
Does Local Adaptation Impact on the Distribution of Competing Aedes Disease Vectors?


{kind=link}
{kind=link}
Abstract
:1. Introduction
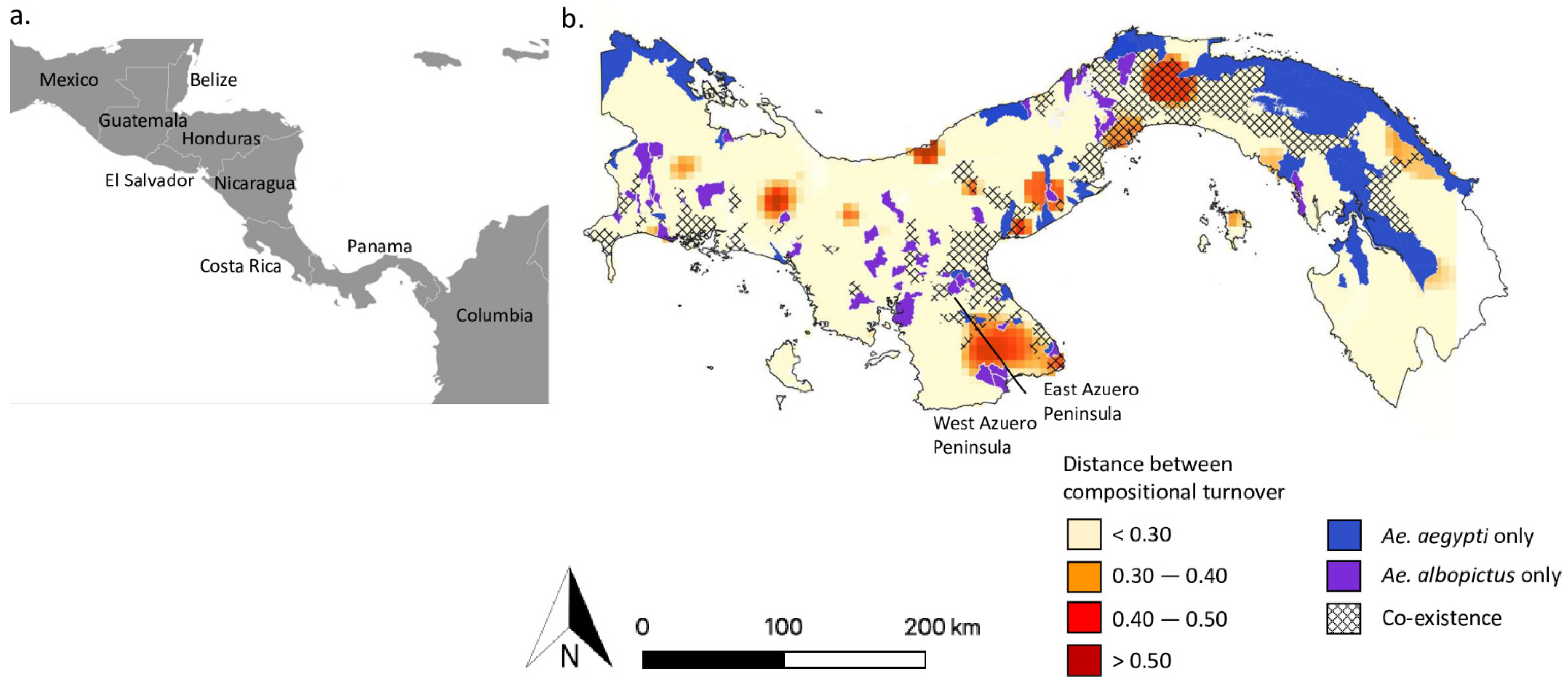
2. The Displacement of Ae. aegypti by Ae. albopictus Is Context-Dependent
3. The Genomic Signal of Local Environmental Adaptation in Ae. aegypti
4. Local Adaptation in Ae. aegypti Determines Coexistence with Ae. albopictus
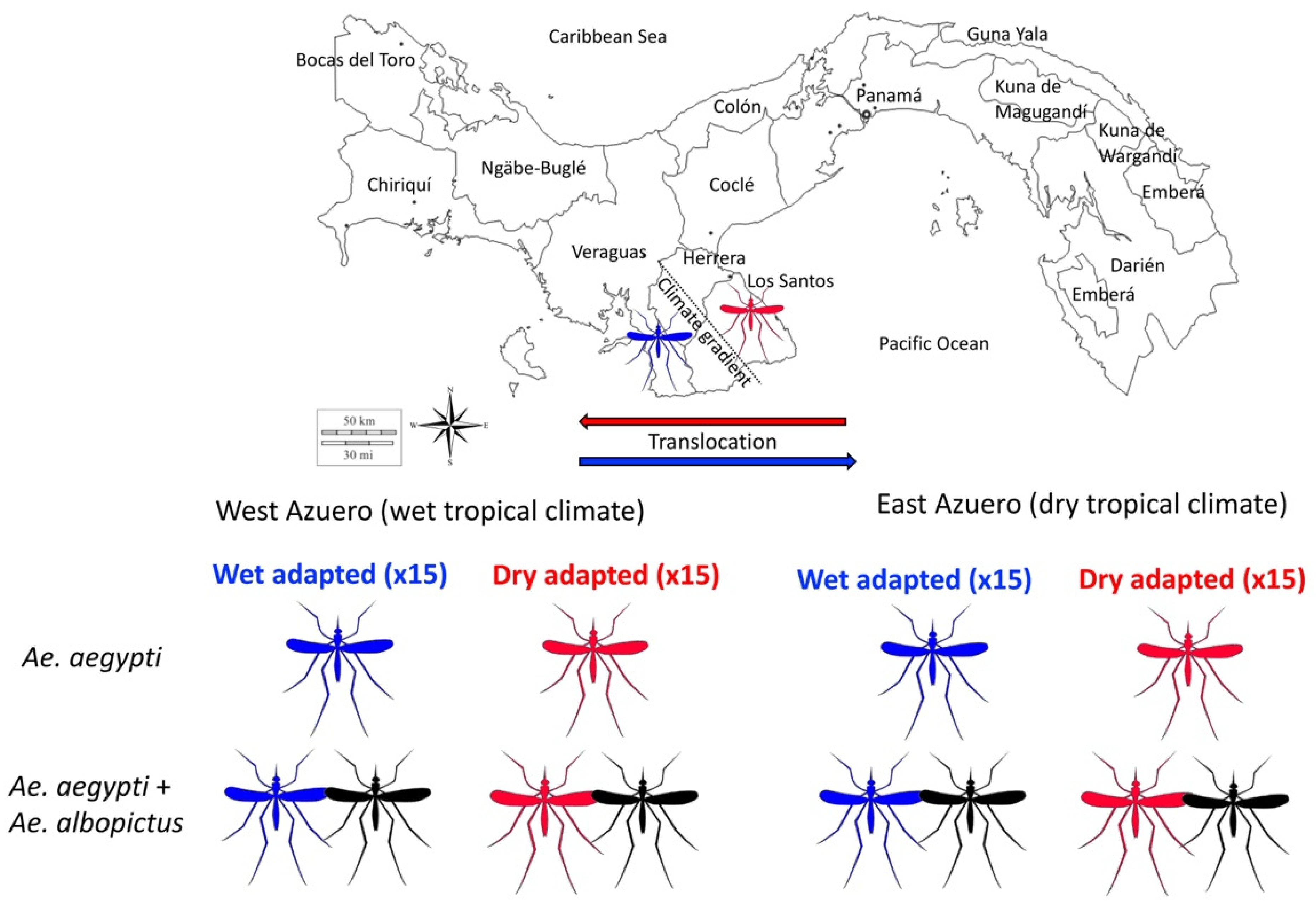
5. An Experimental Framework to Test for Local Adaptation in the Presence of a Competitor
6. Implications
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Yellow Fever Fact Sheet No. 100. 2014. Available online: www.who.int/mediacentre/factsheets/fs100/en (accessed on 7 July 2020).
- World Health Organization. World Health Report. Insect-Borne Diseases. Available online: www.who.int/whr/1996/media_centre/executive_summary1/en/index9.html (accessed on 7 July 2020).
- Barrett, A.D.T. The reemergence of yellow fever. Science 2018, 361, 847–848. [Google Scholar] [CrossRef]
- Roiz, D.; Wilson, A.L.; Scott, T.W.; Fonseca, D.M.; Jourdain, F.; Müller, P.; Velayudhan, R.; Corbel, V. Integrated Aedes management for the control of Aedes-borne diseases. PLoS Negl. Trop. Dis. 2018, 12, e0006845. [Google Scholar] [CrossRef] [Green Version]
- López-Camacho, C.; Kim, Y.C.; Abbink, P.; Larocca, R.A.; Huiskonen, J.T.; Barouch, D.H.; Reyes-Sandoval, A. Assessment of immunogenicity and efficacy of a Zika vaccine using modified vaccinia ankara virus as carriers. Pathogens 2019, 8, 216. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.K.; Dhama, K.; Khandia, R.; Munjal, A.; Karthik, K.; Tiwari, R.; Chakraborty, S.; Malik, Y.S.; Bueno-Marí, R. Prevention and control strategies to counter Zika virus, a special focus on intervention approaches against vector mosquitoes-current updates. Front. Microbiol. 2018, 9, 87. [Google Scholar] [CrossRef] [Green Version]
- Xu, C.; Legros, M.; Gould, F.; Lloyd, A.L. Understanding uncertainties in model-based predictions of Ae. aegypti population dynamics. PLoS Negl. Trop. Dis. 2010, 4, e830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kearney, M.; Porter, W.P.; Williams, C.; Ritchie, S.; Hoffmann, A.A. Integrating biophysical models and evolutionary theory to predict climatic impacts on species’ ranges: The dengue mosquito Ae. aegypti in Australia. Funct. Ecol. 2009, 23, 528–538. [Google Scholar] [CrossRef]
- Lounibos, L.P.; Juliano, S.A. Where vectors collide: The importance of mechanisms shaping the realized niche for modeling ranges of invasive Ae. mosquitoes. Biol. Invasions 2018, 20, 1913–1929. [Google Scholar] [CrossRef] [PubMed]
- Hancock, P.A.; White, V.L.; Ritchie, S.A.; Hoffmann, A.A.; Godfray, H.C.J. Predicting Wolbachia invasion dynamics in Ae. aegypti populations using models of density-dependent demographic traits. BMC Biol. 2016, 14, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kraemer, M.U.G.; Sinka, M.E.; Duda, K.A.; Mylne, A.Q.N.; Shearer, F.M.; Barker, C.M.; Moore, C.G.; Carvalho, R.G.; Coelho, G.E.; Van Bortel, W.; et al. The global distribution of the arbovirus vectors Ae. aegypti and Ae. albopictus. eLife 2015, 4, e08347. [Google Scholar] [CrossRef]
- Bargielowski, I.; Carrasquilla, M.C.; Nishimura, N.; Lounibos, L.P. Coexistence of Ae. aegypti and Ae. albopictus (Diptera: Culicidae) in Peninsular Florida two decades after competitive displacements. J. Med. Entomol. 2016, 53, 1385–1390. [Google Scholar] [CrossRef]
- O’Meara, G.F.; Evans Leonard, F.J.; Gettman, A.D.; Cuda, J.P. Spread of Ae. albopictus and decline of Ae. aegypti (Diptera: Culicidae) in Florida. J. Med. Entomol. 1995, 32, 554–562. [Google Scholar] [CrossRef]
- Braks, M.A.H.; Honório, N.A.; Lounibos, L.P.; Lourenço-de-Oliveira, R.; Juliano, S.A. Interspecific competition between two invasive species of container mosquitoes, Ae. aegypti and Ae. albopictus (Diptera: Culicidae), in Brazil. Ann. Entomol. Soc. Am. 2004, 97, 130–139. [Google Scholar] [CrossRef]
- Juliano, S.A. Species Introduction and replacement among mosquitoes: Interspecific resource competition or apparent competition? Ecology 1998, 79, 255–268. [Google Scholar] [CrossRef]
- Kaplan, L.; Kendell, D.; Robertson, D.; Livdahl, T.; Khatchikian, C. Ae. aegypti and Ae. albopictus in Bermuda: Extinction, invasion, invasion and extinction. Biol. Invasions 2010, 12, 3277–3288. [Google Scholar] [CrossRef]
- Leisnham, P.; Juliano, S. Interpopulation differences in competitive effect and response of the mosquito Ae. aegypti and resistance to invasion by a superior competitor. Oecologia 2010, 164, 221–230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raharimalala, F.N.; Ravaomanarivo, L.H.; Ravelonandro, P.; Rafarasoa, L.S.; Zouache, K.; Tran-Van, V.; Mousson, L.; Failloux, A.-B.; Hellard, E.; Moro, C.V.; et al. Biogeography of the two major arbovirus mosquito vectors, Ae. aegypti and Ae. albopictus (Diptera, Culicidae), in Madagascar. Parasit. Vectors 2012, 5, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagny Beilhe, L.; Delatte, H.; Quilici, S.; Fontenille, D. Progressive decrease in Ae. aegypti distribution in Reunion Island since the 1900s. J. Med. Entomol. 2009, 46, 1541–1545. [Google Scholar] [CrossRef]
- Bagny Beilhe, L.; Arnoux, S.; Delatte, H.; Lajoie, G.; Fontenille, D. Spread of invasive Ae. albopictus and decline of resident Ae. aegypti in urban areas of Mayotte 2007–2010. Biol. Invasions 2012, 14, 1623–1633. [Google Scholar] [CrossRef]
- Bennett, K.; McMillan, W.; Enríquez, V.; Barraza, E.; Díaz, M.; Baca, B.; Whiteman, A.; Medina, J.; Ducasa, M.; Gómez Martínez, C.; et al. The role of heterogeneous environmental conditions in shaping the spatiotemporal distribution of competing Ae. mosquitoes in Panama: Implications for the landscape of arboviral disease. Biol. Invasions 2021, in press. [Google Scholar]
- Sherpa, S.; Guéguen, M.; Renaud, J.; Blum, M.G.B.; Gaude, T.; Laporte, F.; Akiner, M.; Alten, B.; Aranda, C.; Barre-Cardi, H.; et al. Predicting the success of an invader: Niche shift versus niche conservatism. Ecol. Evol. 2019, 9, 12658–12675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tun-Lin, W.; Burkot, T.R.; Kay, B.H. Effects of temperature and larval diet on development rates and survival of the dengue vector Ae. aegypti in north Queensland, Australia. Med. Vet. Entomol. 2000, 14, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Tripet, F.; Lounibos, L.P.; Robbins, D.; Moran, J.; Nishimura, N.; Blosser, E.M. Competitive reduction by satyrization? Evidence for interspecific mating in nature and asymmetric reproductive competition between invasive mosquito vectors. Am. J. Trop. Med. Hyg. 2011, 85, 265–270. [Google Scholar] [CrossRef]
- Lounibos, L.P.; Kramer, L.D. Invasiveness of Ae. aegypti and Ae. albopictus and vectorial capacity for chikungunya virus. J. Infect. Dis. 2016, 214, S453–S458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hopperstad, K.A.; Reiskind, M.H. Recent changes in the local distribution of Ae. aegypti (Diptera: Culicidae) in South Florida, USA. J. Med. Entomol. 2016, 53, 836–842. [Google Scholar] [CrossRef]
- Reiskind, M.H.; Lounibos, L.P. Spatial and temporal patterns of abundance of Ae. aegypti L. (Stegomyia aegypti) and Ae. albopictus (Stegomyia albopictus) (Skuse) in southern Florida. Med. Vet. Entomol. 2013, 27, 421–429. [Google Scholar] [CrossRef]
- Juliano, S.A.; O’Meara, G.F.; Morrill, J.R.; Cutwa, M.M. Desiccation and thermal tolerance of eggs and the coexistence of competing mosquitoes. Oecologia 2002, 130, 458–469. [Google Scholar] [CrossRef] [Green Version]
- Carroll, S.P. Facing change: Forms and foundations of contemporary adaptation to biotic invasions. Mol. Ecol. 2008, 17, 361–372. [Google Scholar] [CrossRef]
- Leibold, M.A.; Urban, M.C.; De Meester, L.; Klausmeier, C.A.; Vanoverbeke, J. Regional neutrality evolves through local adaptive niche evolution. Proc. Natl. Acad. Sci. USA 2019, 116, 2612–2617. [Google Scholar] [CrossRef] [Green Version]
- Pinsky, M.L. Species coexistence through competition and rapid evolution. Proc. Natl. Acad. Sci. USA 2019, 116, 2407–2409. [Google Scholar] [CrossRef] [Green Version]
- Urban, M.C.; De Meester, L. Community monopolization: Local adaptation enhances priority effects in an evolving metacommunity. Proc. R. Soc. Biol. Sci. 2009, 276, 4129–4138. [Google Scholar] [CrossRef] [PubMed]
- Case, T.J.; Taper, M.L. Interspecific competition, environmental gradients, gene flow, and the coevolution of species’ borders. Am. Nat. 2000, 155, 583–605. [Google Scholar] [CrossRef]
- Sherpa, S.; Blum, M.G.B.; Després, L. Cold adaptation in the Asian tiger mosquito’s native range precedes its invasion success in temperate regions. Evolution 2019, 73, 1793–1808. [Google Scholar] [CrossRef] [PubMed]
- Sherpa, S.; Rioux, D.; Goindin, D.; Fouque, F.; François, O.; Després, L. At the origin of a worldwide invasion: Unraveling the genetic makeup of the caribbean bridgehead populations of the dengue vector Ae. aegypti. Genome Biol. Evol. 2018, 10, 56–71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, K.L.; McMillan, W.O.; Loaiza, J.R. The genomic signal of local adaptation in Ae. aegypti mosquitoes. Evol. Appl. 2021, in press. [Google Scholar] [CrossRef]
- Brady, O.J.; Johansson, M.A.; Guerra, C.A.; Bhatt, S.; Golding, N.; Pigott, D.M.; Delatte, H.; Grech, M.G.; Leisnham, P.T.; Maciel-de-Freitas, R.; et al. Modelling adult Ae. aegypti and Ae. albopictus survival at different temperatures in laboratory and field settings. Parasit. Vectors 2013, 6, 351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lounibos, L.P.; O’Meara, G.F.; Juliano, S.A.; Nishimura, N.; Escher, R.L.; Reiskind, M.H.; Cutwa, M.; Greene, K. Differential survivorship of invasive mosquito species in South Florida cemeteries: Do site-specific microclimates explain patterns of coexistence and exclusion? Ann. Entomol. Soc. Am. 2010, 103, 757–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vezzani, D.; Rubio, A.; Velázquez, S.M.; Schweigmann, N.; Wiegand, T. Detailed assessment of microhabitat suitability for Ae. aegypti (Diptera: Culicidae) in Buenos Aires, Argentina. Acta Trop. 2005, 95, 123–131. [Google Scholar] [CrossRef]
- Eskildsen, G.A.; Rovira, J.R.; Dutari, L.C.; Smith, O.; Miller, M.J.; Bennett, K.L.; McMillan, W.O.; Loaiza, J.R. Maternal invasion history of Ae. aegypti and Ae. albopictus into the Isthmus of Panama: Implications for the control of emergent viral disease agents. PLoS ONE 2018, 13, e0194874. [Google Scholar] [CrossRef] [Green Version]
- Bennett, K.L.; Martínez Gómez, C.; Almanza, A.; Rovira, J.R.; McMillan, W.O.; Enriquez, V.; Barraza, E.; Diaz, M.; Galan, J.S.; Whiteman, A.; et al. High infestation of invasive Ae. mosquitoes along the highways of Panama: Impact on vector ecology and disease control. Parasit. Vectors 2019, 12, 264. [Google Scholar] [CrossRef] [Green Version]
- Juneja, P.; Ariani, C.V.; Ho, Y.S.; Akorli, J.; Palmer, W.J.; Pain, A.; Jiggins, F.M. Exome and transcriptome sequencing of Ae. aegypti identifies a locus that confers resistance to Brugia malayi and alters the immune response. PLoS Pathog. 2015, 11, e1004765. [Google Scholar] [CrossRef] [Green Version]
- Faucon, F.; Dusfour, I.; Gaude, T.; Navratil, V.; Boyer, F.; Chandre, F.; Sirisopa, P.; Thanispong, K.; Juntarajumnong, W.; Poupardin, R.; et al. Identifying genomic changes associated with insecticide resistance in the dengue mosquito Ae. aegypti by deep targeted sequencing. Genome Res. 2015, 25, 1347–1359. [Google Scholar] [CrossRef] [Green Version]
- Bradshaw, W.E.; Emerson, K.J.; Catchen, J.M.; Cresko, W.A.; Holzapfel, C.M. Footprints in time: Comparative quantitative trait loci mapping of the pitcher-plant mosquito, Wyeomyia smithii. Proc. R. Soc. Biol. Sci. 2012, 279, 4551–4558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, K.E.; Flick, D.; Fleming, K.H.; Jochim, R.; Beaty, B.J.; Black, W.C., IV. Quantitative Trait Loci that control dengue-2 virus dissemination in the mosquito Ae. aegypti. Genetics 2005, 170, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Josephs, E.B.; Berg, J.J.; Ross-Ibarra, J.; Coop, G. Detecting Adaptive Differentiation in Structured Populations with Genomic Data and Common Gardens. Genetics 2019, 211, 989–1004. [Google Scholar] [CrossRef] [Green Version]
- Montesinos-López, O.A.; Martín-Vallejo, J.; Crossa, J.; Gianola, D.; Hernández-Suárez, C.M.; Montesinos-López, A.; Juliana, P.; Singh, R. New Deep Learning Genomic-Based Prediction Model for Multiple Traits with Binary, Ordinal, and Continuous Phenotypes. G3 Genes Genomes Genet. 2019, 9, 1545–1556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldvogel, A.-M.; Feldmeyer, B.; Rolshausen, G.; Exposito-Alonso, M.; Rellstab, C.; Kofler, R.; Mock, T.; Schmid, K.; Schmitt, I.; Bataillon, T.; et al. Evolutionary genomics can improve prediction of species’ responses to climate change. Evol. Lett. 2020, 4, 4–18. [Google Scholar] [CrossRef]
- Li, M.; Yang, T.; Kandul, N.P.; Bui, M.; Gamez, S.; Raban, R.; Bennett, J.; Sánchez, C.H.M.; Lanzaro, G.C.; Schmidt, H.; et al. Development of a confinable gene drive system in the human disease vector Ae. aegypti. eLife 2020, 9, e51701. [Google Scholar] [CrossRef] [PubMed]
- Faraji, A.; Egizi, A.; Fonseca, D.M.; Unlu, I.; Crepeau, T.; Healy, S.P.; Gaugler, R. Comparative host feeding patterns of the Asian tiger mosquito, Ae. albopictus, in urban and suburban Northeastern USA and implications for disease transmission. PLoS Negl. Trop. Dis. 2014, 8, e3037. [Google Scholar] [CrossRef] [Green Version]
- Pereira Dos Santos, T.; Roiz, D.; Santos de Abreu, F.V.; Luz, S.L.B.; Santalucia, M.; Jiolle, D.; Santos Neves, M.S.A.; Simard, F.; Lourenço-de-Oliveira, R.; Paupy, C. Potential of Ae. albopictus as a bridge vector for enzootic pathogens at the urban-forest interface in Brazil. Emerg. Microbes Infect. 2018, 7, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.E.; Evans, B.R.; Zheng, W.; Obas, V.; Barrera-Martinez, L.; Egizi, A.; Zhao, H.; Caccone, A.; Powell, J.R. Human impacts have shaped historical and recent evolution in Ae. aegypti, the dengue and yellow fever mosquito. Evolution 2014, 68, 514–525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trpis, M.; Hausermann, W. Genetics of house-entering behaviour in East African populations of Ae. aegypti (L.) (Diptera: Culicidae) and its relevance to speciation. Bull. Entomol. Res. 1978, 68, 521–532. [Google Scholar] [CrossRef]
- McBride, C.S.; Baier, F.; Omondi, A.B.; Spitzer, S.A.; Lutomiah, J.; Sang, R.; Ignell, R.; Vosshall, L.B. Evolution of mosquito preference for humans linked to an odorant receptor. Nature 2014, 515, 222. [Google Scholar] [CrossRef] [PubMed]
- Powell, J.R.; Tabachnick, W.J. History of domestication and spread of Ae. aegypti—A Review. Mem. Inst. Oswaldo Cruz. 2013, 108, 11–17. [Google Scholar] [CrossRef] [PubMed]
- Williams, A.E.; Franz, A.W.E.; Reid, W.R.; Olson, K.E. Antiviral effectors and gene drive strategies for mosquito population suppression or replacement to mitigate arbovirus transmission by Ae. aegypti. Insects 2020, 11, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bennett, K.L.; McMillan, W.O.; Loaiza, J.R. Does Local Adaptation Impact on the Distribution of Competing Aedes Disease Vectors? Climate 2021, 9, 36. https://doi.org/10.3390/cli9020036
Bennett KL, McMillan WO, Loaiza JR. Does Local Adaptation Impact on the Distribution of Competing Aedes Disease Vectors? Climate. 2021; 9(2):36. https://doi.org/10.3390/cli9020036
Chicago/Turabian StyleBennett, Kelly L., William Owen McMillan, and Jose R. Loaiza. 2021. "Does Local Adaptation Impact on the Distribution of Competing Aedes Disease Vectors?" Climate 9, no. 2: 36. https://doi.org/10.3390/cli9020036