
Parameter Estimation for a Kinetic Model of a Cellular System Using Model Order Reduction Method


Abstract
:1. Introduction
2. Parameter Estimation for ODEs Using the POD Method
2.1. Creating the Snapshot Matrix
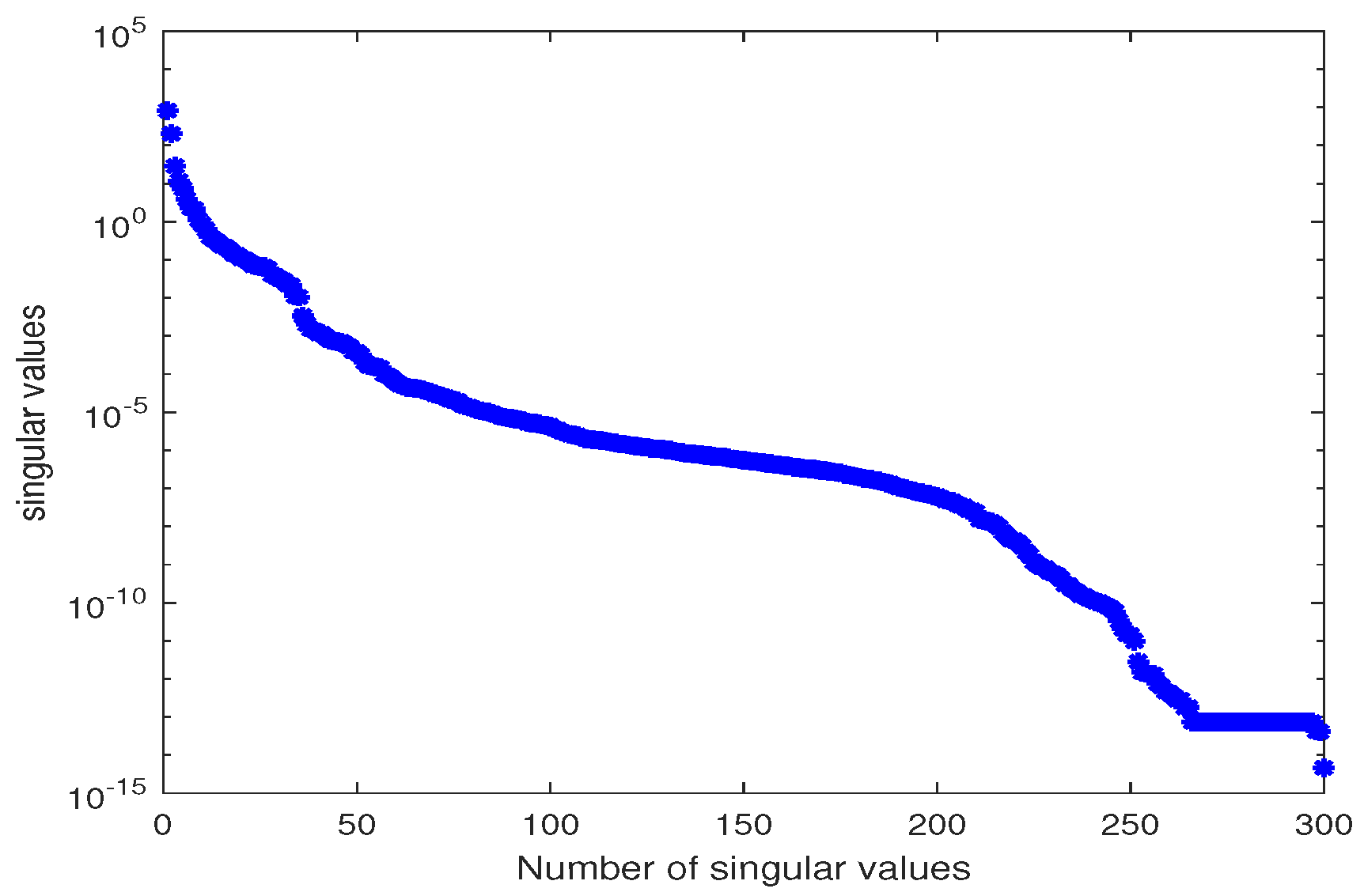
2.2. Computing the Reduced Basis
2.3. Construction of Data-Fitting Function
2.4. Estimating the Parameters
| Algorithm 1 Parameter estimation for a kinetic model. |
|
3. Application of the Parameter Estimation Method to a Kinetic Model of CCR in E. coli
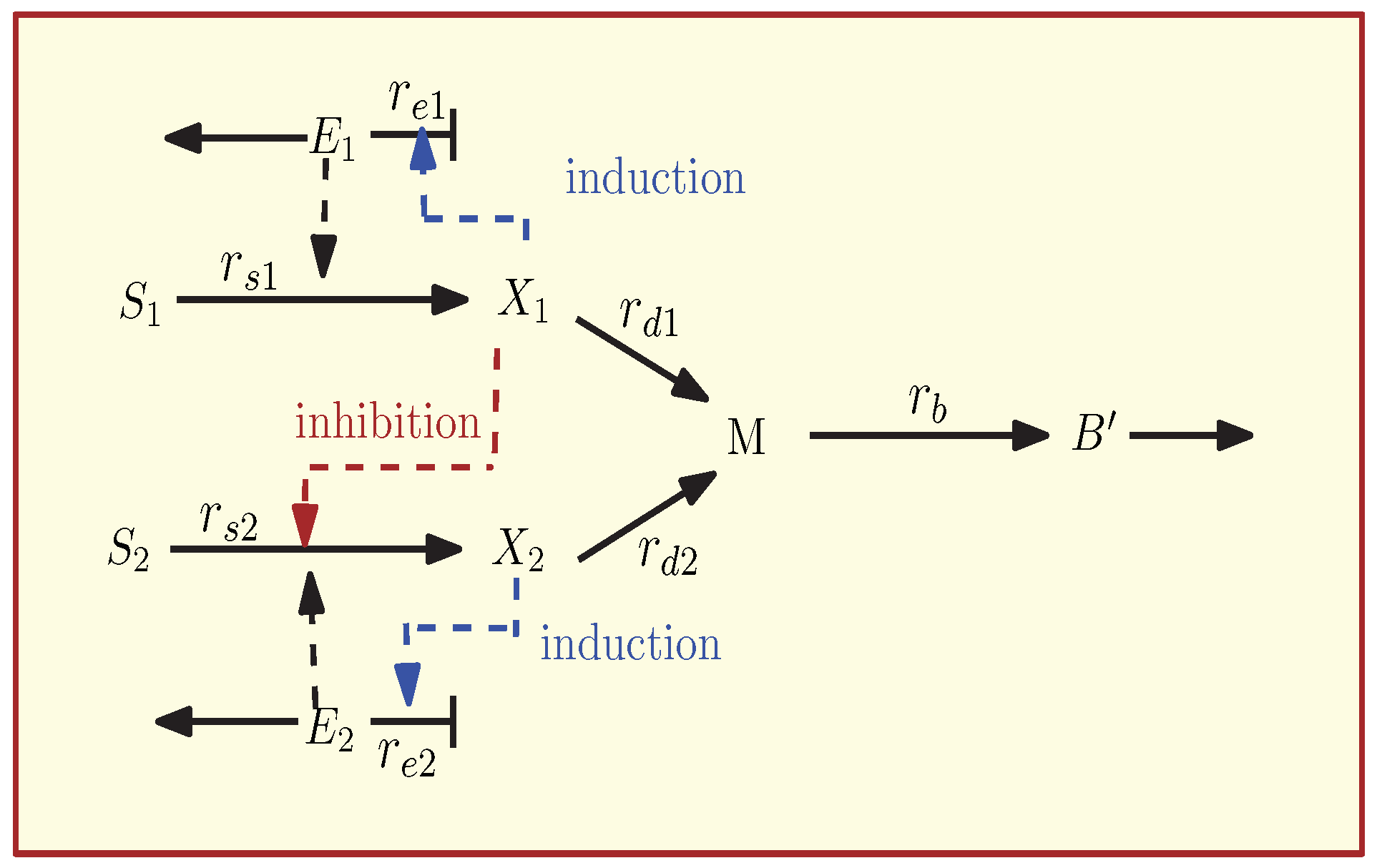
3.1. The Kinetic Model of the Carbon Catabolite Repression
3.2. Numerical Results
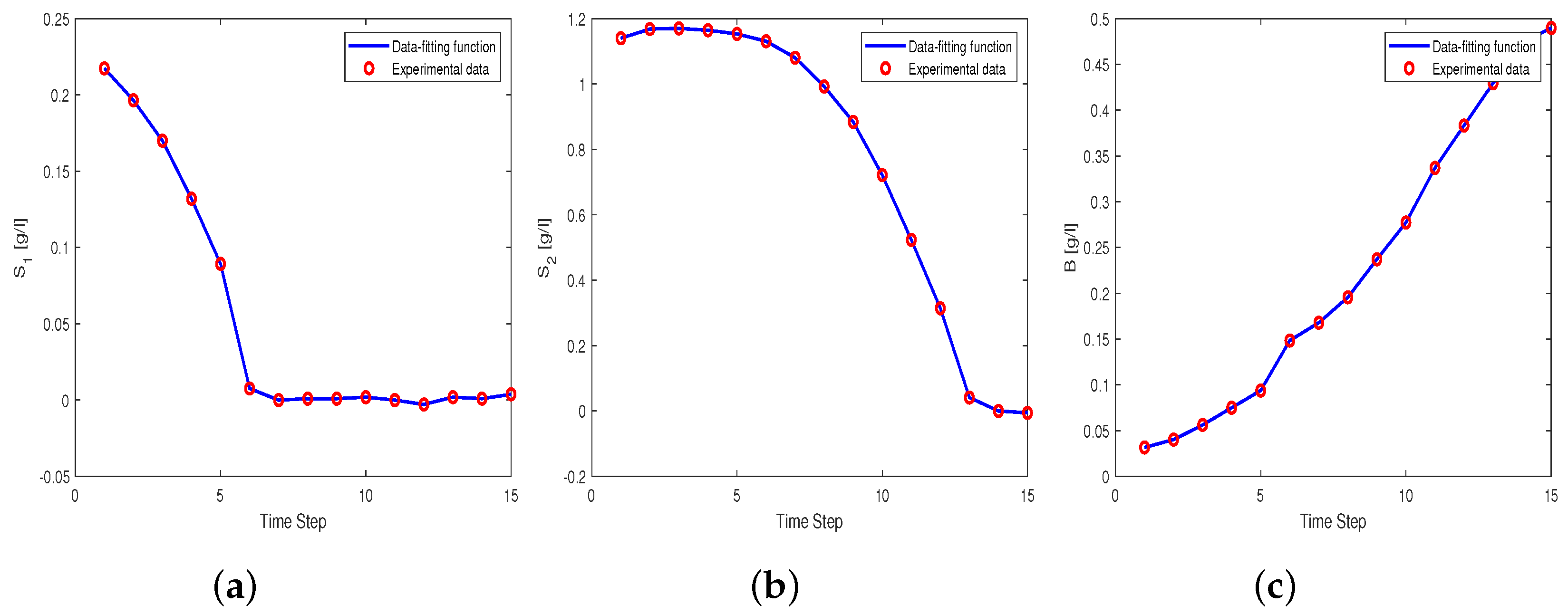
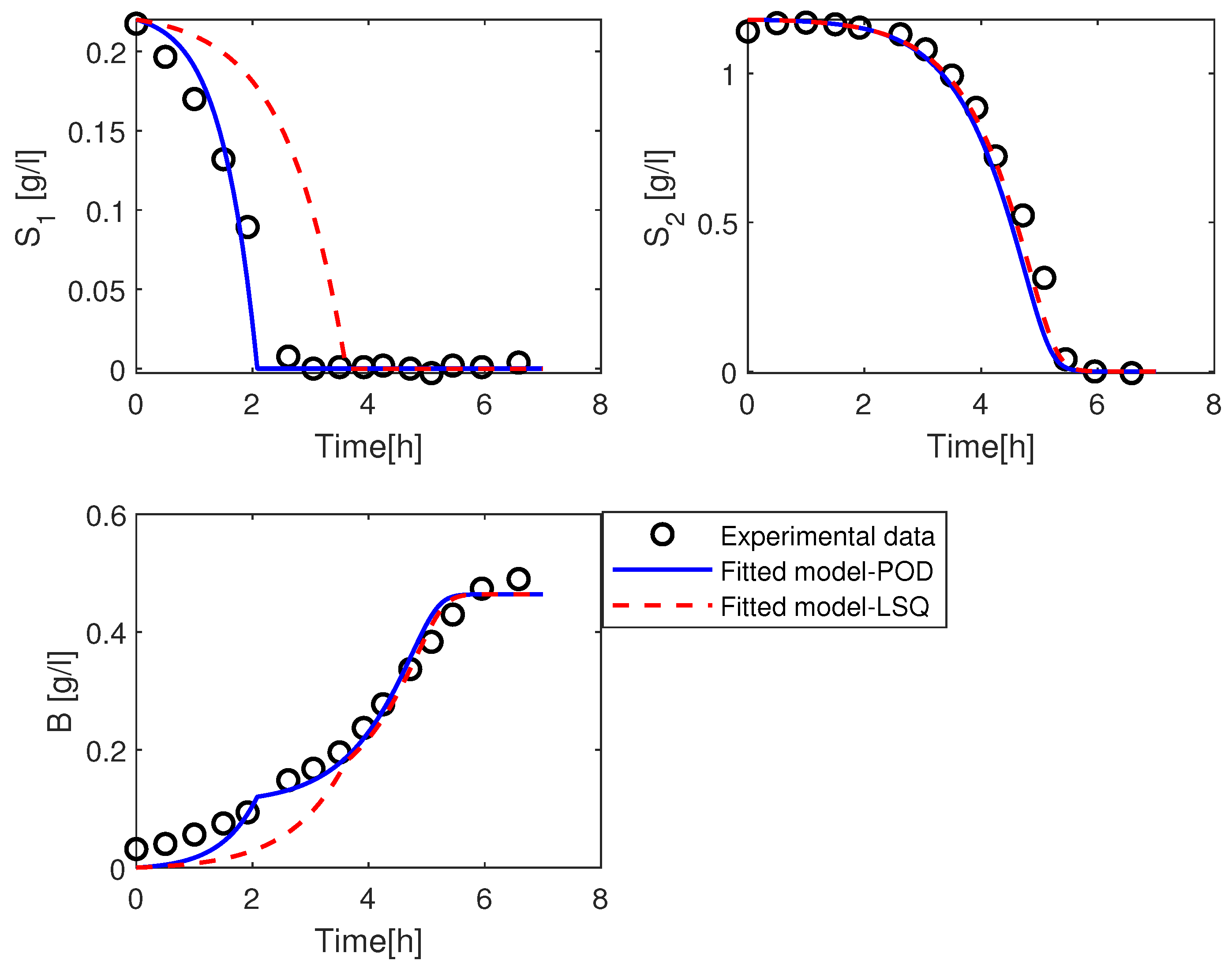
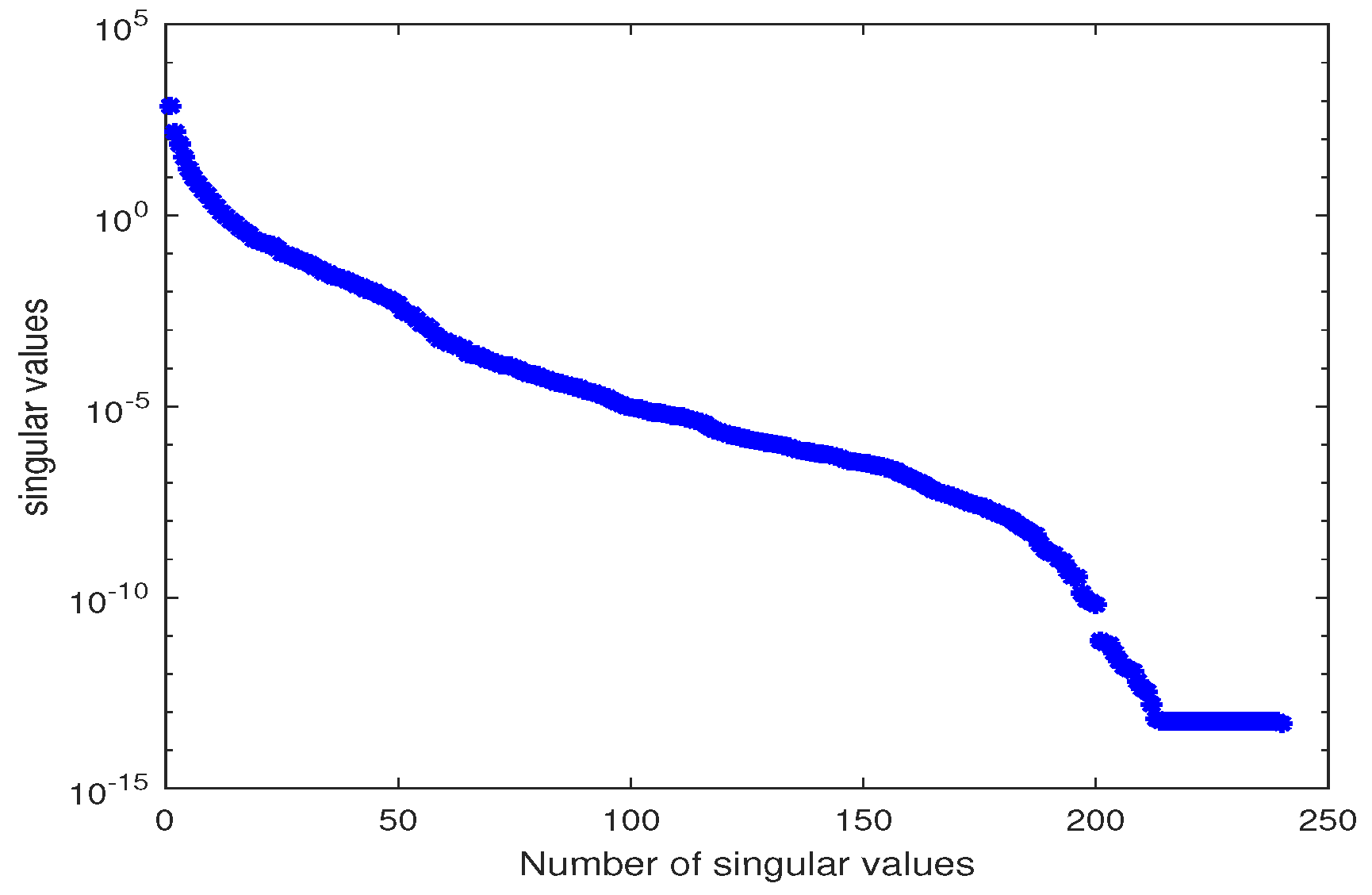
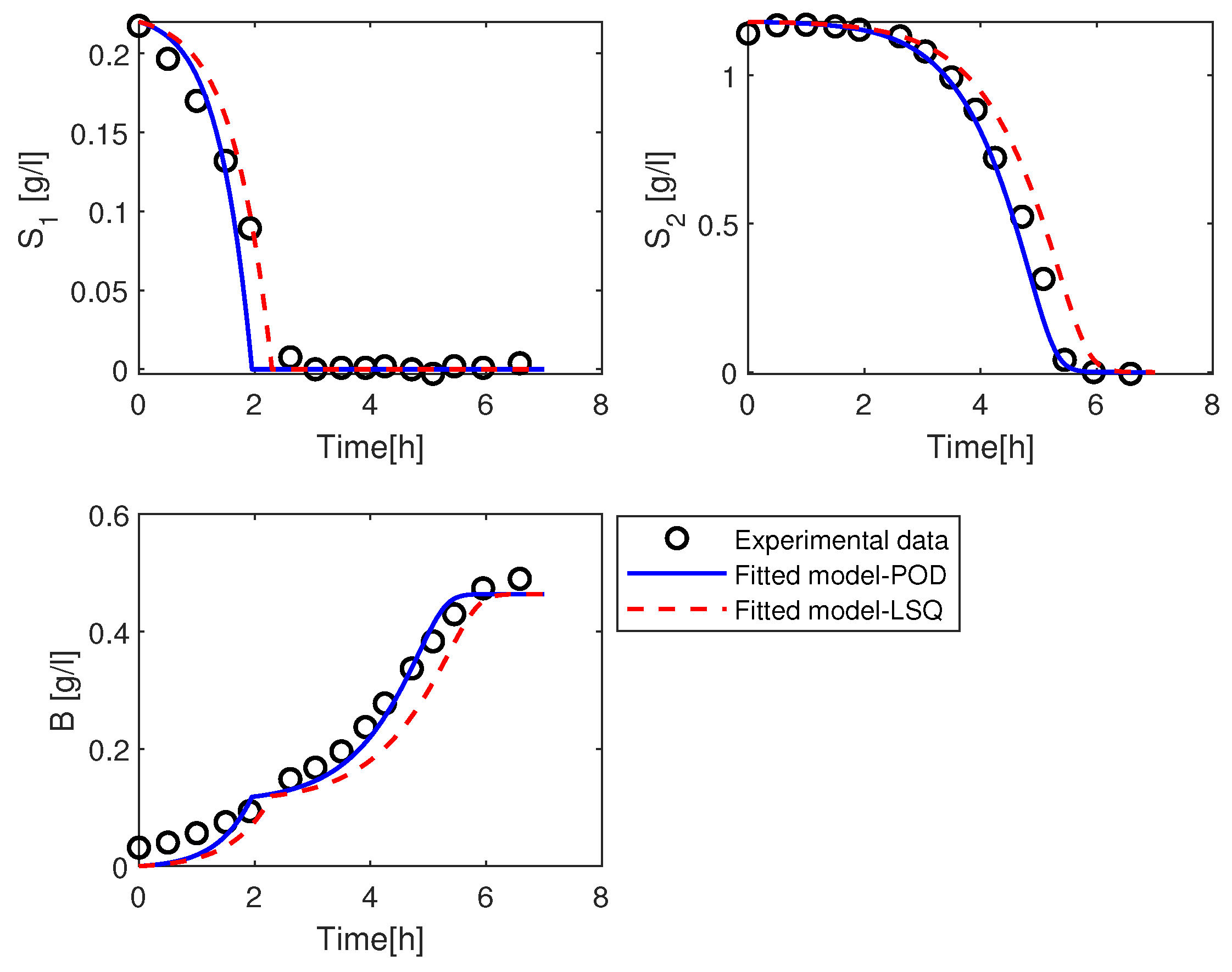
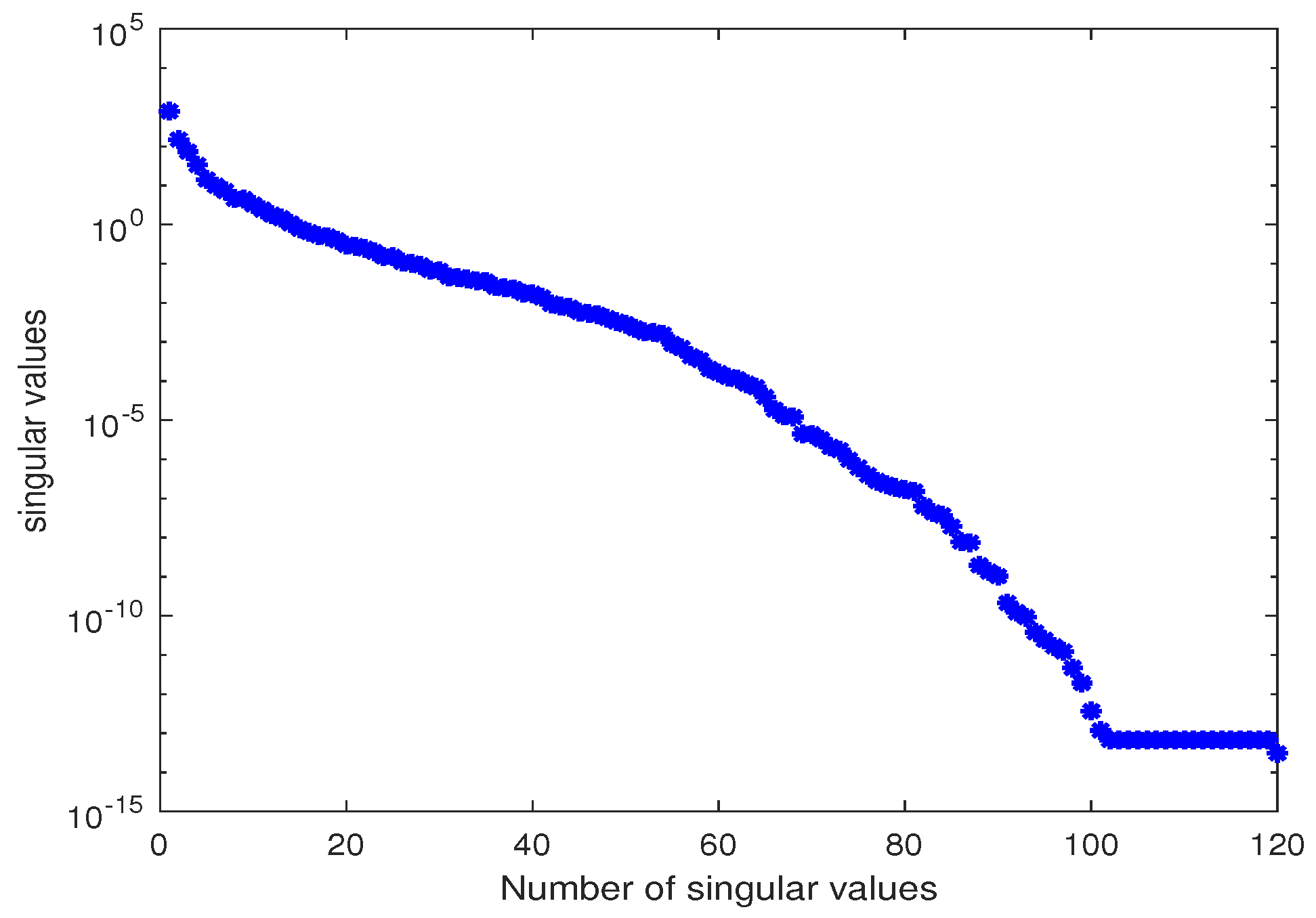
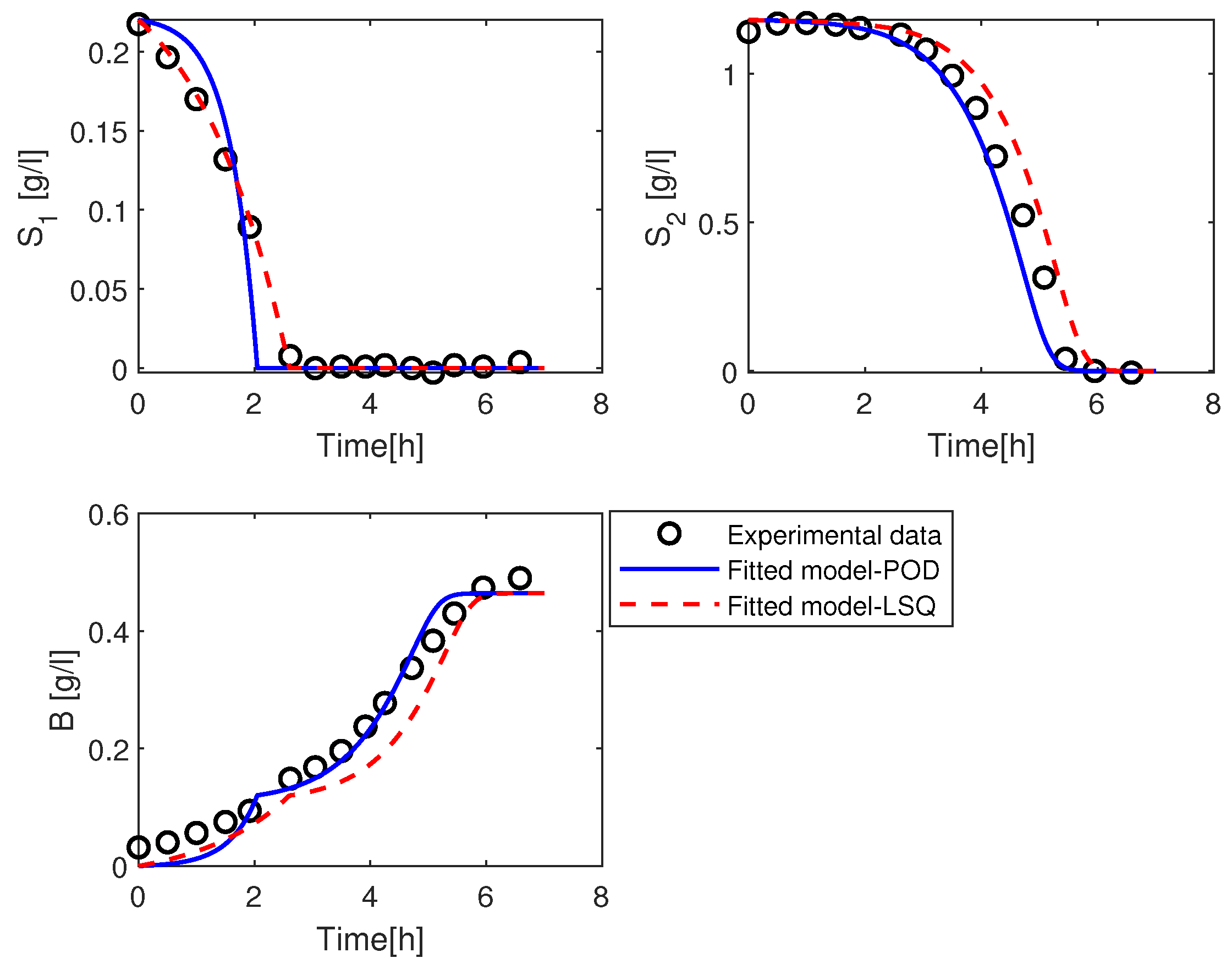
3.2.1. Estimation of the Parameter
3.2.2. Estimation of the Parameters and
3.2.3. Estimation of the Parameters and
4. Conclusions and Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mannan, A.A.; Toya, Y.; Shimizu, K.; McFadden, J.; Kierzek, A.M.; Rocco, A. Integrating kinetic model of E. coli with genome scale metabolic fluxes overcomes its open system problem and reveals bistability in central metabolism. PLoS ONE 2015, 10, e0139507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lima, A.P.; Baixinho, V.; Machado, D.; Rocha, I. A comparative analysis of dynamic models of the central carbon metabolism of Escherichia coli. IFAC-PapersOnLine 2016, 49, 270–276. [Google Scholar] [CrossRef] [Green Version]
- Ali Eshtewy, N. Mathematical Modeling of Metabolic-Genetic Networks; Freie Universität Berlin: Berlin, Germany, 2020. [Google Scholar]
- Aster, R.C.; Borchers, B.; Thurber, C.H. Parameter Estimation and Inverse Problems; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Moles, C.G.; Mendes, P.; Banga, J.R. Parameter estimation in biochemical pathways: A comparison of global optimization methods. Genome Res. 2013, 13, 2467–2474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarantola, A. Inverse Problem Theory and Methods for Model Parameter Estimation; SIAM: Philadelphia, PA, USA, 2005; Volume 89. [Google Scholar]
- Zhdanov, M.S. Inverse Theory and Applications in Geophysics; Elsevier: Amsterdam, The Netherlands, 2015; Volume 36. [Google Scholar]
- Parker, R.L. Understanding inverse theory. Annu. Rev. Earth Planet. Sci. 1977, 5, 35–64. [Google Scholar] [CrossRef]
- Kreutz, C. An easy and efficient approach for testing identifiability. Bioinformatics 2018, 34, 1913–1921. [Google Scholar] [CrossRef] [Green Version]
- Raue, A.; Schilling, M.; Bachmann, J.; Matteson, A.; Schelke, M.; Kaschek, D.; Hug, S.; Kreutz, C.; Harms, B.D.; Theis, F.J.; et al. Lessons Learned from Quantitative Dynamical Modeling in Systems Biology. PLoS ONE 2013, 8, e74335. [Google Scholar] [CrossRef]
- Deuflhard, P. Newton Methods for Nonlinear Problems: Affine Invariance and Adaptive Algorithms; Springer: Berlin/Heidelberg, Germany, 2011; Volume 35. [Google Scholar]
- Altman, N.; Krzywinski, M. Points of Significance: Simple Linear Regression; Nature Publishing Group: London, UK, 2015. [Google Scholar]
- Björck, Å. Numerical Methods for Least Squares Problems; Society for Industrial and Applied Mathematics: Philadelphia, PA, USA, 1996. [Google Scholar]
- Enders, C.K. Maximum likelihood estimation. In Encyclopedia of Statistics in Behavioral Science; Wiley: Hoboken, NJ, USA, 2005. [Google Scholar]
- Rudi, J.; Bessac, J.; Lenzi, A. Parameter Estimation with Dense and Convolutional Neural Networks Applied to the FitzHugh—Nagumo ODE. Math. Sci. Mach. Learn. 2022, 781–808. [Google Scholar] [CrossRef]
- Kerschen, G.; Golinval, J.; Vakakis, A.F.; Bergman, L.A. The method of proper orthogonal decomposition for dynamical characterization and order reduction of mechanical systems: An overview. Nonlinear Dyn. 2005, 41, 147–169. [Google Scholar] [CrossRef]
- Volkwein, S. Proper orthogonal decomposition: Theory and reduced-order modelling. Lect. Notes Univ. Konstanz 2013, 4, 1–29. [Google Scholar]
- Karhunen, K. Über lineare Methoden in der Wahrscheinlichkeitsrechnung. Ann. Acad. Sci. Fenn. Ser. A Math. Phys. 1947, 47, 1–79. [Google Scholar]
- Loeve, M. Elementary Probability Theory; Springer: Berlin/Heidelberg, Germany, 1977; pp. 1–52. [Google Scholar]
- Benner, P.; Goyal, P.; Heiland, J.; Duff, I.P. Operator inference and physics-informed learning of low-dimensional models for incompressible flows. arXiv 2020, arXiv:2010.06701. [Google Scholar] [CrossRef]
- Cazemier, W.; Verstappen, R.W.C.P.; Veldman, A.E.P. Proper orthogonal decomposition and low-dimensional models for driven cavity flows. Phys. Fluids 1998, 10, 1685–1699. [Google Scholar] [CrossRef]
- Luo, Z.; Chen, J.; Navon, I.M.; Yang, X. Mixed finite element formulation and error estimates based on proper orthogonal decomposition for the nonstationary Navier–Stokes equations. SIAM J. Numer. Anal. 2009, 47, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Ali Eshtewy, N.; Scholz, L. Model Reduction for Kinetic Models of Biological Systems. Symmetry 2020, 12, 863. [Google Scholar] [CrossRef]
- Rehm, A.M.; Scribner, E.Y.; Fathallah-Shaykh, H. Proper orthogonal decomposition for parameter estimation in oscillating biological networks. J. Comput. Appl. Math. 2014, 258, 135–150. [Google Scholar] [CrossRef]
- Ly, H.V.; Tran, H.T. Proper orthogonal decomposition for flow calculations and optimal control in a horizontal CVD reactor. Q. Appl. Math. 2002, 60, 631–656. [Google Scholar] [CrossRef] [Green Version]
- Boulakia, M.; Schenone, E.; Gerbeau, J.-F. Reduced-order modeling for cardiac electrophysiology. Application to parameter identification. Int. J. Numer. Methods Biomed. Eng. 2012, 28, 727–744. [Google Scholar] [CrossRef] [Green Version]
- Kahlbacher, M.; Volkwein, S. Estimation of regularization parameters in elliptic optimal control problems by POD model reduction. In Proceedings of the IFIP Conference on System Modeling and Optimization, Cracow, Poland, 23–27 July 2007; Springer: Berlin/Heidelberg, Germany, 2007; pp. 307–318. [Google Scholar]
- Kremling, A.; Geiselmann, J.; Ropers, D.; De Jong, H. An ensemble of mathematical models showing diauxic growth behaviour. BMC Syst. Biol. 2018, 12, 82. [Google Scholar] [CrossRef]
- Helton, J.C.; Davis, F.J. Latin hypercube sampling and the propagation of uncertainty in analyses of complex systems. Reliab. Eng. Syst. Saf. 2003, 81, 23–69. [Google Scholar] [CrossRef] [Green Version]
- McKay, M.D.; Beckman, R.J.; Conover, W.J. A comparison of three methods for selecting values of input variables in the analysis of output from a computer code. Technometrics 2000, 42, 55–61. [Google Scholar] [CrossRef]
- Golub, G.H.; Van Loan, C.F. Matrix Computations, 3rd ed.; Johns Hopkins University Press: Baltimore, MD, USA, 1996. [Google Scholar]
- Hinze, M.; Volkwein, S. Model Order Reduction by Proper Orthogonal Decomposition. Konstanz. Schriften Math. 2019, 47–96. [Google Scholar] [CrossRef]
- Afanasiev, K.; Hinze, M. Adaptive Control of a Wake Flow Using Proper Orthogonal Decomposition. Lect. Notes Pure Appl. Math 2001, 216, 317–332. [Google Scholar]
- Michaelis, L.; Menten, M.L. Die Kinetik der Invertinwirkung. Biochem. Z. 1913, 49, 333–369. [Google Scholar]
- Guldberg, C.M.; Waage, P. Etudes Sur Les Affinités Chimiques; Brøgger & Christie: Christiania, Denmark, 1867. [Google Scholar]
- Per Christian, H. The truncatedSVD as a method for regularization. Bit Numer. Math. 1987, 27, 534–553. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Constant Rates | Value | Unit |
|---|---|---|
| g/L | ||
| g/L | ||
| 6 | mol/gDWh | |
| mol/gDWh | ||
| 10 | 1/h | |
| 10 | 1/h | |
| 10 | 1/h | |
| 1 | mol/gDW | |
| mol/gDW | ||
| mol/gDW | ||
| 90 | gDW/mol | |
| gDW/mol | ||
| 180 | gDW/mol | |
| 342 | gDW/mol |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eshtewy, N.A.; Scholz, L.; Kremling, A. Parameter Estimation for a Kinetic Model of a Cellular System Using Model Order Reduction Method. Mathematics 2023, 11, 699. https://doi.org/10.3390/math11030699
Eshtewy NA, Scholz L, Kremling A. Parameter Estimation for a Kinetic Model of a Cellular System Using Model Order Reduction Method. Mathematics. 2023; 11(3):699. https://doi.org/10.3390/math11030699
Chicago/Turabian StyleEshtewy, Neveen Ali, Lena Scholz, and Andreas Kremling. 2023. "Parameter Estimation for a Kinetic Model of a Cellular System Using Model Order Reduction Method" Mathematics 11, no. 3: 699. https://doi.org/10.3390/math11030699