
Global Stabilization of a Single-Species Ecosystem with Markovian Jumping under Neumann Boundary Value via Laplacian Semigroup


{kind=link}
{kind=link}
Abstract
:1. Introduction
- The uniqueness proof of the positive equilibrium solution is presented in this paper, while it was given in previous work that only involved the existence of the positive equilibrium solution.
- In the case of a single-species ecosystem with impulses, it is the first study using a Laplacian semigroup to globally stabilize the ecosystem.
- A numerical example is designed to illuminate the advantages of Theorem 2 against [22] (Theorem 4.2), as a result of reducing the algorithm’s conservatism.
2. Preparatory Work
3. Main Result
- (A1) g is pth moment continuous at with ;
- (A2) for any given , and exist, and ;
- (A3) ;
- (A4) as , where is a positive scalar with .
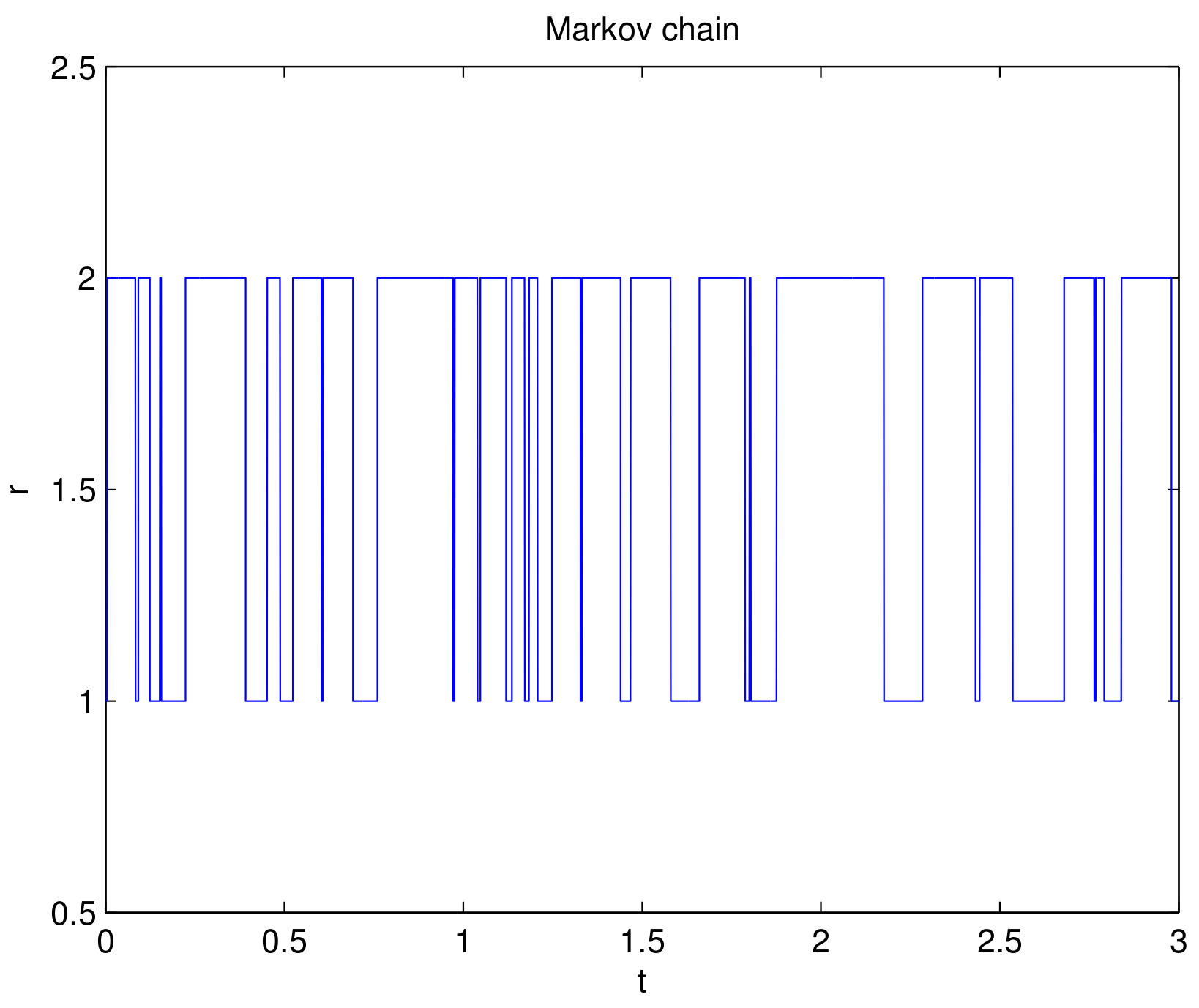
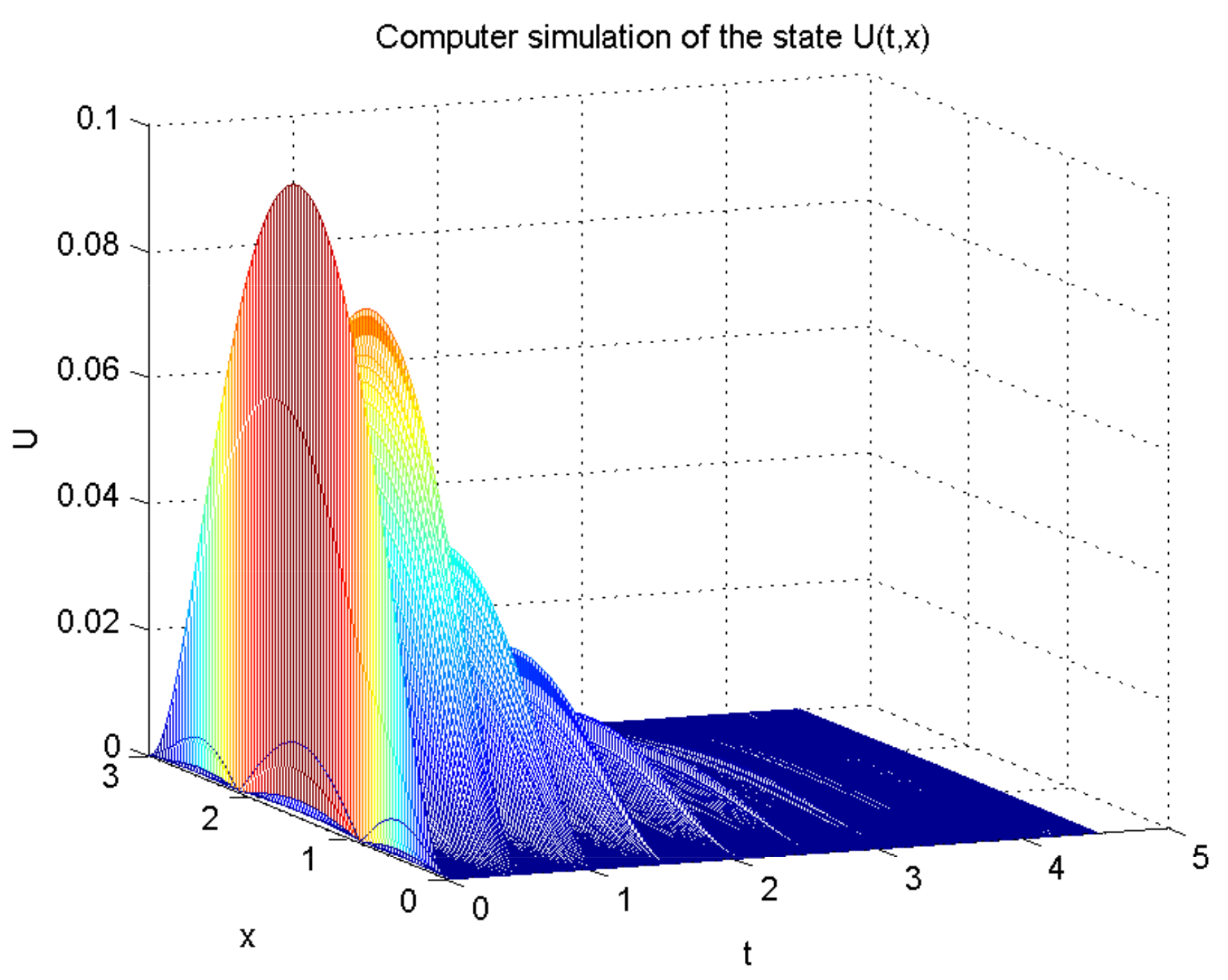
4. Numerical Example
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, L.; Meng, X.; Jiao, J. Biodynamics; Science Press: Beijing, China, 2009. [Google Scholar]
- Zou, X.; Wang, K. A robustness analysis of biological population models with protection zone. Appl. Math. Model. 2011, 35, 5553–5563. [Google Scholar] [CrossRef]
- Ji, W.; Hu, G. Stability and explicit stationary density of a stochastic single-species model. Appl. Math. Comput. 2021, 390, 125593. [Google Scholar] [CrossRef]
- Yu, X.; Yuan, S.; Zhang, T. Persistence and ergodicity of a stochastic single species model with allee effect under regime switching. Comm. Nonlinear Sci. Numer. Simul. 2018, 59, 359–374. [Google Scholar] [CrossRef]
- Jin, Y. Analysis of a stochastic single species model with allee effect and jump-diffusion. Adv. Diff. Equ. 2020, 165, 165. [Google Scholar] [CrossRef]
- Tao, Y.; Winkler, M. Boundedness and stabilization in a population model with cross-diffusion for one species. Proc. London Math. Soc. 2019, 119, 1598–1632. [Google Scholar] [CrossRef]
- Rao, R. Global Stability of Delayed Ecosystem via Impulsive Differential Inequality and Minimax Principle. Mathematics 2021, 9, 1943. [Google Scholar] [CrossRef]
- Rao, R.; Zhu, Q.; Huang, J. Existence, Uniqueness, and Input-to-State Stability of Ground State Stationary Strong Solution of a Single-Species Model via Mountain Pass Lemma. Complexity 2021, 2021, 8855351. [Google Scholar] [CrossRef]
- Borisov, A.; Sokolov, I. Optimal Filtering of Markov Jump Processes Given Observations with State-Dependent Noises: Exact Solution and Stable Numerical Schemes. Mathematics 2020, 8, 506. [Google Scholar] [CrossRef] [Green Version]
- Naranjo, L.; Judith, L.; Esparza, R.; Perez, C.J. A Hidden Markov Model to Address Measurement Errors in Ordinal Response Scale and Non-Decreasing Process. Mathematics 2020, 8, 622. [Google Scholar] [CrossRef] [Green Version]
- Hodara, P.; Papageorgiou, I. Poincare-Type Inequalities for Compact Degenerate Pure Jump Markov Processes. Mathematics 2019, 7, 518. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Chen, W.H.; Ruan, Z.; Huang, T. A new method for global stability analysis of delayed reaction-diffusion neural networks. Neurocomputing 2018, 317, 127–136. [Google Scholar] [CrossRef]
- Rao, R.; Li, X. Input-to-state stability in the meaning of switching for delayed feedback switched stochastic financial system. AIMS Math. 2020, 6, 1040–1064. [Google Scholar] [CrossRef]
- Pan, J.; Liu, X.; Zhong, S. Stability criteria for impulsive reaction-diffusion Cohen-Grossberg neural networks with time-varying delays. Math. Comput. Model. 2010, 51, 1037–1050. [Google Scholar] [CrossRef]
- Feng, Y.; Yang, X.; Song, Q.; Cao, J. Synchronization of memristive neural networks with mixed delays via quantized intermittent control. Appl. Math. Comput. 2018, 339, 874–887. [Google Scholar] [CrossRef]
- Tu, Z.; Wang, D.; Yang, X.; Cao, J. Lagrange stability of memristive quaternion-valued neural networks with neutral items. Neurocomputing 2020, 399, 380–389. [Google Scholar] [CrossRef]
- Song, Q.; Long, L.; Zhao, Z.; Liu, Y.; Alsaadi, F.E. Stability criteria of quaternion-valued neutral-type delayed neural networks. Neurocomputing 2020, 412, 287–294. [Google Scholar] [CrossRef]
- Yang, X.; Song, Q.; Cao, J.; Lu, J. Synchronization of coupled Markovian reaction-diffusion neural networks with proportional delays via quantized control. IEEE Trans. Neu. Net. Learn. Syst. 2019, 3, 951–958. [Google Scholar] [CrossRef]
- Liu, Y.; Wan, X.; Wu, E.; Yang, X.; Alsaadi, F.E.; Hayat, T. Finite-time synchronization of Markovian neural networks with proportional delays and discontinuous activations. Nonlinear Anal. Model. Cont. 2018, 23, 515–532. [Google Scholar] [CrossRef]
- Yang, X.; Feng, Z.; Feng, J.; Cao, J. Synchronization of discrete-time neural networks with delays and Markov jump topologies based on tracker information. Neu. Net. 2017, 85, 157–164. [Google Scholar] [CrossRef]
- Yang, X.; Wan, X.; Cheng, Z.; Cao, J.; Liu, Y.; Rutkowski, L. Synchronization of switched discrete-time neural networks via quantized output control with actuator fault. IEEE Trans. Neu. Net. Learn. Syst. 2021, 32, 4191–4201. [Google Scholar] [CrossRef]
- Rao, R.; Yang, X.; Tang, R.; Zhang, Y.; Li, X. Impulsive stabilization and stability analysis for Gilpin-Ayala competition model involved in harmful species via LMI approach and variational methods. Math. Comput. Simu. 2021, 188, 571–590. [Google Scholar] [CrossRef]
- Winkler, M. Aggregation vs. global diffusive behavior in the higher-dimensional Keller-Segel model. J. Diff. Equ. 2010, 248, 2889–2905. [Google Scholar] [CrossRef] [Green Version]
- Rao, R.; Zhong, S.; Pu, Z. LMI-based robust exponential stability criterion of impulsive integro-differential equations with uncertain parameters via contraction mapping theory. Adv. Diff. Equ. 2017, 2017, 19. [Google Scholar] [CrossRef] [Green Version]
- Huisman, J.; Arrayas, M.; Ebert, U.; Sommeijer, B. How do sinking phytoplankton species manage to persist? Amer. Natur. 2002, 159, 245–254. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- Skellam, J.G. Random dispersal in theoretical populations. Biometrika 1951, 38, 196–218. [Google Scholar] [CrossRef]
- Rao, R.; Huang, J.; Li, X. Stability analysis of nontrivial stationary solution of reaction-diffusion neural networks with time delays under Dirichlet zero boundary value. Neurocomputing 2021, 445C, 105–120. [Google Scholar] [CrossRef]
- Chakraborty, K.; Manthena, V. Modelling and analysis of spatio-temporal dynamics of a marine ecosystem. Nonlinear Dyn. 2015, 81, 1895–1906. [Google Scholar] [CrossRef]
- Kabir, M.H. Reaction-diffusion modeling of the spread of spruce budworm in boreal ecosystem. J. Appl. Math. Comp. 2021, 66, 203–219. [Google Scholar] [CrossRef]
- Bastiaansen, R.; Jaipbi, O.; Deblauwe, V.; Eppinga, M.B.; Siteur, K.; Siero, E.; Mermoz, S.; Bouvet, A.; Doelman, A.; Rietkerk, M. Multistability of model and real dryland ecosystems through spatial self-organization. Proc. Nat. Acad. Sci. USA 2018, 115, 11256–11261. [Google Scholar] [CrossRef] [Green Version]
- Rao, R.; Zhu, Q.; Shi, K. Input-to-State Stability for Impulsive Gilpin-Ayala Competition Model with Reaction Diffusion and Delayed Feedback. IEEE Access 2020, 8, 222625–222634. [Google Scholar] [CrossRef]
- Xiang, H.; Liu, B.; Fang, Z. Optimal control strategies for a new ecosystem governed by reaction-diffusion equations. J. Math. Anal. Appl. 2018, 467, 270–291. [Google Scholar] [CrossRef]
- Rao, R. Impulsive control and global stabilization of reaction-diffusion epidemic model. Math. Meth. Appl. Sci. 2021. [Google Scholar] [CrossRef]
- Rao, R.; Zhong, S.; Pu, Z. Fixed point and p-stability of T-S fuzzy impulsive reaction-diffusion dynamic neural networks with distributed delay via Laplacian semigroup. Neurocomputing 2019, 335, 170–184. [Google Scholar] [CrossRef]
- Istratescu, V.I. Fixed Point Theory: An Introduction; Springer: Dordrecht, The Netherlands, 1981. [Google Scholar]
- Korobenko, L.; Kamrujjaman, M.D.; Braverman, E. Persistence and extinction in spatial models with a carrying capacity driven diffusion and harvesting. J. Math. Anal. Appl. 2013, 399, 352–368. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rao, R.; Huang, J.; Yang, X. Global Stabilization of a Single-Species Ecosystem with Markovian Jumping under Neumann Boundary Value via Laplacian Semigroup. Mathematics 2021, 9, 2446. https://doi.org/10.3390/math9192446
Rao R, Huang J, Yang X. Global Stabilization of a Single-Species Ecosystem with Markovian Jumping under Neumann Boundary Value via Laplacian Semigroup. Mathematics. 2021; 9(19):2446. https://doi.org/10.3390/math9192446
Chicago/Turabian StyleRao, Ruofeng, Jialin Huang, and Xinsong Yang. 2021. "Global Stabilization of a Single-Species Ecosystem with Markovian Jumping under Neumann Boundary Value via Laplacian Semigroup" Mathematics 9, no. 19: 2446. https://doi.org/10.3390/math9192446