

The Universal Theory for Multiscale Modelling of Infectious Disease Dynamics

Abstract
:1. Introduction
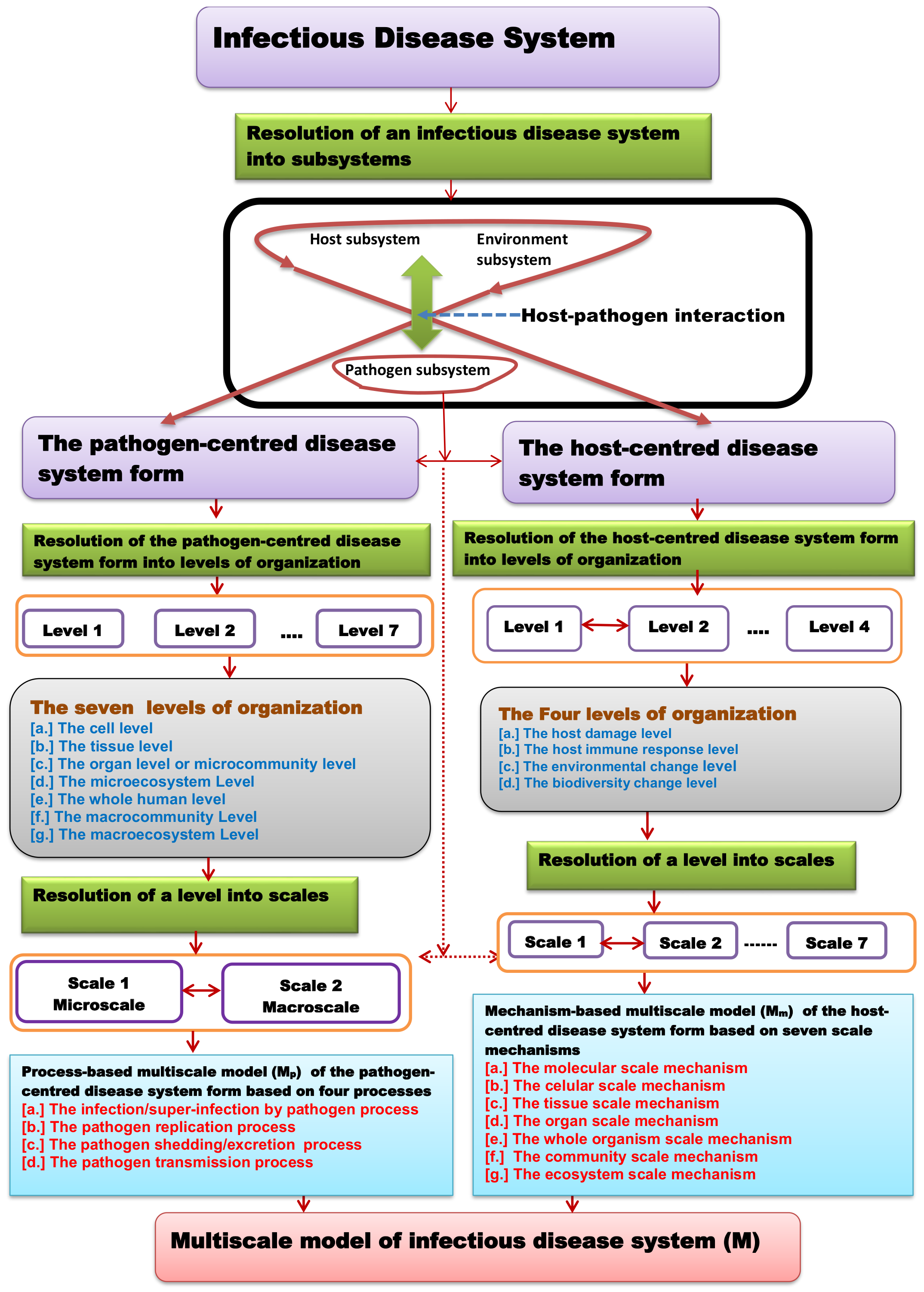
2. Interpretation of the Universal Theory for Multiscale Modelling of Disease Dynamics
- [I.]
- First, that infectious disease dynamics are a result of the interaction of three subsystems, which are the host subsystem, which we call the primary subsystem; the environment subsystem, which we call the secondary subsystem; and the pathogen subsystem, which we call the tertiary subsystem.
- [II.]
- Second, that the environment subsystem is considered to be an extended form of the host subsystem and thus an infectious disease system is considered to be a result of host–pathogen interaction only in such a way that every infectious disease system is considered to exist in two different forms: a host–centred disease system form, which is the form that the whole disease system takes when the host perspective of the interaction is considered, and a pathogen–centred disease system form, which is the form that the whole disease system takes when the pathogen perspective of the interaction is considered.
- [III.]
- Third, that the two different disease system forms; that is, the host-centred disease system form and the pathogen-centred disease system form, have different multilevel and multiscale organisations and are necessarily described by different types of multiscale models, which are process-based multiscale models for the pathogen-centred disease system form and mechanism-based multiscale models for the host-centred disease system form.
- [IV.]
- Fourth, that, together, the mechanism-based multiscale model of the host-centred disease system form and the process-based multiscale model of the pathogen-centred disease system form present a fuller description of the overall multiscale dynamics of an infectious disease system than either of the two when considered separately.
3. The Pathogen-Centred Disease System Form
3.1. The Multilevel Organisation of the Pathogen-Centred Disease System Form
- [a.]
- The cell level: The two hierarchically organised scales at this level of organisation of the pathogen-centred disease system form are the within-cell scale, which is the microscale, and the between-cell scale, which is the macroscale.
- [b.]
- The tissue level: The two hierarchically organised scales at this level of organisation of the pathogen-centred disease system form are the within-tissue scale, which is the microscale, and the between-tissue scale, which is the macroscale.
- [c.]
- The organ level or microcommunity level: For this level of organisation of the pathogen-centred disease system form, the two hierarchically organised scales are the within-organ or within-microcommunity scale, which is the microscale, and the between-organ or between-microcommunity scale, which is the macroscale.
- [d.]
- The microecosystem level: The two hierarchically organised scales at this level of organisation of the pathogen-centred disease system form are the within-microecosystem scale, which is the microscale, and the between-microecosystem scale, which is the macroscale.
- [e.]
- The whole organism level: At this level of organisation of the pathogen-centred disease system form, the two hierarchically organised scales; that is, the microscale and the macroscale, are the within-whole organism scale and the between-whole organism scale, respectively.
- [f.]
- The macrocommunity level: This level of organisation of the pathogen-centred disease system form consists of the within-macrocommunity scale as the microscale and the between-macrocommunity scale as the macroscale.
- [g.]
- The macroecosystem level: The two hierarchically organised scales at this level of organisation of the pathogen-centred disease system form are the within-macroecosystem scale, which is the microscale, and the between-macroecosystem scale, which is the macroscale.
3.2. The Multiscale Organisation of the Pathogen-Centred Disease System Form
- [a.]
- Category I—Individual-Based Multiscale Models (IMSMs): A generic category of multiscale models that provides a simplified means of describing the multiscale dynamics of the pathogen-centred disease system form at a particular level of its organisation. We also sometimes refer to these individual-based multiscale models (IMSMs) as phenomenological-based multiscale models. This is because in this category of multiscale models, only the microscale is explicitly incorporated into the multiscale model and, as a result, the four main disease processes, which include the infection or super-infection by pathogen process, the pathogen replication process, the pathogen shedding or excretion process, and the pathogen transmission process, are only phenomenologically incorporated into the multiscale model with no explicit representation. In these phenomenological-based multiscale models, no macroscale sub-model is considered and so the macroscale is often observed as emergent behaviour of the microscale entities’ behaviour. In order to describe the macroscale, the microscale results are converted by summing, averaging, or performing some detailed statistical analysis of them and aggregating the information into macroscale variables for interpretation at that scale. By failing to explicitly incorporate details of disease processes, phenomenological-based multiscale models only make reference to the complexity of the multiscale dynamics of the pathogen-centred disease system form at a level of its organisation without incorporating the exact content of the complexity. Examples of phenomenological-based multiscale models are agent-based models [4] and graph-theoretic models [5]. Because they implicitly incorporate disease processes of the pathogen-centred disease system form, phenomenological-based multiscale models are typically simpler than the other four categories of multiscale models.
- [b.]
- Category II—Nested Multiscale Models (NMSMs): A generic category of process-based multiscale models developed to characterise the multiscale dynamics of the pathogen-centred disease system form at a particular level of its organisation by incorporating four main disease processes, which include the infection by pathogen process, the pathogen replication process, the pathogen shedding or excretion process, and the pathogen transmission process, across the microscale and the macroscale. In this category of process-based multiscale models, the microscale sub-model and the macroscale sub-model must be described by the same mathematical formalism or mathematical representation and are linked through the exchange of pathogen. For these multiscale models, the macroscale is linked to the microscale through the infection by pathogen process while the microscale is linked to the macroscale through the pathogen shedding or excretion process. A wide range of mathematical techniques can be used to integrate the microscale sub-model and the macroscale sub-model. This gives rise to different classes of process-based multiscale models of the pathogen-centred disease system form in this category of multiscale models. Typical examples of nested multiscale models include [6] for COVID-19 and [7] for paratuberculosis.
- [c.]
- Category III—Embedded Multiscale Models (EMSMs): A generic category of process-based multiscale models developed to characterise the multiscale dynamics of the pathogen-centred disease system form at a particular level of its organisation by incorporating four main disease processes, which include the super-infection by pathogen process, the pathogen replication process, the pathogen shedding or excretion process, and the pathogen transmission process, across the microscale and the macroscale. For this category of process-based multiscale models, the microscale sub-model and the macroscale sub-model must also be described by the same mathematical formalism or mathematical representation and are linked through the exchange of pathogen. Further, the macroscale is linked to the microscale through the super-infection by pathogen process while the microscale is linked to the macroscale through the pathogen shedding or excretion process. A wide range of mathematical techniques can also be used to integrate the microscale sub-model and the macroscale sub-model. This also gives rise to different classes of process-based multiscale models of the pathogen-centred disease system form in this category of multiscale models. Specific examples of embedded multiscale models are [7] for paratuberculosis and [1] for hookworm infection.
- [d.]
- Category IV: Hybrid multiscale models (HMSMs): A generic category of process-based multiscale models developed to characterise the multiscale dynamics of the pathogen-centred disease system form at a particular level of its organisation by incorporating four main disease processes, which include the infection or super-infection by pathogen process, the pathogen replication process, the pathogen shedding or excretion process, and the pathogen transmission process across the microscale and the macroscale. In this category of multiscale models, the microscale sub-model and macroscale sub-model are described by different mathematical formalisms or mathematical representations, depending on differences between scale variables of the microscale and the macroscale, which can be, for example, due to:
- [i.]
- The nature of variation in time scale variables, which can be discrete time at one scale and continuous time at the other scale;
- [ii.]
- The nature of variation in the state variables, which can be stochastic state variables at one scale and deterministic state variables at the other scale;
- [iii.]
- The nature of variation in the spatial scale, which can be homogeneous spatial variables at one scale described by ODEs, and heterogeneous spatial variables at the other scale described by PDEs, etc. In these process-based multiscale models, the macroscale is linked to the microscale through the infection by pathogen process or super-infection by pathogen process while the microscale is linked to the macroscale through the pathogen shedding or excretion process. A wide range of mathematical techniques can also be used to integrate the microscale sub-model and the macroscale sub-model. This gives rise to different classes of process-based multiscale models of the pathogen-centred disease system form in this category of multiscale models. Typical examples of hybrid multiscale models are [8,9,10].
- [e.]
- Category V—Coupled Multiscale Models (CMSMs): A generic category of multiscale models, which may be process-based multiscale models, phenomenological-based multiscale models, or a combination of these two types of multiscale models, that provides a simplified means of describing the multiscale dynamics of a pathogen-centred disease system form at a particular level of its organisation. For this category of multiscale models, the process-based multiscale models from any of categories I, II, III, and IV are used as sub-models in the development of multiscale models of the pathogen-centred disease system form. This is because the multiscale models developed in this category consider multiple pathogen strain infections, and/or multiple pathogen species infections, and/or multiple host group infections, and/or multiple host species infections, and/or multiple communities infections, and/or multiple organ/anatomical compartment infections. They are not like process-based multiscale models of categories I, II, III, and IV, which focus on a specific combination of (i) one-host and (ii) one-pathogen species/strain of a pathogen-centred disease system form relationships in multiscale modelling. Examples of coupled multiscale models are [11] for malaria and [12] for for river blindness, which are described in the context of multiple host species.
4. The Host-Centred Disease System Form
4.1. The Multilevel Organisation of the Host-Centred Disease System Form
- [a.]
- The host damage level: This functional level of organisation of the host-centred disease system form consists of two main complementary level forms: [i.] the pathogen-mediated damage level form and [ii.] the host-mediated damage level form. Usually, the host damage level begins with the pathogen-mediated damage level form, which then triggers the host-mediated damage level form. Currently, host damage can only be quantified in relative terms [13,14]. The term ‘relative’ is necessitated by the fact that, at present, host damage cannot be fully quantified because precise readouts of host damage remain limited and available mathematical modelling tools and computational platforms are insufficient for the quantification of host damage. When host damage surpasses a threshold that maintains host homeostasis, clinical disease occurs [14]. In the multiscale dynamics of the host-centred disease system form, the host damage level triggers the host immune response level [13,14].
- [b.]
- The host immune response level: This functional level of organisation of the host-centred disease system form also consists of two main complementary level forms, which are [i.] the innate immune response level form and [ii.] the adaptive immune response level form. Usually, the host immune response level begins with the innate immune response level form, which then triggers the adaptive host immune response level form. Each of these two complementary level forms of the host immune response level consists of three main scale mechanisms [15,16,17]: [i.] the molecular scale mechanism, which includes the proteome, lipodome, genome, metabolome, transcriptome, and complex molecular processes, such as gene expression, gene regulatory networks, signaling, and metabolic pathways involved in immunity and inflammation; [ii.] the cellular scale mechanism, which includes the activities and behaviour of the different immune cells, such as T-cells, B-cells, and different pathogen processes; and [iii.] the tissue scale mechanism, which includes inflammation mechanisms.
- [c.]
- The environmental change level: In the context of the universal theory for the multiscale modelling of infectious disease dynamics, this level of organisation of the host-centred disease system form arises because the environment subsystem is considered as an extended form of the host subsystem. This functional level of organisation of the host centred disease system form consists of two main complementary levels forms: [i.] the human-induced environmental-change-level form and [ii.] the naturally induced environmental-change-level form [18]. Usually, the environmental change level begins with the human-induced environmental-change-level form through mechanisms such as land use and population growth, which then triggers the naturally-induced environmental-change-level form such as extreme weather events, natural disasters, and climate change. Environmental change can significantly influence infectious disease dynamics through effects such as the survival and reproductive capacity of vectors and pathogens [19]. In the multiscale dynamics of the host-centred disease system form, the environmental change level triggers the biodiversity change level [20].
- [d.]
- The biodiversity change level: In the context of the universal theory for the multiscale modelling of infectious disease dynamics, the term biodiversity or biological diversity refers to the diversity of living organisms implicated in infectious disease dynamics, which are the pathogen subsystem and the host subsystem. This functional level of organisation of the host-centred disease system form also consists of two main complementary level forms: [i.] the pathogen-biodiversity-change-level form, which we also call the microbial-diversity-change-level form or microbiodiversity-change-level form [21], and [ii] the host-biodiversity-change-level form, which we also call the macrobiodiversity-change-level form [22,23]. In multiscale dynamics of the host-centred disease system form, biodiversity change occurs through various evolutionary mechanisms or adaptive mechanisms, which include [i.] mutation mechanisms, and/or [ii.] migration/dispersal mechanisms, and/or [iii.] genetic drift mechanisms, and/or [iv.] natural selection mechanisms. Usually, the biodiversity change level begins with the pathogen-biodiversity-change-level form, which then triggers the host-biodiversity-change-level form, resulting in the reciprocity of change occurring in both the pathogen subsystem and the host subsystem, whereby both the host subsystem and the pathogen subsystem impose evolutionary change on the other in a process involving co-evolutionary mechanisms. In this case, co-evolutionary mechanisms of the host subsystem and pathogen subsystem are a dynamic process of ongoing reciprocal change where a pathogen subsystem imposes an evolutionary influence on a host subsystem, which responds to the evolutionary pressure, in turn imposing an evolutionary influence on the pathogen subsystem, with this cycle potentially repeated over and over again. The outcome of this arms race may involve traits like parasite infectivity, host resistance, parasite host-finding ability, and parasite avoidance behaviour by the host [2]. The multiscale mechanisms of the biodiversity change level of the host-centred disease system form lie at the heart of the emergence and spread of pandemics, resulting in the emergence of new variants at the microbial-diversity-change-level form and new host species at the macrobiodiversity-change-level form.
4.2. The Multiscale Organisation of the Host-Centred Disease System Form
- [a.]
- The molecular-scale mechanism: This involves molecular-scale mechanisms at each of the four main levels of organisation of the host-centred disease system form: the host damage level, the host immune response level, the environmental change level, and the biodiversity change level. Examples of molecular-scale mechanisms include mutation and genetic drift at the biodiversity change level [20], and cytokine, chemokine, and antibody mechanisms at the host immune response level [15,16,17]. Other examples of molecular mechanisms include nutrient cycle mechanisms such as nitrogen cycle and carbon cycle mechanisms at the environmental change level [25].
- [b.]
- The cellular-scale mechanism: This involves cellular=scale mechanisms at each of the four main levels of organisation of the host-centred disease system form: the host damage level, the host immune response level, the environmental change level, and the biodiversity change level. Examples of cellular-scale mechanisms include apoptosis and necrosis mechanisms at the host damage level [13], and T-cell, dendritic cell, and B-cell mechanisms, including macrophage mechanisms such as phagocytosis, at the host immune response level [15,16,17].
- [c.]
- The tissue-scale mechanism: This involves tissue-scale mechanisms at each of the four main levels of organisation of the host-centred disease system form: the host damage level, the host immune response level, the environmental change level, and the biodiversity change level. Examples of tissue-scale mechanisms include inflammation and fibrosis at the host immune response level [26].
- [d.]
- The organ-scale mechanism: This involves organ-scale mechanisms at each of the four main levels of organisation of the host-centred disease system form: the host damage level, the host immune response level, the environmental change level, and the biodiversity change level. Examples of organ-scale mechanisms include sepsis and gangrene formation [27] at the host damage level.
- [e.]
- The whole-organism-scale mechanism: This involves whole-organism-scale mechanisms at each of the four main levels of organisation of the host-centred disease system form: the host damage level, the host immune response level, the environmental change level, and the biodiversity change level. Examples of whole-organism-scale mechanisms include fever mechanisms and behavioural mechanisms such as hydrophobia at the host immune response level [13], and whole organism death at the host damage level.
- [f.]
- The community-scale mechanism: This involves community-scale mechanisms at each of the four main levels of organisation of the host-centred disease system form: the host damage level, the host immune response level, the environmental change level, and the biodiversity change level. Examples of community-scale mechanisms include adaptation and natural selection at the biodiversity change level and population extinction at the host damage level.
- [g.]
- The ecosystem-scale mechanism: This involves ecosystem-scale mechanisms at each of the four main levels of organisation of the host-centred disease system form: the host damage level, the host immune response level, the environmental change level, and the biodiversity change level. Examples of ecosystem-scale mechanisms include predation, competition, and the dilution effect—in which diverse host communities can reduce disease risk [28,29].
5. A Process-Based Multiscale of the Pathogen-Centred Disease System Form of Malaria Disease System as an Example
- [a.]
- At the between-human scale, we have the following three variables: —susceptible human population size; —infected human population size; and —community sporozoite load.
- [b.]
- At the between-mosquito scale, we have the following three variables: —susceptible mosquito population size; —infected mosquito population size; and —community gametocyte load.
- [c.]
- At the within-human scale, we have the following four variables: —susceptible erythrocytes (red blood cells); —merozoite-infected erythrocytes; —merozoites; and —gametocyte-infected erythrocytes.
- [d.]
- At the within-mosquito scale, we have the following five variables: —gametocyte-infected erythrocytes; —sex cells called gametes; —zygotes; —oocysts; and —sporozoites.
6. Characterising the Multiscale Dynamics of the Pathogen-Centred Disease System Form of Malaria Disease System Using the Reproductive Number
7. Characterising the Multiscale Dynamics of the Pathogen-Centred Disease System Form of Malaria Disease System Using Numerical Methods
- ;
- ;
- ;
- ;
- ;
- ;
- ;
- ;
- ;
- ;
- ;
- ;
- ;
- ;
- .
8. Discussion and Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A. Derivation of the Process-Based Multiscale Model of the Pathogen-Centred Disease System Form of Malaria Disease System as an Example
Appendix A.1. Stage I: Single-Scale Sub-models of the Process-Based Multiscale Model of the Pathogen-Centred Disease System Form of Malaria Disease System
- [a.]
- The between-human scale sub-model: The development of a single-scale malaria sub-model at the macroscale of the whole human level of organisation of the pathogen-centred disease system form of a malaria disease system based on [30] involves establishing a new variable called community pathogen load . For the whole human level of organisation, the community pathogen load is the community sporozoite load. This community sporozoite load is a measure of the total infectious reservoir of mosquitoes in the community; that is, an aggregate population-level biomarker of a community’s sporozoite burden over a specific time period [11]. It is a useful metric for assessing the overall impact of malaria health interventions targeted at the mosquito vector or the uptake of malaria interventions targeted at the mosquito vector and quantifying their impact on the transmission of malaria from mosquitoes to humans as [30]: [i.] an indicator of a community’s level of infectiousness and transmission probability of malaria to humans, [ii.] a measure of the effectiveness of malaria interventions targeted at the mosquito vector, and [iii.] a proximal maker of malaria incidence among mosquitoes and their potential to propagate malaria to humans. Then, the single-scale malaria sub-model becomes an SIPS; that is, susceptible human hosts , infected human hosts , community sporozoite load , and susceptible human hosts , as follows:The three variables of the between-human host scale sub-model (A1)vary at slow time scale . The units of the seven between-human host scale sub-model parametersare per annum and have meanings as defined in Table A1. As indicated in [30], the expressionin the sub-model (A1) is the probability that a random bite by a mosquito vector in a particular community with a community sporozoite load will infect the human host with malaria in that community. Since the transmission rate of malaria (A4), which we also refer to as the infectivity response function of malaria, is a probability, it can be modelled by any function with the specification that . Further, since the function is a probability, it must have the following properties [30]:
- [i.]
- Property I: The probability of infection vanishes in the absence of pathogen [i.e., ] and approaches 1 as the community sporozoite load becomes large [i.e., ];
- [ii.]
- Property II: The probability of infection increases with the community sporozoite load ; that is, , where the prime denotes the derivative with respect to the argument.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Description | Parameter Value and Range | Units | Source/Rational |
|---|---|---|---|---|
| Rate of elimination of community sporozoite load | 0.9 [0.09–0.99] | day | Variable | |
| Rate of supply of susceptible mosquitoes | 6000 [5000–7000] | day | Variable | |
| Contact rate of humans with the infectious reservoir of mosquitoes | 0.2 [0.1–0.5] | day | [38] | |
| Half-saturation constant for community gametocyte load | [–] | day | Variable | |
| Natural death rate of mosquitoes | 0.12 [0.033–0.3] | day | [39,40] | |
| Infection-induced death rate of mosquitoes | 0.00000426 [0.00000426–0.00000533] | day | Assumed | |
| Rate of supply of susceptible humans | 1000 [1000–2000] | day | Assumed | |
| Contact rate of mosquitoes with the infectious reservoir of humans | 0.3 [0.1–0.5] | day | [41] | |
| Rate of elimination of community gametocyte load | 0.0000913 [0.000467–0.000274] | day | Variable | |
| Natural death rate of humans | 0.00004 [0.00001–0.00008] | day | [39] | |
| Natural recovery rate of humans | 0.25 [0.1–0.5] | day | Variable | |
| Half-saturation constant for community sporozoite load | [–] | day | Variable | |
| Disease-induced death rate of humans | 0.0027 [0.0001–0.5] | day | Assumed | |
| Proportion of new infected humans in the total infected human population | 0.001 [0.0001–0.5] | Number | Assumed | |
| Proportion of new infected mosquito vectors in the total infected mosquito vector population | 0.002 [0.0001–0.5] | Number | Assumed |
- [b.]
- The between-mosquito scale sub-model: Similarly, following [30], the community pathogen load for the whole mosquito level of organisation of the pathogen-centred disease system form of the malaria disease system is the community gametocyte load. This community gametocyte load is a measure of the total infectious reservoir of humans in the community; that is, an aggregate population-level biomarker of a community’s gametocyte burden over a specific time period [11]. It is a useful metric for assessing the overall impact of malaria health interventions targeted at the human host or the uptake of malaria interventions targeted at the human host and quantifying their impact on the transmission of malaria from humans to mosquitoes as [30]: [i.] an indicator of a community’s level of infectiousness and transmission probability of malaria to mosquitoes, [ii.] a measure of the effectiveness of malaria interventions targeted at the whole human host scale, and [iii.] a proximal maker of malaria incidence among humans and their potential to propagate malaria to mosquito vectors. Then, the single-scale malaria sub-model becomes an SIP; that is, susceptible mosquito hosts , infected mosquito hosts , and community gametocyte load , as follows:The three variables of the between-mosquito host scale sub-model (A5)vary at slow time scale . The units of the six between-mosquito host scale sub-model parametersare per annum and also have meanings as defined in Table A1.Similarly, the expressionin the sub-model (A5) is the probability that a random bite of a human host by a mosquito vector in a particular community with a community gametocyte load will infect the mosquito host with malaria in that community. Since the transmission rate of malaria (A8), which we also refer to as the infectivity response function of malaria, is a probability, it must have the specification that . Equally, since the function is a probability, it must have the following properties [30]:
- [i.]
- Property I: The probability of infection vanishes in the absence of pathogen [i.e., ] and approaches 1 as the community gametocyte load becomes large [i.e., ];
- [ii.]
- Property II: The probability of infection increases with the community gametocyte load ; that is, , where the prime denotes the derivative with respect to the argument.
- [c.]
- The within-human scale sub-model: Following [11], the within-human scale single-scale sub-model with variables —susceptible erythrocytes (red blood cells); —merozoite-infected erythrocytes; —merozoites; and —gametocyte-infected erythrocytes becomes:
| Parameter | Description | Parameter Value and Range | Units | Source/Rational |
|---|---|---|---|---|
| Rate at which gametocyte-infected erythrocytes burst | 96 [90–100] | day | [42] | |
| Rate of uptake of gametocytes through super-infection of mosquito | 300 [100–300] | day | Variable | |
| Death rate of gametocytes | 0.0625 [0.0326–0.0725] | day | [42] | |
| Number of gametes produced per gametocyte-infected erythrocyte | 2 [1–3] | day | Estimated | |
| Fertilisation rate of gametes | 0.08 [0.01–0.2] | noday | [42] | |
| Natural decay rate of gametes | 58.0 [40–129] | day | [42] | |
| Rate at which zygotes develop into oocysts | 0.4240 [0.01–0.05] | day | [43] | |
| Natural decay rate of zygotes | 1 [1–4] | day | [42] | |
| Bursting rate of oocysts to produce sporozoites | 0.2 [0.0–1.0] | day | Variable | |
| Natural decay rate of oocysts | 0.01 [0.071–0.143] | day | [44] | |
| Number of sporozoites produced per bursting oocyst | 3000 [1000–10,000] | day | [42] | |
| Rate at which sporozoites become infectious to humans | 0.025 [0.167–1.00] | day | [42] | |
| Natural decay rate of sporozoites | 0.0001 [0.0001–0.0] | day | [42] | |
| Rate of supply of susceptible red blood cells (erythrocytes) | 200 [100–300] | day | [45] | |
| Infection rate of erythrocytes by free merozoites | 0.1 [–0.2] | day | [39,46] | |
| Natural decay rate of susceptible erythrocytes | 0.0083 [0.006–0.01] | day | [46] | |
| Proportion of gametocyte-infected erythrocytes | 0.1 [0.1–0.5] | Number | Assumed | |
| Natural decay rate of free merozoites | 0.001 [0.001–0.5] | day | [39,46] | |
| Rate at which erythrocytes burst to produce merozoites | 0.5 [0.1–7.0] | day | [46,47] | |
| Number of merozoites produced per bursting erythrocyte | 16 [10–30] | day | [46] | |
| Rate at which gametocytes develop and become infectious | 0.02 [0.01–0.9] | day | [40] | |
| Natural decay rate of gametocyte-infected erythrocytes within infected humans | 0.0625 [0.0600–0.0625] | day | [40] |
- The within-mosquito scale sub-model: Similarly, following [11], the within-mosquito scale sub-model with variables —gametocyte infected erythrocytes; —sex cells called gametes; —zygotes; —oocysts; and —sporozoites becomes:The main difference between the within-mosquito scale sub-model in [11] and the sub-model (A12) is that this sub-model does not incorporate super-infection in the mosquito vector. The five variables of the within-mosquito scale sub-model given by (A12)vary at fast time scale t. The units of the ten within-mosquito scale sub-model parametersare per day and also have meanings as defined in Table A2.
Appendix A.2. Stage II: Integrating the Sub-models into a Single Process-Based Multiscale Model of the Pathogen-Centred Disease System Form of Malaria Disease System of Scale Order Two
Appendix A.3. Stage III: Scale Order Reduction of the Coupled Multiscale Model of the Pathogen-Centred Disease System Form of Malaria Disease System from a Multiscale Model of Scale Order Two to a Multiscale Model of Scale Order One
Appendix A.4. Stage IV: Extending the Coupled Multiscale Model of the Pathogen-Centred Disease System Form of Malaria Disease System by Incorporating Variable Super-Infection in Both the Mosquito Host and the Human Host
- [a.]
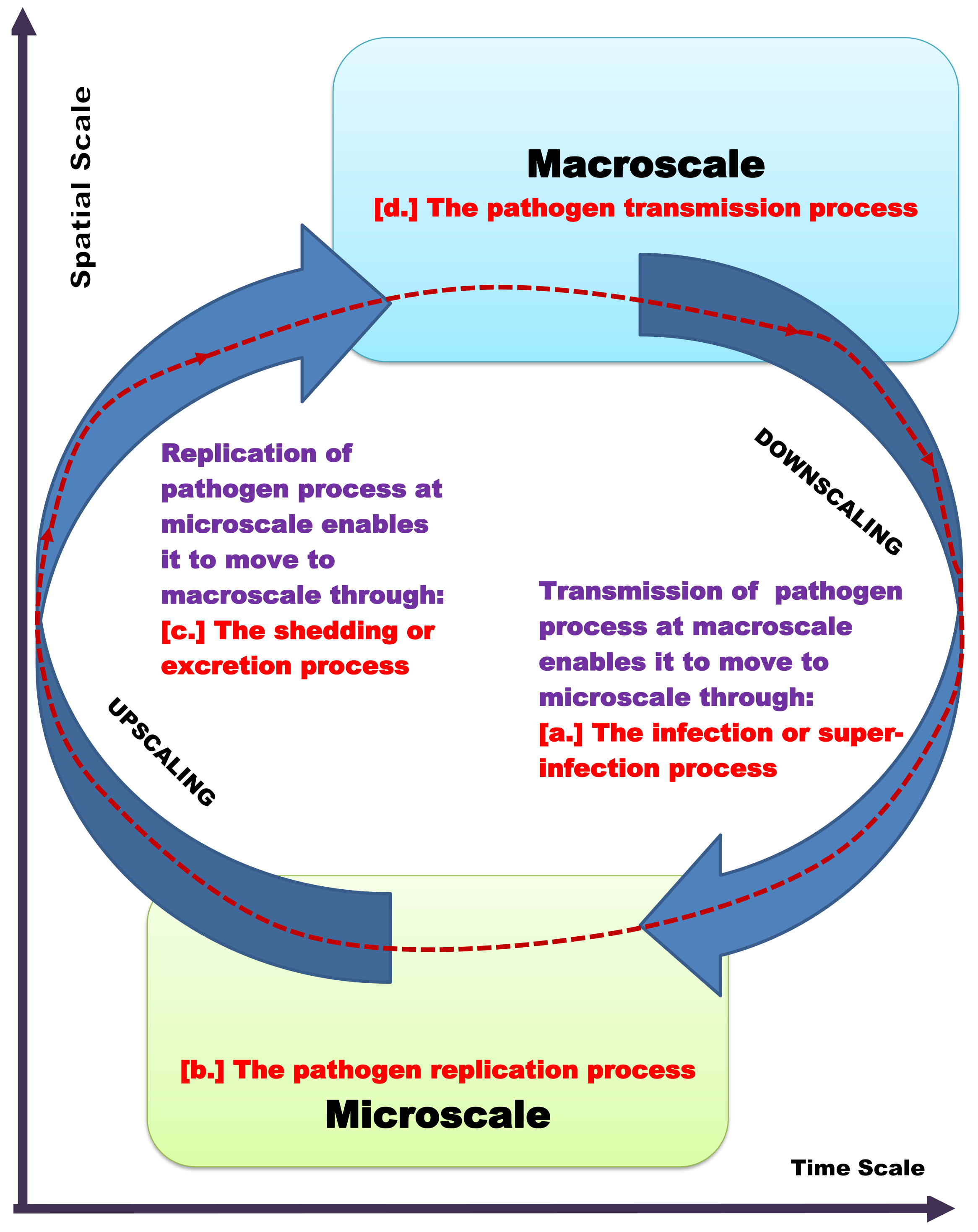
- For the Super-infection of the Human Host: By down-scaling and up-scaling disease process variables and parameters across the microscale and the macroscale as illustrated in Figure 3, we have the following additional expressions:
- [i.]
- Influence of macroscale on microscale: The influence of the between-whole mosquito scale (the macroscale for the whole mosquito level) on the within-whole human scale (the microscale for the whole human level) is modeled by down-scaling the uptake of the pathogen in the community through the infection of humans at the between-human scale at a rate to the repeated infection of the within-human scale at a rate . However, this representation of super-infection is a refinement of the approach in [49] in the following way. The approach in [49] over-estimates the number of new infections, while, here, the number of new infections is assumed to be a proportion of the existing cumulative number of infections.
- [ii.]
- Influence of microscale on macroscale: The influence of the within-whole human scale (the microscale for the whole human level) on the between-whole human scale (the macroscale for the whole human level) is modeled by up-scaling the individual human excretion/shedding of the pathogen at the within-whole human scale at a rate to the between-whole mosquito scale at a rate .
- [b.]
- For the Super-infection of the Mosquito Host: Equally, by down-scaling and up-scaling disease process variables and parameters across the microscale and the macroscale as illustrated in Figure 3, we have the following additional expressions:
- [i.]
- Influence of macroscale on microscale: The influence of the between-whole human scale (the macroscale for the whole human level) on the within-whole mosquito scale (the microscale for the whole mosquito level) is modeled by down-scaling the uptake of the pathogen in the community through the infection of mosquitoes at the between-mosquito scale at a rate to the repeated infection of the within-mosquito scale at a rate . However, this representation of super-infection is also a refinement of the approach in [49] in the following way. The approach in [49] over-estimates the number of new infections, while, here, the number of new infections is assumed to be a proportion of the existing cumulative number of infections.
- [ii.]
- Influence of microscale on macroscale: The influence of the within-whole mosquito scale (the microscale for the whole mosquito level) on the between-whole human scale (the macroscale for the whole human level) is modeled by up-scaling the individual mosquito excretion/shedding of the pathogen at the within-whole mosquito scale at a rate to the between-whole human scale at a rate .
References
- Garira, W. The Replication-Transmission Relativity theory for Multiscale Modelling of infectious Disease Systems. Sci. Rep. 2019, 9, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Garira, W. A primer on multiscale modelling of infectious disease systems. Infect. Dis. Model. 2018, 3, 176–191. [Google Scholar] [CrossRef] [PubMed]
- Garira, W. The research and development process for multiscale models of infectious disease systems. PLoS Comput. Biol. 2020, 16, e1007734. [Google Scholar] [CrossRef] [PubMed]
- Hunter, E.; Mac Namee, B.; Kelleher, J.D. A Taxonomy for Agent-Based Models in Human Infectious Disease Epidemiology. J. Artif. Soc. Soc. Simul. 2017, 20, 1–2. [Google Scholar] [CrossRef]
- Keeling, M.J.; Eames, K.T. Networks and epidemic models. J. R. Soc. Interface 2005, 2, 295–307. [Google Scholar] [CrossRef]
- Chhetri, B.; Vamsi, D.K.; Sanjeevi, C. A Nested Multi-Scale Model for COVID-19 Viral Infection. arXiv 2021, arXiv:2108.12150. [Google Scholar]
- Netshikweta, R.; Garira, W. A nested multiscale model to study paratuberculosis in ruminants. Front. Appl. Math. Stat. 2022, 79, 8762. [Google Scholar] [CrossRef]
- Mideo, N.; Alizon, S.; Day, T. Linking within-and between-host dynamics in the evolutionary epidemiology of infectious diseases. Trends Ecol. Evol. 2008, 23, 511–517. [Google Scholar] [CrossRef]
- Quintela, B.M.; Conway, J.M.; Hyman, J.M.; Guedj, J.; Dos Santos, R.W.; Lobosco, M.; Perelson, A.S. A New Age-Structured Multiscale Model of the Hepatitis C Virus Life-Cycle During Infection and Therapy with Direct-Acting Antiviral Agents. Front. Microbiol. 2018, 9, 601. [Google Scholar] [CrossRef]
- Zitzmann, C.; Kaderali, L. Mathematical Analysis of Viral Replication Dynamics and Antiviral Treatment Strategies: From Basic Models to Age-Based Multi-Scale Modeling. Front. Microbiol. 2018, 9, 1546. [Google Scholar] [CrossRef]
- Garira, W.; Mathebula, D. Development and application of multiscale models of acute viral infections in intervention research. Math. Methods Appl. Sci. 2020, 43, 3280–3306. [Google Scholar] [CrossRef]
- Garira, W.; Chirove, F. A general method for multiscale modelling of vector-borne disease systems. Interface Focus 2020, 10, 20190047. [Google Scholar] [CrossRef]
- Casadevall, A.; Pirofski, L.A. The damage-response framework of microbial pathogenesis. Nat. Rev. Microbiol. 2003, 1, 17–24. [Google Scholar] [CrossRef]
- Pirofski, L.A.; Casadevall, A. The damage-response framework of microbial pathogenesis and infectious diseases. GI Microbiota Regul. Immune Syst. 2008, 4, 135–146. [Google Scholar]
- Waites, W.; Cavaliere, M.; Danos, V.; Datta, R.; Eggo, R.M.; Hallett, T.B.; Manheim, S.; Griffiths, J.P.; Russell, T.; Zarnitsyna, V.I. Compositional modelling of immune response and virus transmission dynamics. Philos. Trans. R. Soc. A 2022, 380, 20210307. [Google Scholar] [CrossRef] [PubMed]
- Holding, T.; Valletta, J.J.; Recker, M. Multiscale immune selection and the transmission-diversity feedback in antigenically diverse pathogen systems. Am. Nat. 2018, 192, E189–E201. [Google Scholar] [CrossRef] [PubMed]
- Cappuccio, A.; Tieri, P.; Castiglione, F. Multiscale modelling in immunology: A review. Brief. Bioinform. 2016, 17, 408–418. [Google Scholar] [CrossRef] [PubMed]
- Budria, A.; Candolin, U. How does human-induced environmental change influence host-parasite interactions? Parasitology 2014, 141, 462–474. [Google Scholar] [CrossRef]
- Johnson, P.T.; Wood, C.L.; Joseph, M.B.; Preston, D.L.; Haas, S.E.; Springer, Y.P. Habitat heterogeneity drives the host-diversity-begets-parasite-diversity relationship: Evidence from experimental and field studies. Ecol. Lett. 2016, 19, 752–761. [Google Scholar] [CrossRef]
- Brunner, F.S.; Eizaguirre, C. Can environmental change affect host/parasite-mediated speciation? Zoology 2016, 119, 384–394. [Google Scholar] [CrossRef]
- Gudelj, I.; Weitz, J.S.; Ferenci, T.; Claire Horner-Devine, M.; Marx, C.J.; Meyer, J.R.; Forde, S.E. An integrative approach to understanding microbial diversity: From intracellular mechanisms to community structure. Ecol. Lett. 2010, 13, 1073–1084. [Google Scholar] [CrossRef] [PubMed]
- Ostfeld, R.S.; Keesing, F. Effects of host diversity on infectious disease. Annu. Rev. Ecol. Evol. Syst. 2012, 43, 2012. [Google Scholar] [CrossRef]
- Johnson, P.T.; Preston, D.L.; Hoverman, J.T.; LaFonte, B.E. Host and parasite diversity jointly control disease risk in complex communities. Proc. Natl. Acad. Sci. USA 2013, 110, 16916–16921. [Google Scholar] [CrossRef] [PubMed]
- Halliday, F.W.; Rohr, J.R. Measuring the shape of the biodiversity-disease relationship across systems reveals new findings and key gaps. Nat. Commun. 2019, 10, 5032. [Google Scholar] [CrossRef] [PubMed]
- Johnson, P.T.; Townsend, A.R.; Cleveland, C.C.; Glibert, P.M.; Howarth, R.W.; McKenzie, V.J.; Rejmankova, P.; Ward, M.H. Linking environmental nutrient enrichment and disease emergence in humans and wildlife. Ecol. Appl. 2010, 20, 16–29. [Google Scholar] [CrossRef]
- Gangell, C.; Gard, S.; Douglas, T.; Park, J.; De Klerk, N.; Keil, T.; Brennan, A.; Ranganathan, S.; Browne, R.; Sly, P.D. Inflammatory responses to individual microorganisms in the lungs of children with cystic fibrosis. Clin. Infect. Dis. 2011, 53, 425–432. [Google Scholar] [CrossRef]
- Jiang, J.L.; Tseng, L.W.; Chang, H.R. Symmetrical peripheral gangrene in sepsis after treatment with inotropes. Tzu-Chi Med. J. 2017, 29, 121. [Google Scholar] [CrossRef]
- Luis, A.D.; Kuenzi, A.J.; Mills, J.N. Species diversity concurrently dilutes and amplifies transmission in a zoonotic host–pathogen system through competing mechanisms. Proc. Natl. Acad. Sci. USA 2018, 115, 7979–7984. [Google Scholar] [CrossRef]
- Halliday, F.W.; Rohr, J.R.; Laine, A.L. Biodiversity loss underlies the dilution effect of biodiversity. Ecol. Lett. 2020, 23, 1611–1622. [Google Scholar] [CrossRef]
- Garira, W.; Maregere, B. The transmission mechanism theory of disease dynamics: Its aims, assumptions and limitations. Infect. Dis. Model. 2023, 8, 122. [Google Scholar] [CrossRef]
- Castillo-Chavez, C.; Feng, Z.; Huang, W. On the computation of R0 and its role in global stability. In Mathematical Approaches for Emerging and Re-Emerging Infectious Diseases Part 1: An Introduction to Models, Methods and Theory; Castillo-Chavez, C., Blower, S., Driessche, P.V., Kirschner, D., Eds.; The IMA Volumes in Mathematics and Its Applications; Springer: Berlin/Heidelberg, Germany, 2002; Volume 125, pp. 229–250. [Google Scholar]
- Mickens, R.E. Nonstandard Finite Difference Models of Differential Equations; World Scientific: Singapore, 1994. [Google Scholar]
- Mickens, R.E. Advances in the Applications of Nonstandard Finite Difference Schemes; World Scientific: Singapore, 2005. [Google Scholar]
- Mickens, R.E. Nonstandard finite difference schemes for differential equations. J. Differ. Equ. Appl. 2002, 8, 823–847. [Google Scholar] [CrossRef]
- Mickens, R.E. Dynamic consistency: A fundamental principle for constructing nonstandard finite difference schemes for differential equations. J. Differ. Equ. Appl. 2005, 11, 645–653. [Google Scholar] [CrossRef]
- Garira, W. A complete categorization of multiscale models of infectious disease systems. J. Biol. Dyn. 2017, 11, 378–435. [Google Scholar] [CrossRef]
- Netshikweta, R.; Garira, W. An Embedded Multiscale Modelling to Guide Control and Elimination of Paratuberculosis in Ruminants. Comput. Math. Methods Med. 2021, 2021, 9919700. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, L.L.M.; Whitehead, S.A.; Thomas, M.B. Quantifying the effects of temperature on mosquito and parasite traits that determine the transmission potential of human malaria. PLoS Biol. 2017, 15, e2003489. [Google Scholar] [CrossRef] [PubMed]
- Cai, L.; Tuncer, N.; Martcheva, M. How does within-host dynamics affect population level dynamics? Insights from an immuno-epidemiological model of malaria. Math. Methods Appl. Sci. 2017, 40, 6424–6450. [Google Scholar] [CrossRef]
- Legros, M.; Bonhoeffer, S. A combined within-host and between-hosts modelling framework for the evolution of resistance to antimalarial drugs. J. R. Soc. Interface 2016, 13, 148. [Google Scholar] [CrossRef]
- Smith, D.L.; McKenzie, F.E. Statics and dynamics of malaria infection in Anopheles mosquitoes. Malar. J. 2004, 3, 1–14. [Google Scholar] [CrossRef]
- Teboh-Ewungkem, M.I.; Yuster, T. A within-vector mathematical model of Plasmodium falciparum and implications of incomplete fertilization on optimal gametocyte sex ratio. J. Theor. Biol. 2010, 264, 273–286. [Google Scholar] [CrossRef]
- Gouagna, L.C.; Bonnet, S.; Gounoue, R.; Verhave, J.P.; Eling, W.; Sauerwein, R.; Boudin, C. Stage-specific effects of host plasma factors on the early sporogony of autologous Plasmodium falciparum isolates within Anopheles gambiae. Trop. Med. Int. Health 2004, 9, 937–948. [Google Scholar] [CrossRef]
- Howells, R.E.; Davies, E.E. Nuclear division in the oocyst of Plasmodium berghei. Ann. Trop. Med. Parasitol. 1971, 65, 451–459. [Google Scholar] [CrossRef] [PubMed]
- Tabo, Z.; Luboobi, L.S.; Ssebuliba, J. Mathematical modelling of the in-host dynamics of malaria and the effects of treatment. J. Math. Comput. Sci. 2017, 17, 1–21. [Google Scholar] [CrossRef]
- Hellriegel, B. Modelling the immune response to malaria with ecological concepts: Short-term behaviour against long-term equilibrium. Proc. R. Soc. Lond. B 1992, 250, 249–256. [Google Scholar]
- Chiyaka, C.; Garira, W.; Dube, S. Modelling immune response and drug therapy in human malaria infection. Comput. Math. Methods Med. 2008, 9, 143–163. [Google Scholar] [CrossRef]
- Holmes, M.H. Introduction to Perturbation Methods; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; Volume 20. [Google Scholar]
- Garira, W.; Mathebula, D.; Netshikweta, R. A mathematical modelling framework for linked within-host and between-host dynamics for infections with free-living pathogens in the environment. Math. Biosci. 2014, 256, 58–78. [Google Scholar] [CrossRef]












Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garira, W.; Muzhinji, K. The Universal Theory for Multiscale Modelling of Infectious Disease Dynamics. Mathematics 2023, 11, 3874. https://doi.org/10.3390/math11183874
Garira W, Muzhinji K. The Universal Theory for Multiscale Modelling of Infectious Disease Dynamics. Mathematics. 2023; 11(18):3874. https://doi.org/10.3390/math11183874
Chicago/Turabian StyleGarira, Winston, and Kizito Muzhinji. 2023. "The Universal Theory for Multiscale Modelling of Infectious Disease Dynamics" Mathematics 11, no. 18: 3874. https://doi.org/10.3390/math11183874