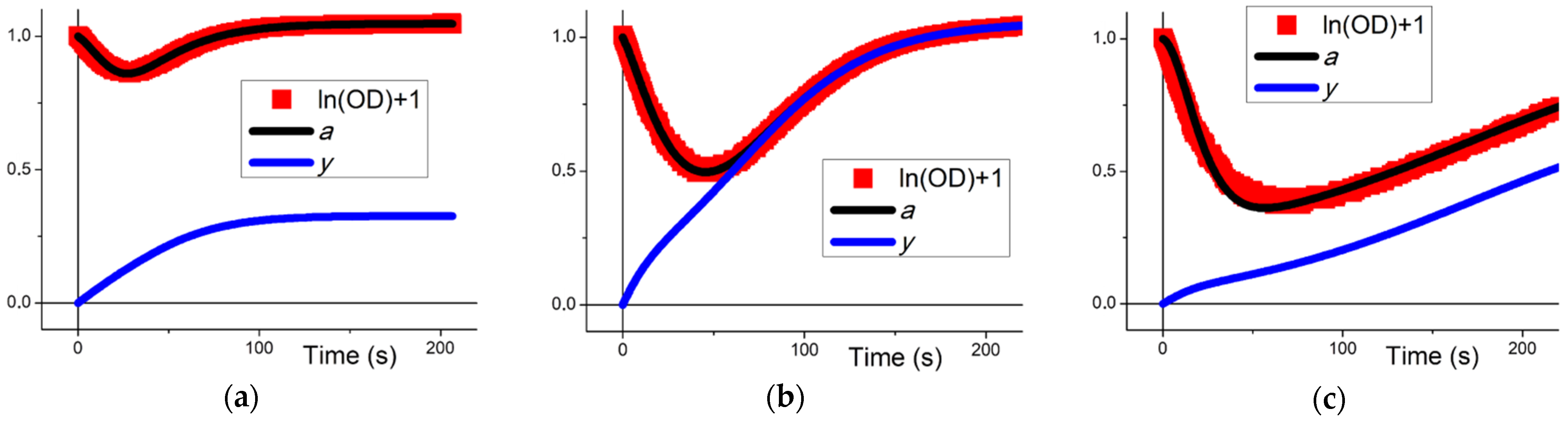
3. Results
3.1. Transformation of the Model
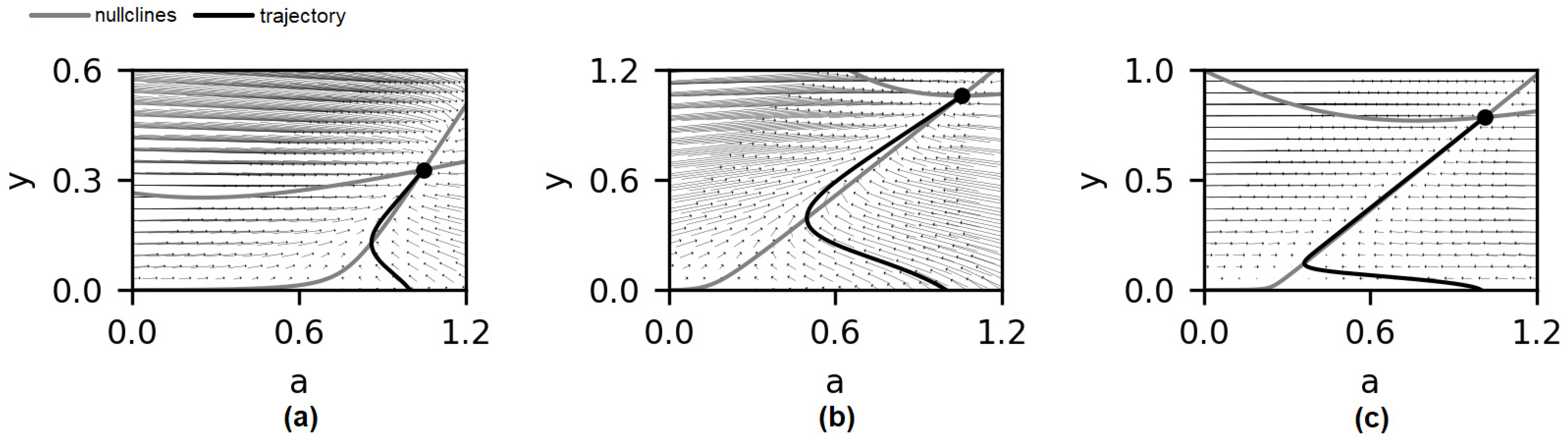
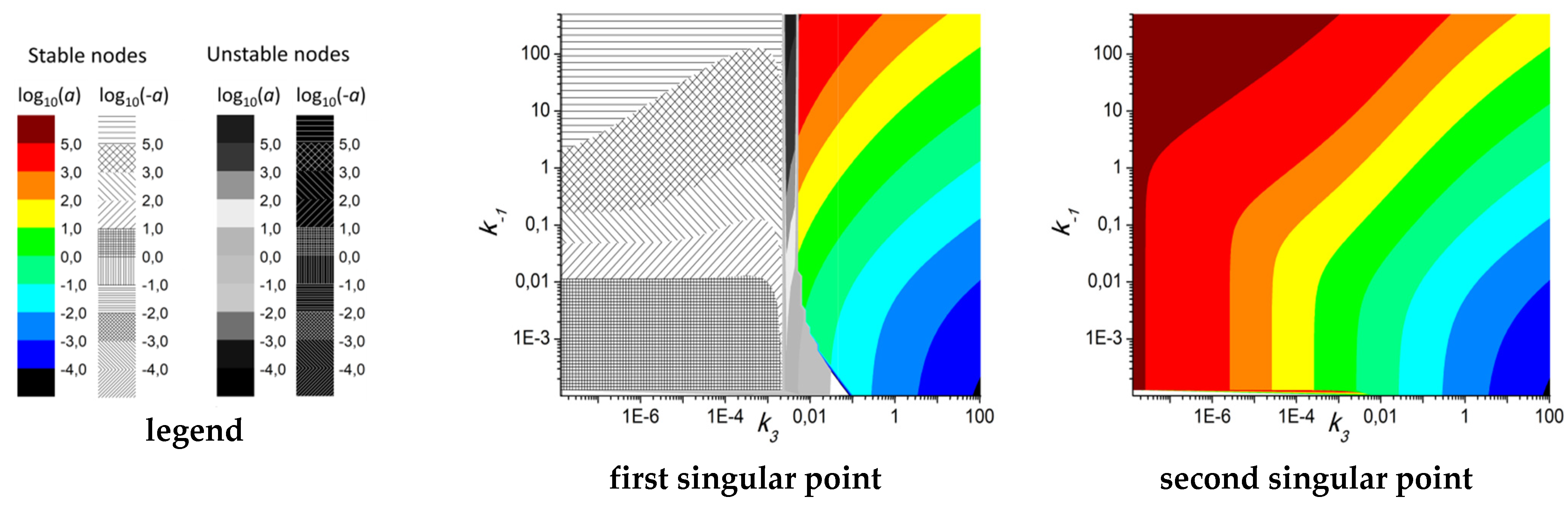
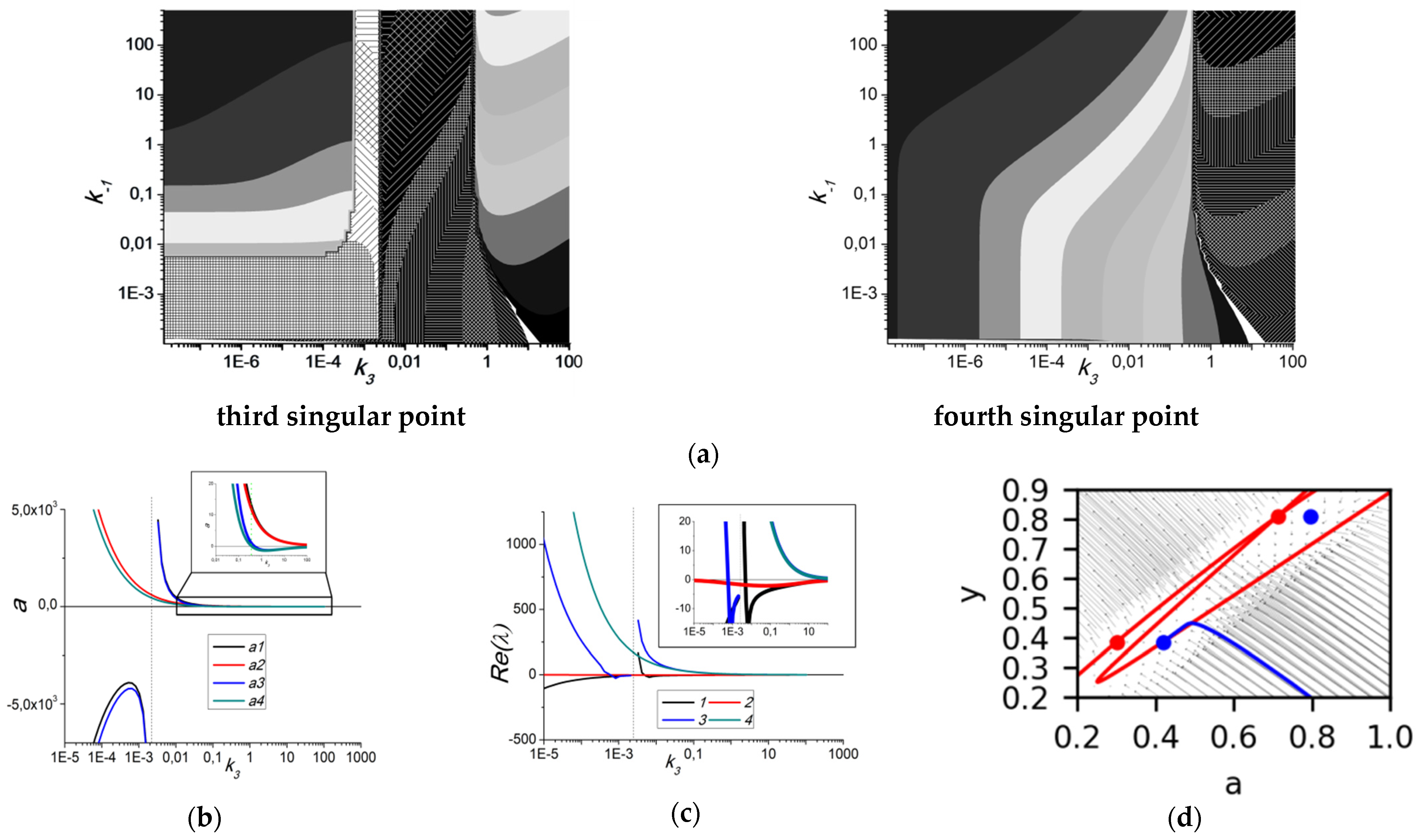
3.2. Bifurcation Analysis of the System
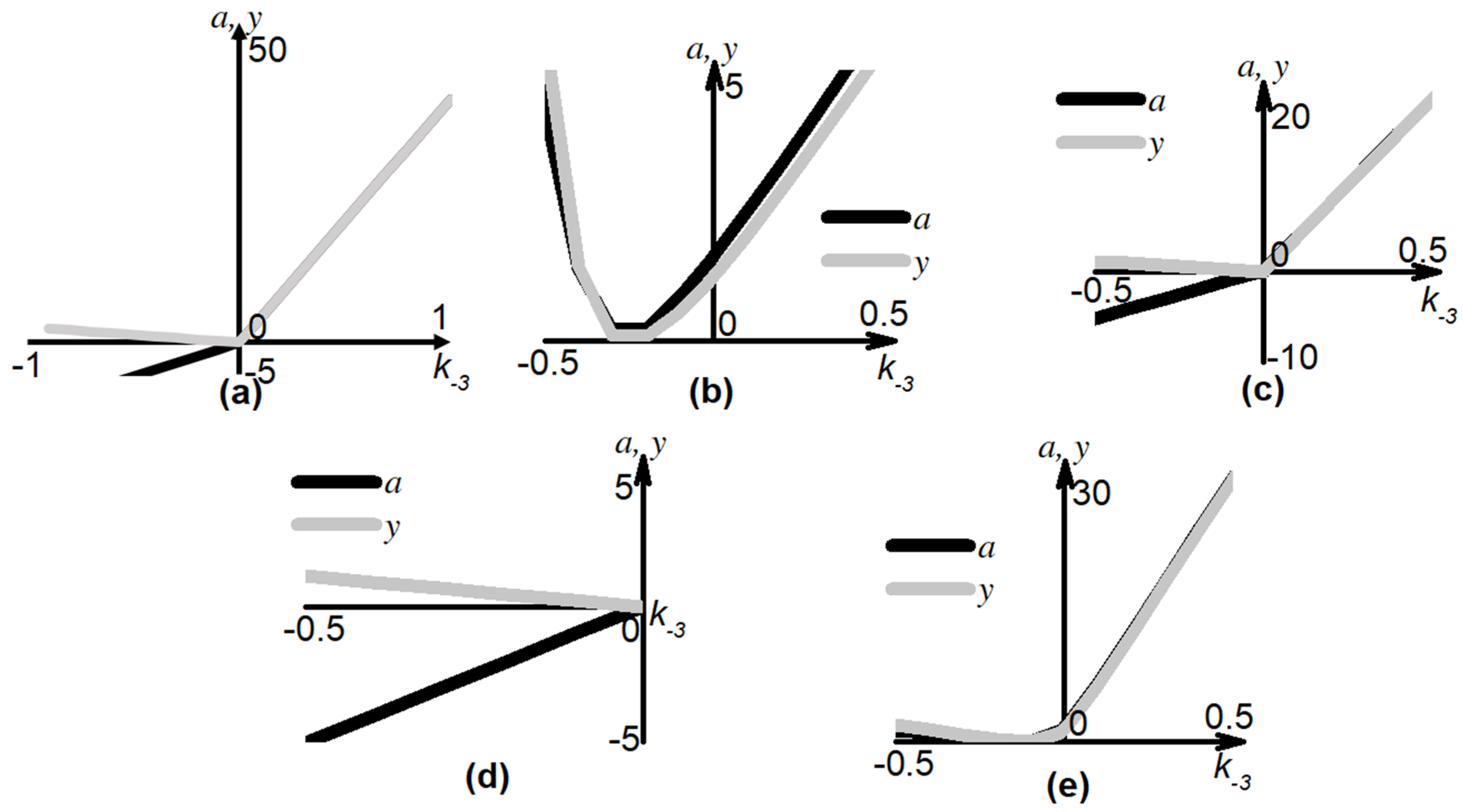
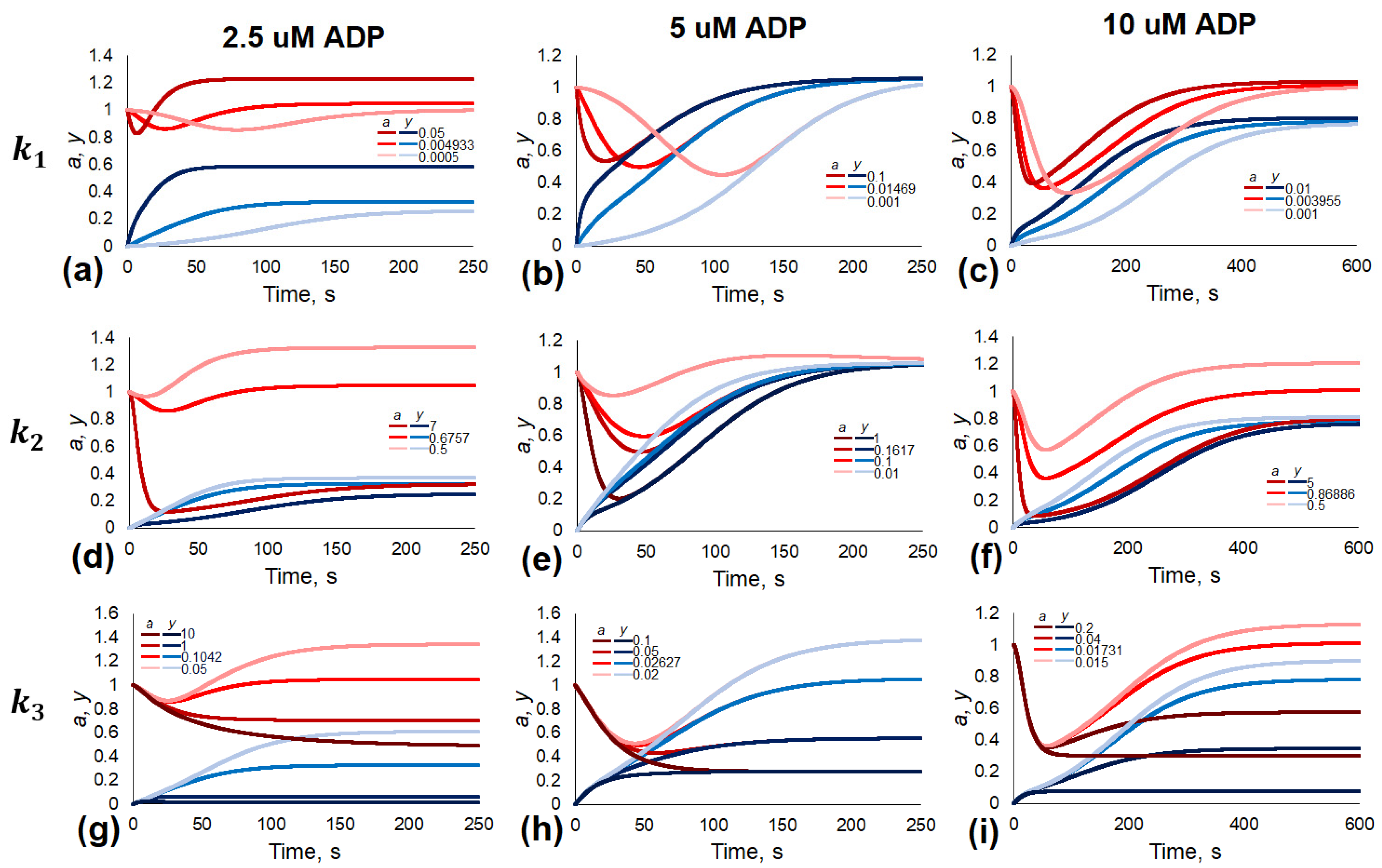
3.3. Effects of Parameter Variation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Varga-Szabo, D.; Pleines, I.; Nieswandt, B. Cell Adhesion Mechanisms in Platelets. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 403–412. [Google Scholar] [CrossRef]
- Filkova, A.A.; Martyanov, A.A.; Dasgupta, A.K.G.; Panteleev, M.A.; Sveshnikova, A.N. Quantitative Dynamics of Reversible Platelet Aggregation: Mathematical Modelling and Experiments. Sci. Rep. 2019, 9, 6217. [Google Scholar] [CrossRef]
- Born, G.V.R. Aggregation of Blood Platelets by Adenosine Diphosphate and Its Reversal. Nature 1962, 194, 927. [Google Scholar] [CrossRef]
- Stalker, T.J.; Traxler, E.A.; Wu, J.; Wannemacher, K.M.; Cermignano, S.L.; Voronov, R.; Diamond, S.L.; Brass, L.F. Hierarchical Organization in the Hemostatic Response and Its Relationship to the Platelet-Signaling Network. Blood 2013, 121, 1875–1885. [Google Scholar] [CrossRef]
- Ling, L.; Liao, J.; Niu, Q.; Wang, X.; Jia, J.; Zuo, C.; Jiang, H.; Zhou, J. Evaluation of an Automated Light Transmission Aggregometry. Platelets 2017, 28, 712–719. [Google Scholar] [CrossRef] [Green Version]
- Kaneva, V.N.; Dunster, J.L.; Volpert, V.; Ataullahanov, F.; Panteleev, M.A.; Nechipurenko, D.Y. Modeling Thrombus Shell: Linking Adhesion Receptor Properties and Macroscopic Dynamics. Biophys. J. 2021, 120, 334–351. [Google Scholar] [CrossRef]
- Gupta, P.; Zhang, P.; Sheriff, J.; Bluestein, D.; Deng, Y. A Multiscale Model for Recruitment Aggregation of Platelets by Correlating with In Vitro Results. Cell. Mol. Bioeng. 2019, 12, 327–343. [Google Scholar] [CrossRef]
- Zheng, X.; Yazdani, A.; Li, H.; Humphrey, J.D.; Karniadakis, G.E. A Three-Dimensional Phase-Field Model for Multiscale Modeling of Thrombus Biomechanics in Blood Vessels. PLoS Comput. Biol. 2020, 16, e1007709. [Google Scholar] [CrossRef]
- Danes, N.A.; Leiderman, K. A Density-dependent FEM-FCT Algorithm with Application to Modeling Platelet Aggregation. Int. J. Numer. Methods Biomed. Eng. 2019, 35. [Google Scholar] [CrossRef]
- Ye, T.; Shi, H.; Phan-Thien, N.; Lim, C.T. The Key Events of Thrombus Formation: Platelet Adhesion and Aggregation. Biomech. Model. Mechanobiol. 2020, 19, 943–955. [Google Scholar] [CrossRef]
- Smoluchowski, M. Attempt for a Mathematical Theory of Kinetic Coagulation of Colloid Solutions. Z. Phys. Chem. 1917, 19, 129–135. [Google Scholar]
- Bentz, J.; Nir, S. Mass Action Kinetics and Equilibria of Reversible Aggregation. J. Chem. Soc. Faraday Trans. 1 Phys. Chem. Condens. Phases 1981. [Google Scholar] [CrossRef]
- Garzon Dasgupta, A.K.; Martyanov, A.A.; Filkova, A.A.; Panteleev, M.A.; Sveshnikova, A.N. Development of a Simple Kinetic Mathematical Model of Aggregation of Particles or Clustering of Receptors. Life 2020, 10, 97. [Google Scholar] [CrossRef] [PubMed]
- Pitt, M.A.; Myung, J.I.; Montenegro, M.; Pooley, J. Measuring Model Flexibility With Parameter Space Partitioning: An Introduction and Application Example. Cogn. Sci. 2008, 32, 1285–1303. [Google Scholar] [CrossRef] [PubMed]
- Reichert, P.; Omlin, M. On the Usefulness of Overparameterized Ecological Models. Ecol. Model. 1997, 95, 289–299. [Google Scholar] [CrossRef]
- Sagawa, S.; Raghunathan, A.; Koh, P.W.; Liang, P. An Investigation of Why Overparameterization Exacerbates Spurious Correlations. arXiv 2020, arXiv:2005.04345. [Google Scholar]
- Kennedy, J.; Eberhart, R. Particle Swarm Optimization. In Proceedings of the IEEE International Conference on Neural Networks, Perth, WA, Australia, 27 November–1 December 1995; Volume IV, pp. 1942–1948. [Google Scholar]
- Hoops, S.; Sahle, S.; Gauges, R.; Lee, C.; Pahle, J.; Simus, N.; Singhal, M.; Xu, L.; Mendes, P.; Kummer, U. COPASI–a COmplex PAthway SImulator. Bioinformatics 2006, 22, 3067–3074. [Google Scholar] [CrossRef] [Green Version]
- Clewley, R. Hybrid Models and Biological Model Reduction with PyDSTool. PLoS Comput. Biol. 2012. [Google Scholar] [CrossRef] [Green Version]
- Volino, P.; Magnenat-Thalmann, N. Comparing Efficiency of Integration Methods for Cloth Simulation. In Proceedings of the Computer Graphics International 2001, Hong Kong, China, 6 July 2001; pp. 265–272. [Google Scholar]
- Lu, M.; Huang, J.; Ruan, S.; Yu, P. Bifurcation Analysis of an SIRS Epidemic Model with a Generalized Nonmonotone and Saturated Incidence Rate. J. Differ. Equ. 2019, 267, 1859–1898. [Google Scholar] [CrossRef]
- Wu, L.-I.; Feng, Z. Homoclinic Bifurcation in an SIQR Model for Childhood Diseases. J. Differ. Equ. 2000, 168, 150–167. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhang, Q.; Zhang, Y. Bifurcations of a Class of Singular Biological Economic Models. Chaos Solitons Fractals 2009, 40, 1309–1318. [Google Scholar] [CrossRef]
- Cai, L.; Chen, G.; Xiao, D. Multiparametric Bifurcations of an Epidemiological Model with Strong Allee Effect. J. Math. Biol. 2013, 67, 185–215. [Google Scholar] [CrossRef]
- Menger, F.M. The Negative Rate Constants of Breslow and Huang. J. Org. Chem. 1991, 56, 6251–6252. [Google Scholar] [CrossRef]
- Proimos, G. Platelet Aggregation Inhibition with Glycoprotein IIb-IIIa Inhibitors. J. Thromb. Thrombolysis 2001, 11, 99–110. [Google Scholar] [CrossRef]
- Peter, K.; Schwarz, M.; Ylänne, J.; Kohler, B.; Moser, M.; Nordt, T.; Salbach, P.; Kübler, W.; Bode, C. Induction of Fibrinogen Binding and Platelet Aggregation as a Potential Intrinsic Property of Various Glycoprotein IIb/IIIa (IIbβ3) Inhibitors. Blood 1998, 92, 3240–3249. [Google Scholar] [CrossRef]
- Mindukshev, I.; Gambaryan, S.; Kehrer, L.; Schuetz, C.; Kobsar, A.; Rukoyatkina, N.; Nikolaev, V.O.; Krivchenko, A.; Watson, S.P.; Walter, U.; et al. Low Angle Light Scattering Analysis: A Novel Quantitative Method for Functional Characterization of Human and Murine Platelet Receptors. Clin. Chem. Lab. Med. 2012, 50. [Google Scholar] [CrossRef] [PubMed]
- Cazenave, J.-P.; Ohlmann, P.; Cassel, D.; Eckly, A.; Hechler, B.; Gachet, C. Preparation of Washed Platelet Suspensions From Human and Rodent Blood. In Platelets and Megakaryocytes; Humana Press: New Jersey, NJ, USA, 2004; Volume 272, pp. 13–28. ISBN 978-1-59259-782-6. [Google Scholar]
- Babakhani, P.; Bridge, J.; Phenrat, T.; Fagerlund, F.; Doong, R.; Whittle, K.R. Comparison of a New Mass-Concentration, Chain-Reaction Model with the Population-Balance Model for Early- and Late-Stage Aggregation of Shattered Graphene Oxide Nanoparticles. Colloids Surf. A Physicochem. Eng. Asp. 2019, 582, 123862. [Google Scholar] [CrossRef]
- Guy, R.D.; Fogelson, A.L. Probabilistic Modeling of Platelet Aggregation: Effects of Activation Time and Receptor Occupancy. J. Theor. Biol. 2002, 219, 33–53. [Google Scholar] [CrossRef]
- Link, K.G.; Sorrells, M.G.; Danes, N.A.; Neeves, K.B.; Leiderman, K.; Fogelson, A.L. A Mathematical Model of Platelet Aggregation in an Extravascular Injury Under Flow. Multiscale Model. Simul. 2020, 18, 1489–1524. [Google Scholar] [CrossRef]
- Liu, Z.L.; Ku, D.N.; Aidun, C.K. Mechanobiology of Shear-Induced Platelet Aggregation Leading to Occlusive Arterial Thrombosis: A Multiscale in Silico Analysis. J. Biomech. 2021, 120, 110349. [Google Scholar] [CrossRef] [PubMed]
- Du, J.; Kim, D.; Alhawael, G.; Ku, D.N.; Fogelson, A.L. Clot Permeability, Agonist Transport, and Platelet Binding Kinetics in Arterial Thrombosis. Biophys. J. 2020, 119, 2102–2115. [Google Scholar] [CrossRef]
- Pothapragada, S. Supercomputer Simulations of Platelet Activation in Blood Plasma at Multiple Scales. In Proceedings of the 2014 International Conference on High Performance Computing & Simulation (HPCS), Bologna, Italy, 21–25 July 2014; pp. 1011–1013. [Google Scholar]
- Van Rooij, B.J.M.; Závodszky, G.; Azizi Tarksalooyeh, V.W.; Hoekstra, A.G. Identifying the Start of a Platelet Aggregate by the Shear Rate and the Cell-Depleted Layer. J. R. Soc. Interface 2019, 16, 20190148. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Sampani, K.; Zheng, X.; Papageorgiou, D.P.; Yazdani, A.; Bernabeu, M.O.; Karniadakis, G.E.; Sun, J.K. Predictive Modelling of Thrombus Formation in Diabetic Retinal Microaneurysms. R. Soc. Open Sci. 2020, 7, 201102. [Google Scholar] [CrossRef] [PubMed]
- Strogatz, S.H. Nonlinear Dynamics and Chaos with Student Solutions Manual: With Applications to Physics, Biology, Chemistry, and Engineering, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2018; ISBN 978-0-429-68015-1. [Google Scholar]
- Bustos, D.M.; Iglesias, A.A. The Kinetic Properties of Liver Glucokinase and Its Function in Glucose Physiology as a Model for the Comprehensive Study of Enzymes’ Kinetic Parameters and Reversible Inhibitors. Biochem. Mol. Biol. Educ. 2000, 28, 332–337. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 2.5 μM 1 | 5 μM | 10 μM | |
|---|---|---|---|
| k−1 | 0.5027 | 1.73 × 10−11 | 0.1995 |
| k−3 | 0.0262 | 0.02778 | 0.01329 |
| k1 | 0.004933 | 0.01469 | 0.003955 |
| k2 | 0.6757 | 0.1617 | 0.86886 |
| k3 | 0.1042 | 0.02627 | 0.01731 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vasilev, G.A.; Filkova, A.A.; Sveshnikova, A.N. Study of Reversible Platelet Aggregation Model by Nonlinear Dynamics. Mathematics 2021, 9, 759. https://doi.org/10.3390/math9070759
Vasilev GA, Filkova AA, Sveshnikova AN. Study of Reversible Platelet Aggregation Model by Nonlinear Dynamics. Mathematics. 2021; 9(7):759. https://doi.org/10.3390/math9070759
Chicago/Turabian StyleVasilev, Grigorii A., Aleksandra A. Filkova, and Anastasia N. Sveshnikova. 2021. "Study of Reversible Platelet Aggregation Model by Nonlinear Dynamics" Mathematics 9, no. 7: 759. https://doi.org/10.3390/math9070759