

Mathematical Analysis of an Anthroponotic Cutaneous Leishmaniasis Model with Asymptomatic Infection

Abstract
:1. Introduction
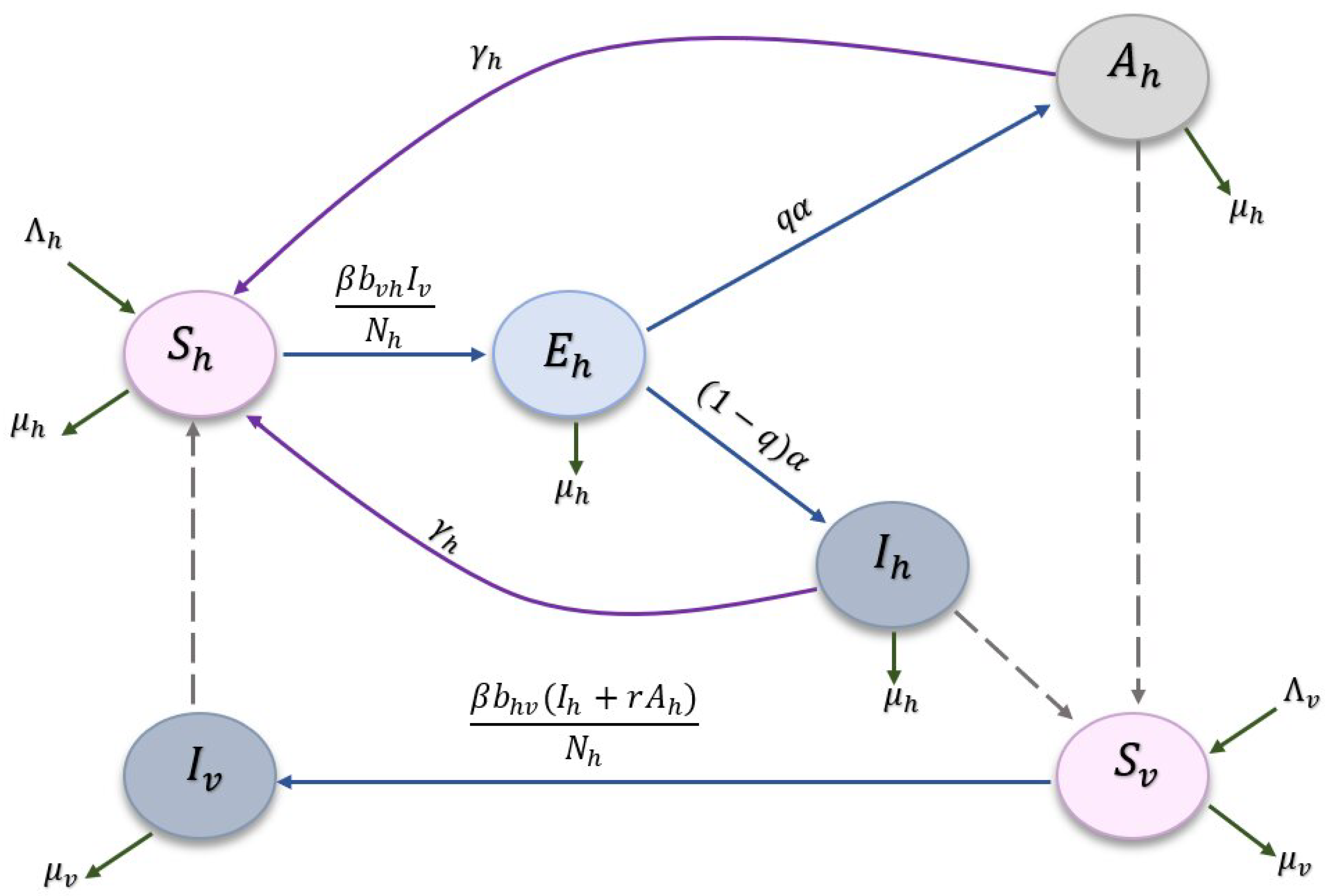
2. Model Formulation and Its Basic Properties
3. Equilibrium Analysis and the Basic Reproduction Number
3.1. CL-Free Equilibrium
3.2. The Basic Reproduction Number
3.3. Endemic Equilibrium
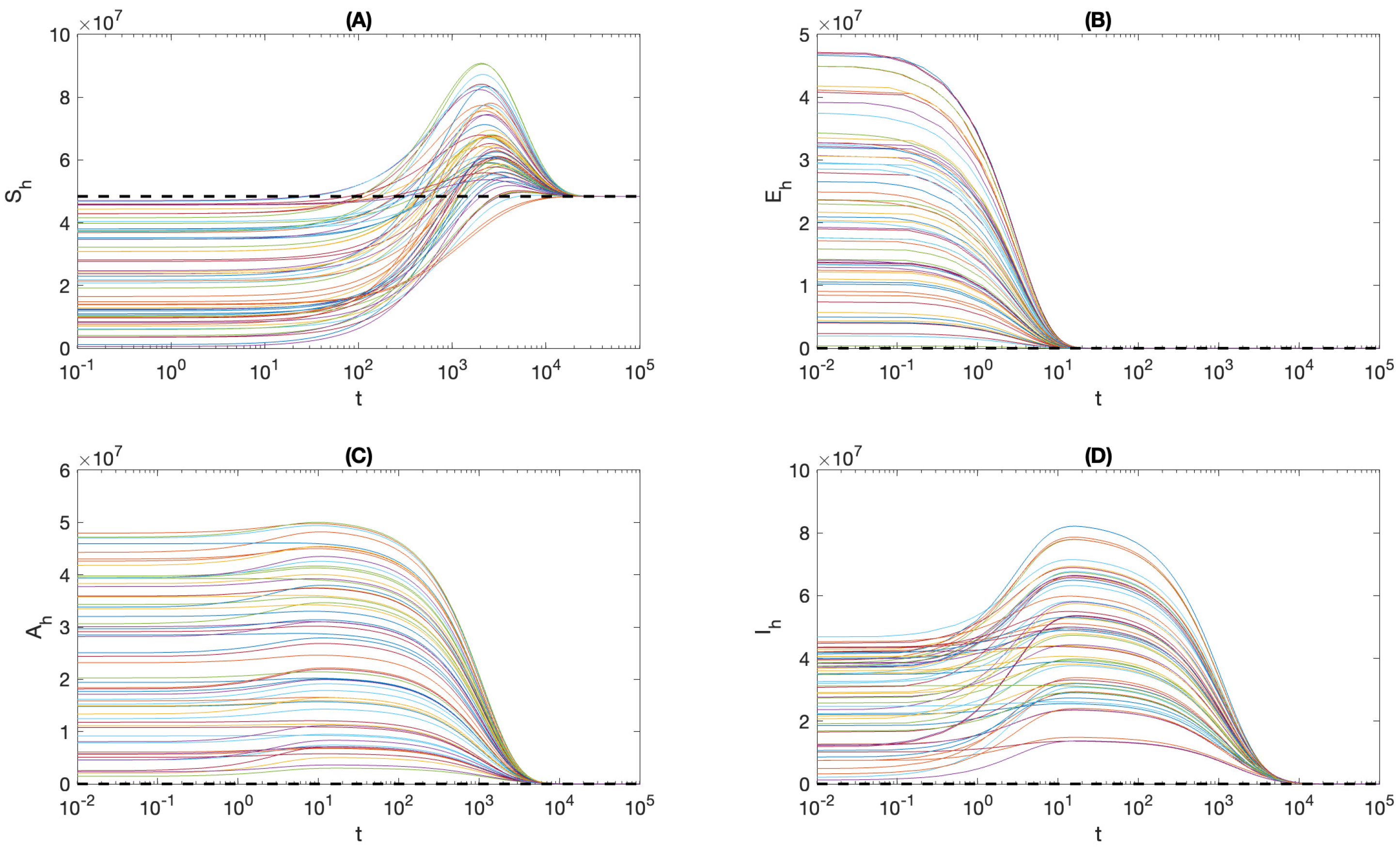
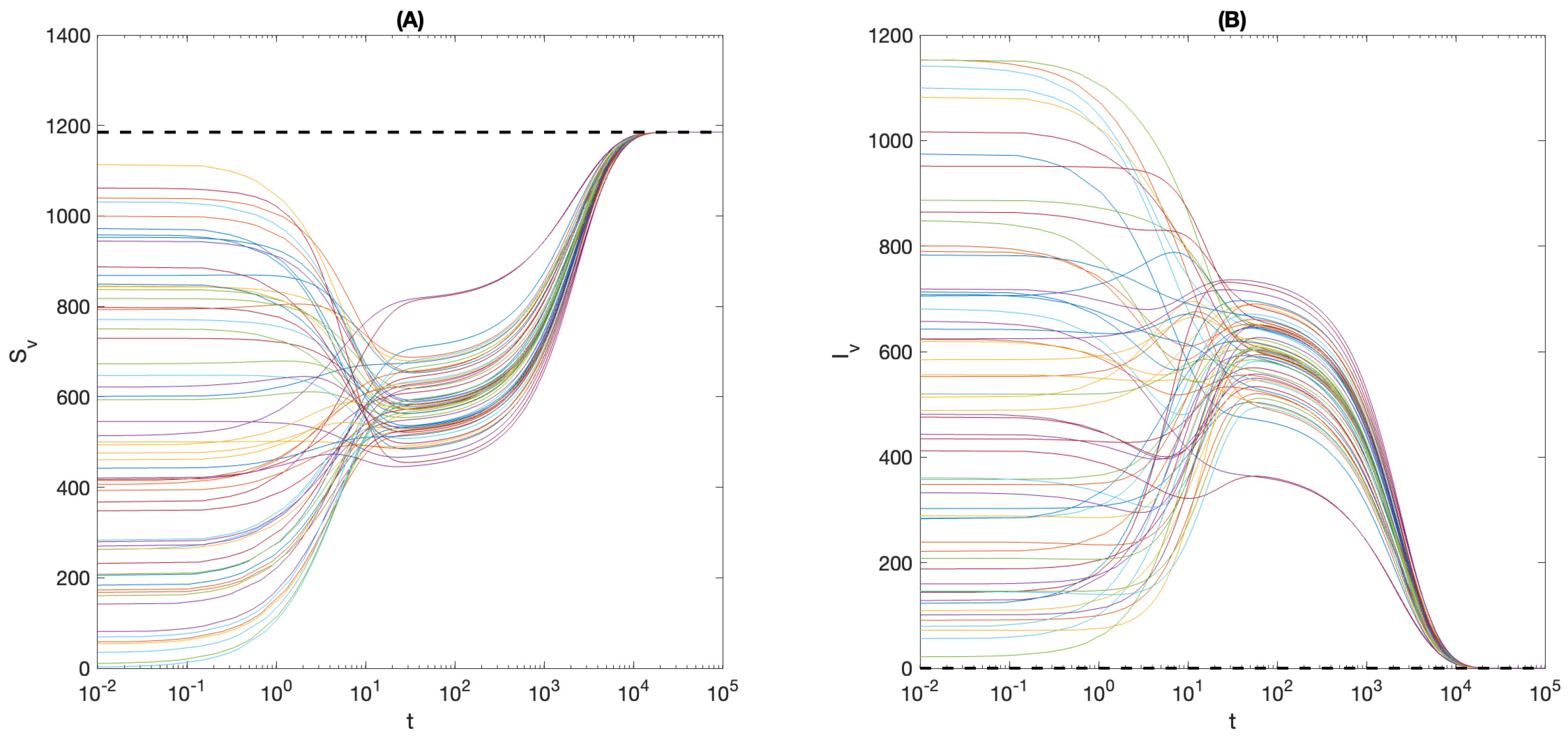
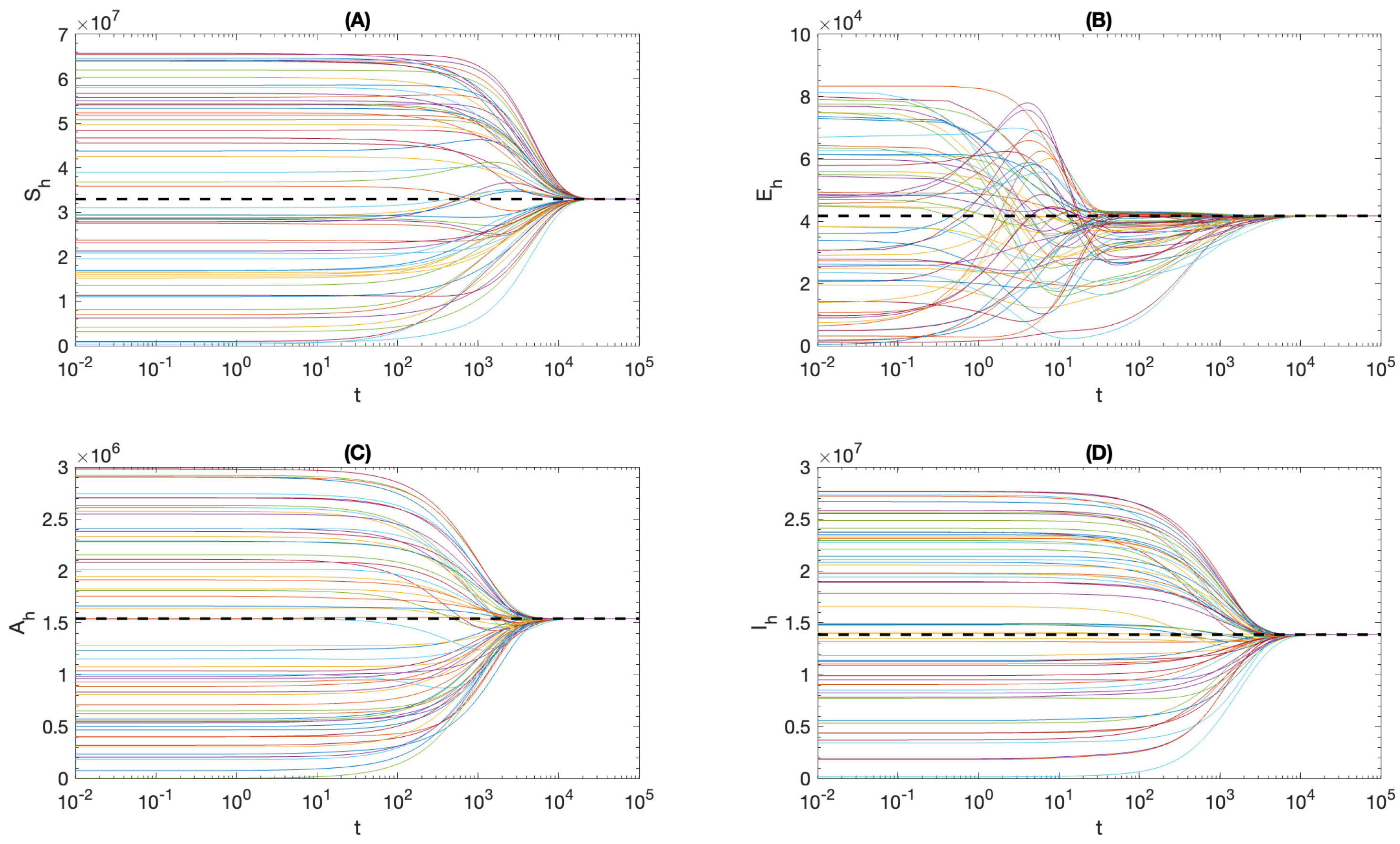
4. Local Stability Analysis
4.1. Local Stability of
4.2. Local Stability of the Endemic Equilibrium
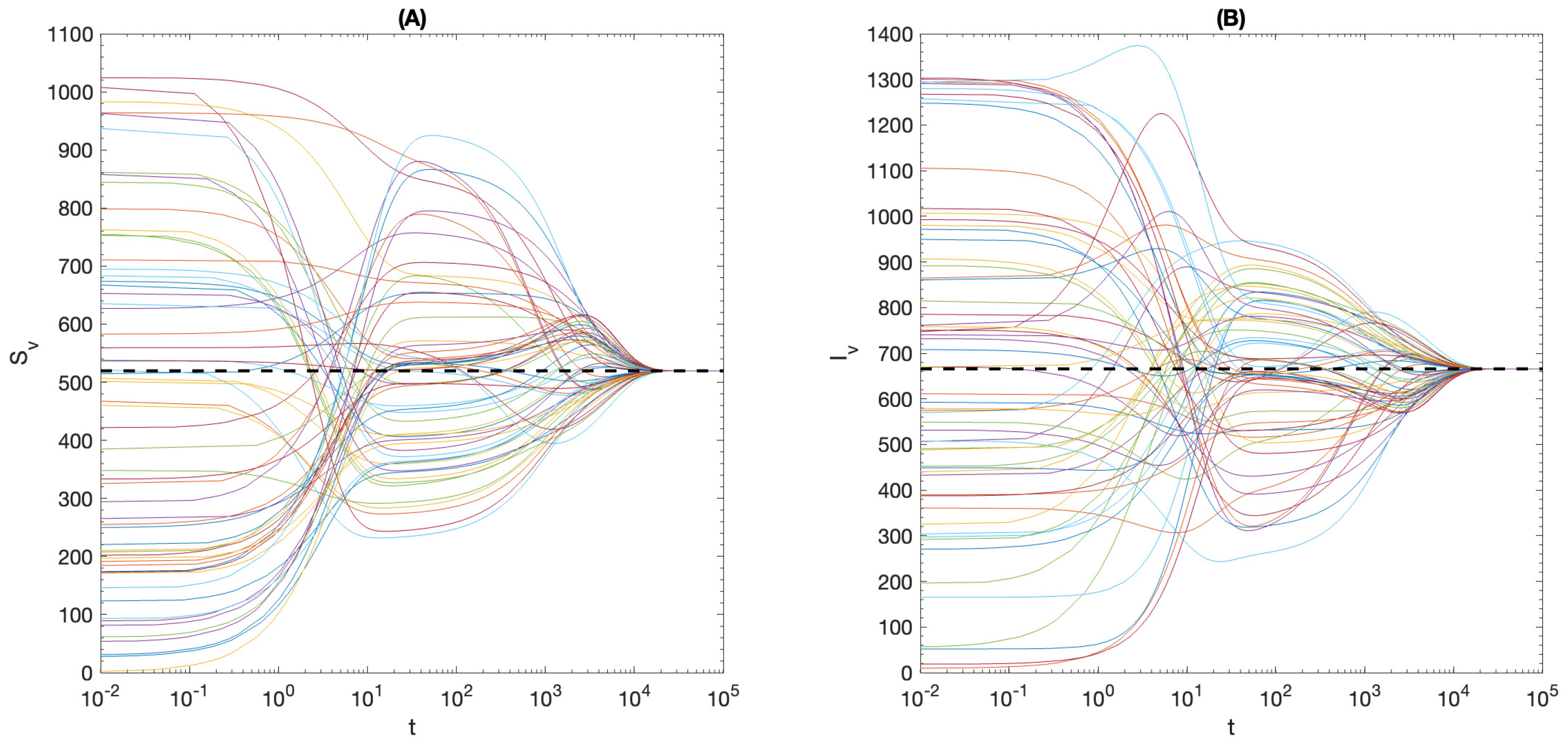
5. Global and Asymptotic Stability
- (H1)
- For is globally asymptotically stable.
- (H2)
- for .
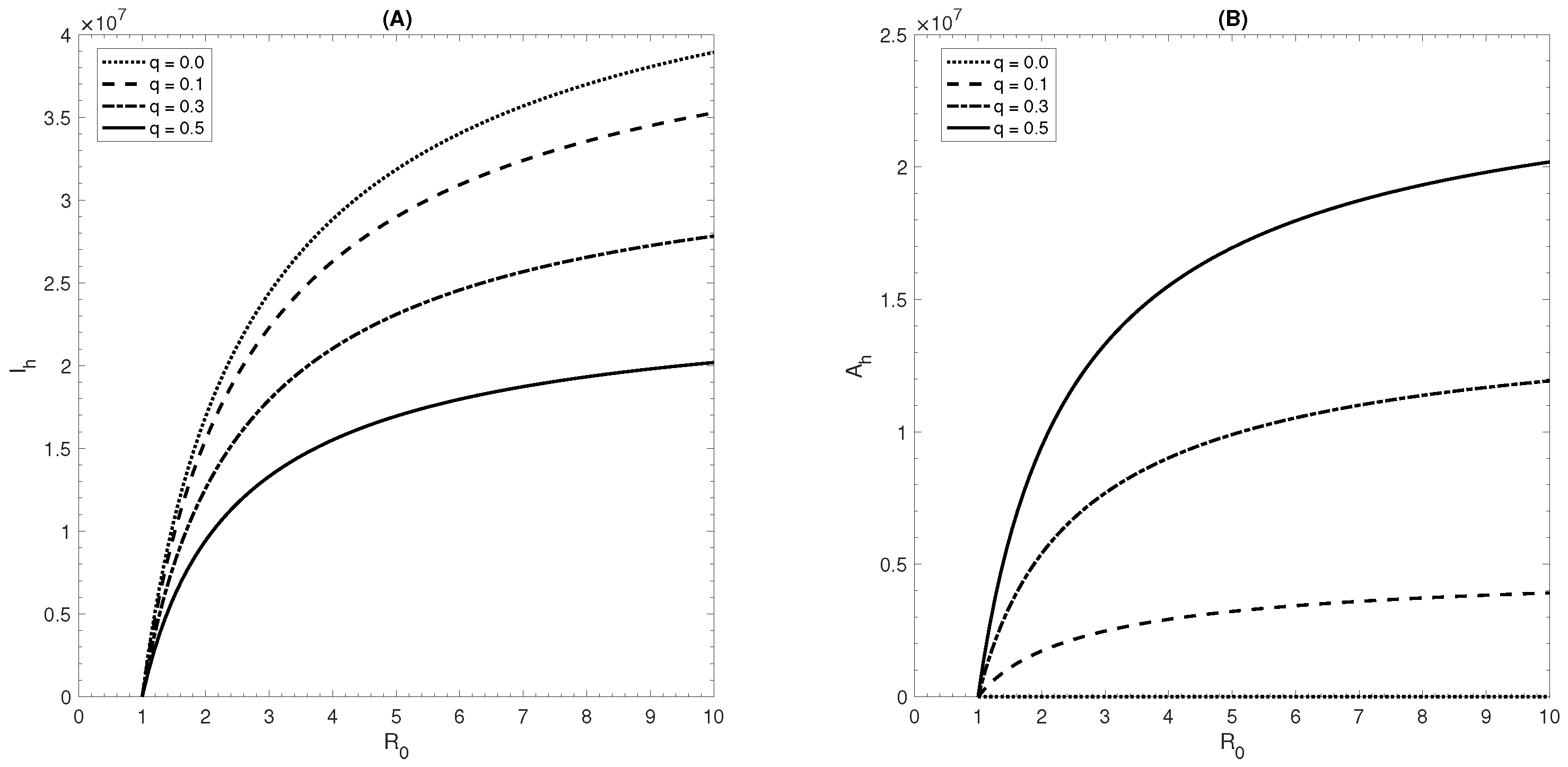
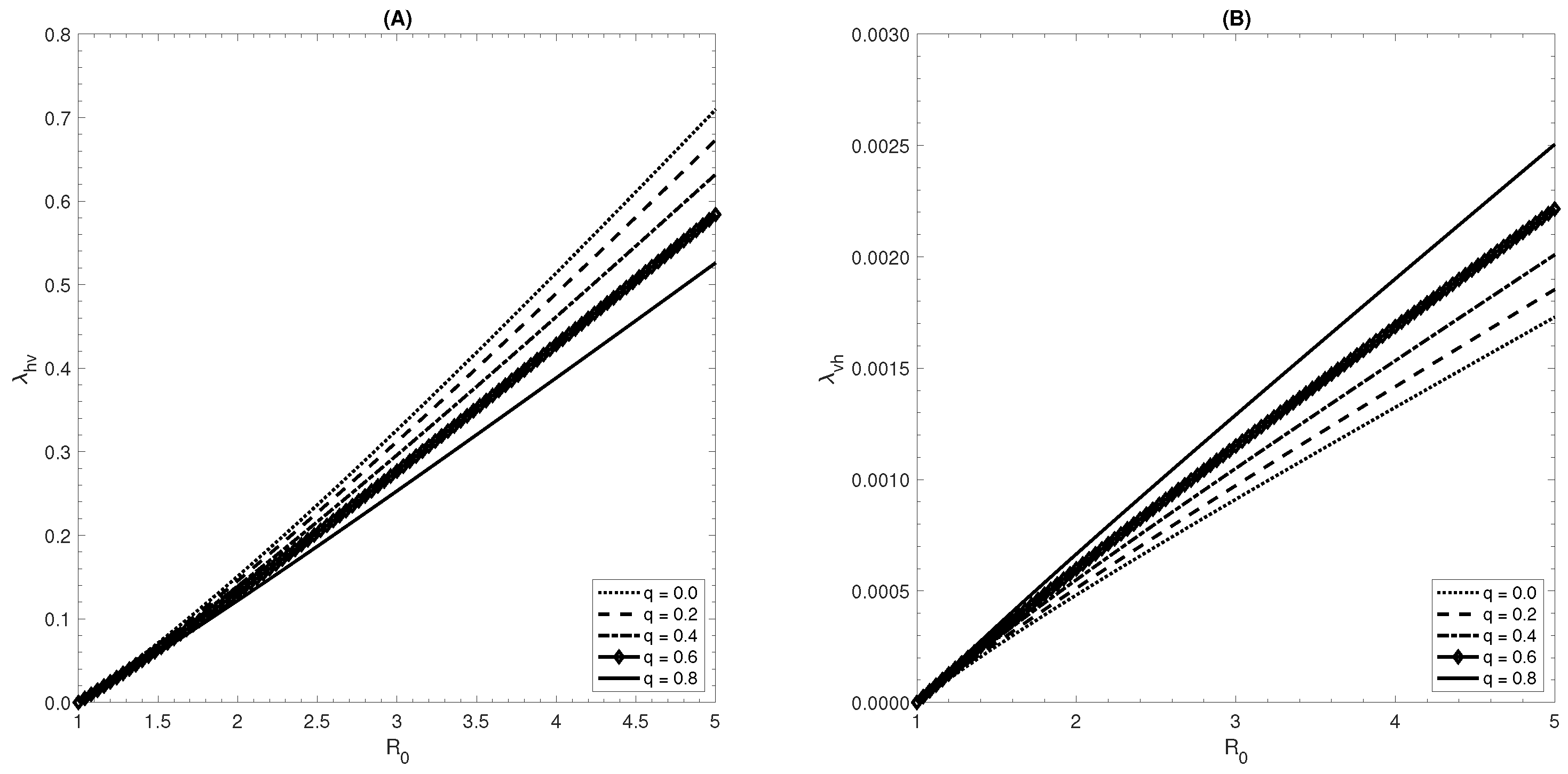
6. Effect of Ignoring Asymptomatic Infections
7. Summary and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Locally Lipschitz
Appendix B. Positivity and Boundedness—Proof of Proposition 1
Appendix C. Proof of the Positivity of the Hurwitz Determinant
References
- de Vries, H.J.; Reedijk, S.H.; Schallig, H.D. Cutaneous leishmaniasis: Recent developments in diagnosis and management. Am. J. Clin. Dermatol. 2015, 16, 99–109. [Google Scholar] [CrossRef] [PubMed]
- Karimi, T.; Sharifi, I.; Aflatoonian, M.R.; Aflatoonian, B.; Mohammadi, M.A.; Salarkia, E.; Babaei, Z.; Zarinkar, F.; Sharifi, F.; Hatami, N.; et al. A long-lasting emerging epidemic of anthroponotic cutaneous leishmaniasis in southeastern Iran: Population movement and peri-urban settlements as a major risk factor. Parasites Vectors 2021, 14, 122. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization, Leishmaniasis. Available online: https://www.who.int/news-room/fact-sheets/detail/leishmaniasis (accessed on 2 January 2023).
- Abuzaid, A.A.; Abdoon, A.M.; Aldahan, M.A.; Alzahrani, A.G.; Alhakeem, R.F.; Asiri, A.M.; Alzahrani, M.H.; Memish, Z.A. Cutaneous leishmaniasis in Saudi Arabia: A comprehensive overview. Vector-Borne Zoonotic Dis. 2017, 17, 673–684. [Google Scholar] [CrossRef] [PubMed]
- Haouas, N.; Amer, O.; Alshammri, F.F.; Al-Shammari, S.; Remadi, L.; Ashankyty, I. Cutaneous leishmaniasis in northwestern Saudi Arabia: Identification of sand fly fauna and parasites. Parasites Vectors 2017, 10, 544. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Kuang, Y.; Wu, C.H.; Ben-Arieh, D.; Ramalho-Ortigao, M.; Bi, K. Zoonotic visceral leishmaniasis transmission: Modeling, backward bifurcation, and optimal control. J. Math. Biol. 2016, 73, 1525–1560. [Google Scholar] [CrossRef]
- Bi, K.; Chen, Y.; Zhao, S.; Ben-Arieh, D.; Wu, C.H.J. A new zoonotic visceral leishmaniasis dynamic transmission model with age-structure. Chaos Solitons Fractals 2020, 133, 109622. [Google Scholar] [CrossRef]
- Barley, K.; Mubayi, A.; Safan, M.; Castillo-Chavez, C. A comparative assessment of visceral leishmaniasis burden in two eco-epidemiologically different countries, India and Sudan. bioRxiv 2019. [Google Scholar] [CrossRef]
- Hussaini, N.; Okuneye, K.; Gumel, A.B. Mathematical analysis of a model for zoonotic visceral leishmaniasis. Infect. Dis. Model. 2017, 2, 455–474. [Google Scholar] [CrossRef]
- Kaabi, B.; Zhioua, E. Modeling and comparative study of the spread of zoonotic visceral leishmaniasis from Northern to Central Tunisia. Acta Trop. 2018, 178, 19–26. [Google Scholar] [CrossRef]
- Rock, K.S.; Quinnell, R.J.; Medley, G.F.; Courtenay, O. Progress in the mathematical modelling of visceral leishmaniasis. Adv. Parasitol. 2016, 94, 49–131. [Google Scholar]
- Chaves, L.F.; Hernandez, M.J. Mathematical modelling of American cutaneous leishmaniasis: Incidental hosts and threshold conditions for infection persistence. Acta Trop. 2004, 92, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Bacaér, N.; Guernaoui, S. The epidemic threshold of vector-borne diseases with seasonality: The case of cutaneous leishmaniasis in Chichaoua, Morocco. J. Math. Biol. 2006, 53, 421–436. [Google Scholar] [CrossRef] [PubMed]
- Barradas, I.; Caja Rivera, R.M. Cutaneous leishmaniasis in Peru using a vector-host model: Backward bifurcation and sensitivity analysis. Math. Methods Appl. Sci. 2018, 41, 1908–1924. [Google Scholar] [CrossRef]
- Zamir, M.; Zaman, G.; Alshomrani, A.S. Sensitivity analysis and optimal control of anthroponotic cutaneous leishmania. PLoS ONE 2016, 11, e0160513. [Google Scholar] [CrossRef]
- De Almeida, M.C.; Moreira, H.N. A mathematical model of immune response in cutaneous leishmaniasis. J. Biol. Syst. 2007, 15, 313–354. [Google Scholar] [CrossRef]
- Agyingi, E.O.; Ross, D.S.; Bathena, K. A model of the transmission dynamics of leishmaniasis. J. Biol. Syst. 2011, 19, 237–250. [Google Scholar] [CrossRef]
- Biswas, D.; Kesh, D.K.; Datta, A.; Chatterjee, A.N.; Roy, P.K. A mathematical approach to control cutaneous leishmaniasis through insecticide spraying. SOP Trans. Appl. Math. 2014, 1, 44–54. [Google Scholar] [CrossRef]
- Saudi Ministry of Health, Communicable Diseases (Leishmaniasis). Available online: https://www.moh.gov.sa/en/HealthAwareness/EducationalContent/Diseases/Infectious/Pages/016.aspx (accessed on 15 May 2023).
- World Population Review. 2021. Available online: https://worldpopulationreview.com/continents/sub--saharan-africa-population (accessed on 21 July 2022).
- Rabinovich, J.E.; Feliciangeli, M.D. Parameters of Leishmania braziliensis transmission by indoor Lutzomyia ovallesi in Venezuela. Am. J. Trop. Med. Hyg. 2004, 70, 373–382. [Google Scholar] [CrossRef]
- Sierra, D.; Vélez, I.D.; Uribe, S. Identificacion de Lutzomyia Spp. (Diptera: Psychodidae) Grupo verrucarum Por Medio De Microsc. Electron. De Sus Huevos. Rev. Biol. Trop. 2000, 48, 615–622. [Google Scholar] [CrossRef]
- Piscopo, T.V.; Mallia Azzopardi, C. Leishmaniasis. Postgrad Med. J. 2007, 83, 649–657. [Google Scholar] [CrossRef]
- Valle, P.A.; Coria, L.N.; Plata, C.; Salazar, Y. CAR-T cell therapy for the treatment of ALL: Eradication conditions and in silico experimentation. Hemato 2021, 2, 441–462. [Google Scholar] [CrossRef]
- De Leenheer, P.; Aeyels, D. Stability properties of equilibria of classes of cooperative systems. IEEE Trans. Autom. Control 2001, 46, 1996–2001. [Google Scholar] [CrossRef]
- Van den Driessche, P.; Watmough, J. Reproduction numbers and sub-threshold endemic equilibria for compartmental models of disease transmission. Math. Biosci. 2002, 180, 29–48. [Google Scholar] [CrossRef] [PubMed]
- Chavez, C.C.; Feng, Z.; Huang, W. On the computation of R0 and its role on global stability. In Mathematical Approaches for Emerging and Re-Emerging Infection Diseases: An Introduction; The IMA Volumes in Mathematics and Its Applications; Springer: Berlin/Heidelberg, Germany, 2002; Volume 125, pp. 31–65. [Google Scholar]
- Safan, M. Mathematical analysis of an SIR respiratory infection model with sex and gender disparity: Special reference to influenza A. Math. Biosci. Eng. 2019, 16, 2613–2649. [Google Scholar] [CrossRef] [PubMed]
- Colquitt, R.B.; Colquhoun, D.A.; Thiele, R.H. In silico modelling of physiologic systems. Best Pract. Res. Clin. Anaesthesiol. 2011, 25, 499–510. [Google Scholar] [CrossRef]
- Bathena, K. A Mathematical Model of Cutaneous Leishmaniasis. Master’s Thesis, School of Mathematica Sciences, Rochester Institute of Technology, Rochester, NY, USA, 2009. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| State Variables and Parameters | Description |
|---|---|
| Number of susceptible humans at time t. | |
| Number of exposed humans in the latent period at time t. | |
| Number of asymptomatic humans (infected but without noticeable disease and infectious ) at time t. | |
| Number of infected humans at time t. | |
| Number of susceptible sandflies at time t. | |
| Number of infectious sandflies at time t. | |
| Total human population size at time t. | |
| Total sandflies population size at time t. | |
| Recruitment rate of humans. | |
| Recruitment rate of sandflies. | |
| Natural death rate of humans. | |
| Natural death rate of sandflies. | |
| Recovery rate (with temporal immunity) from symptomatic/asymptomatic cutaneous Leishmaniasis (CL) infection. | |
| The rate at which human individuals leave the exposed state. | |
| The rate at which sandflies bite the body of human individuals. | |
| q | The probability that an exposed human develops asymptomatic CL infection after leaving the incubation period. |
| r | The relative transmissibility of asymptomatic with respect to symptomatic CL infection. |
| The probability at which a susceptible sandfly acquires CL infection while taking its meal from a CL-infected human. | |
| The probability at which a susceptible human acquires CL infection while being bitten by a CL-infected sandfly. |
| Parameter | Value | Range | Unit | Reference |
|---|---|---|---|---|
| 12,327 | – | Per week | [20] | |
| 112 | [35–350] | Per week | [14,21] | |
| – | Per week | [20] | ||
| 0.0945 | [0.077, 0.525] | Per week | [14,22] | |
| 0.0006391 | [0.0000791–0.004991] | Per week | [14,21] | |
| 0.33 | [0.125, 0.5] | Per week | [23] | |
| 0.48146 | [4.1944–18.7754] | Per week | [14,21] | |
| q | 0.1 | – | Dimensionless | Assumed |
| r | 0.3 | – | Dimensionless | Assumed |
| 0.0097 | [0.0028–0.08] | Dimensionless | [14,21] | |
| 0.7198 | [0.08–0.9] | Dimensionless | [14] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Safan, M.; Altheyabi, A. Mathematical Analysis of an Anthroponotic Cutaneous Leishmaniasis Model with Asymptomatic Infection. Mathematics 2023, 11, 2388. https://doi.org/10.3390/math11102388
Safan M, Altheyabi A. Mathematical Analysis of an Anthroponotic Cutaneous Leishmaniasis Model with Asymptomatic Infection. Mathematics. 2023; 11(10):2388. https://doi.org/10.3390/math11102388
Chicago/Turabian StyleSafan, Muntaser, and Alhanouf Altheyabi. 2023. "Mathematical Analysis of an Anthroponotic Cutaneous Leishmaniasis Model with Asymptomatic Infection" Mathematics 11, no. 10: 2388. https://doi.org/10.3390/math11102388