
Grassland Growth in Response to Climate Variability in the Upper Indus Basin, Pakistan

 ,
,  ,
,
Abstract
:1. Introduction
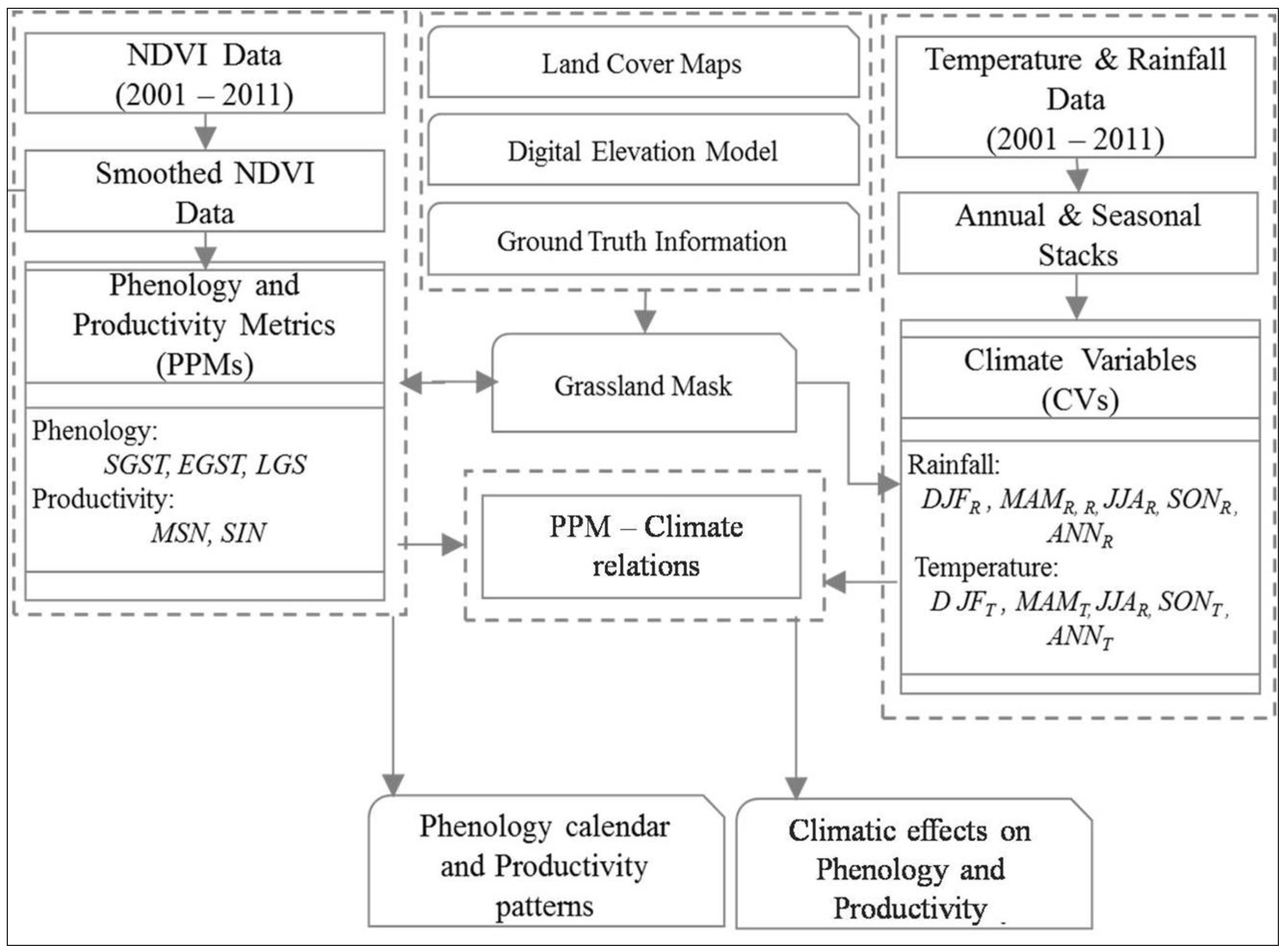
2. Materials and Methods
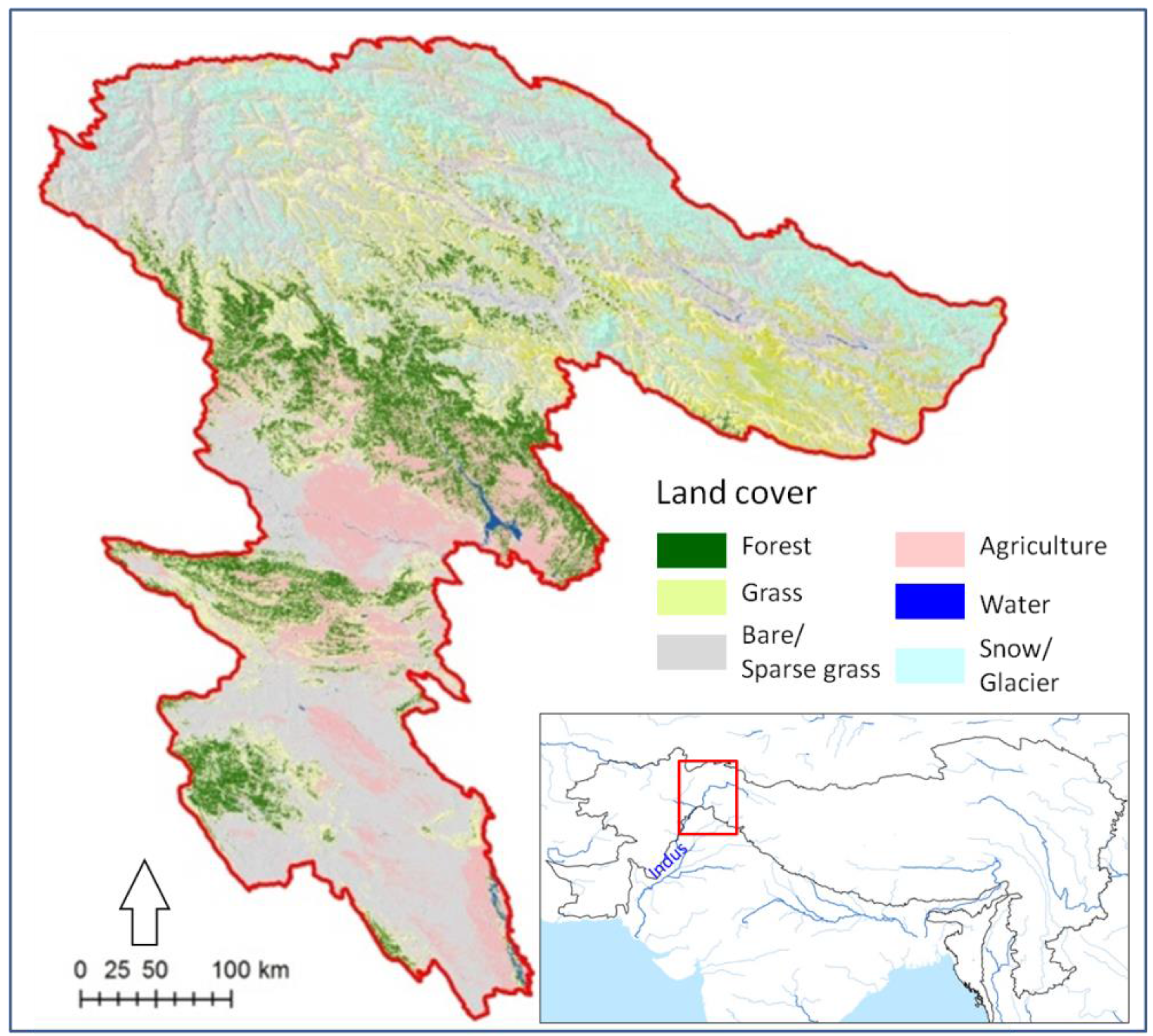
2.1. Study Area
2.2. Methods Summary


3. Results
3.1. Relationship between Phenology Metrics and Temperature


3.2. Relationship between Phenology Metrics and Rainfall
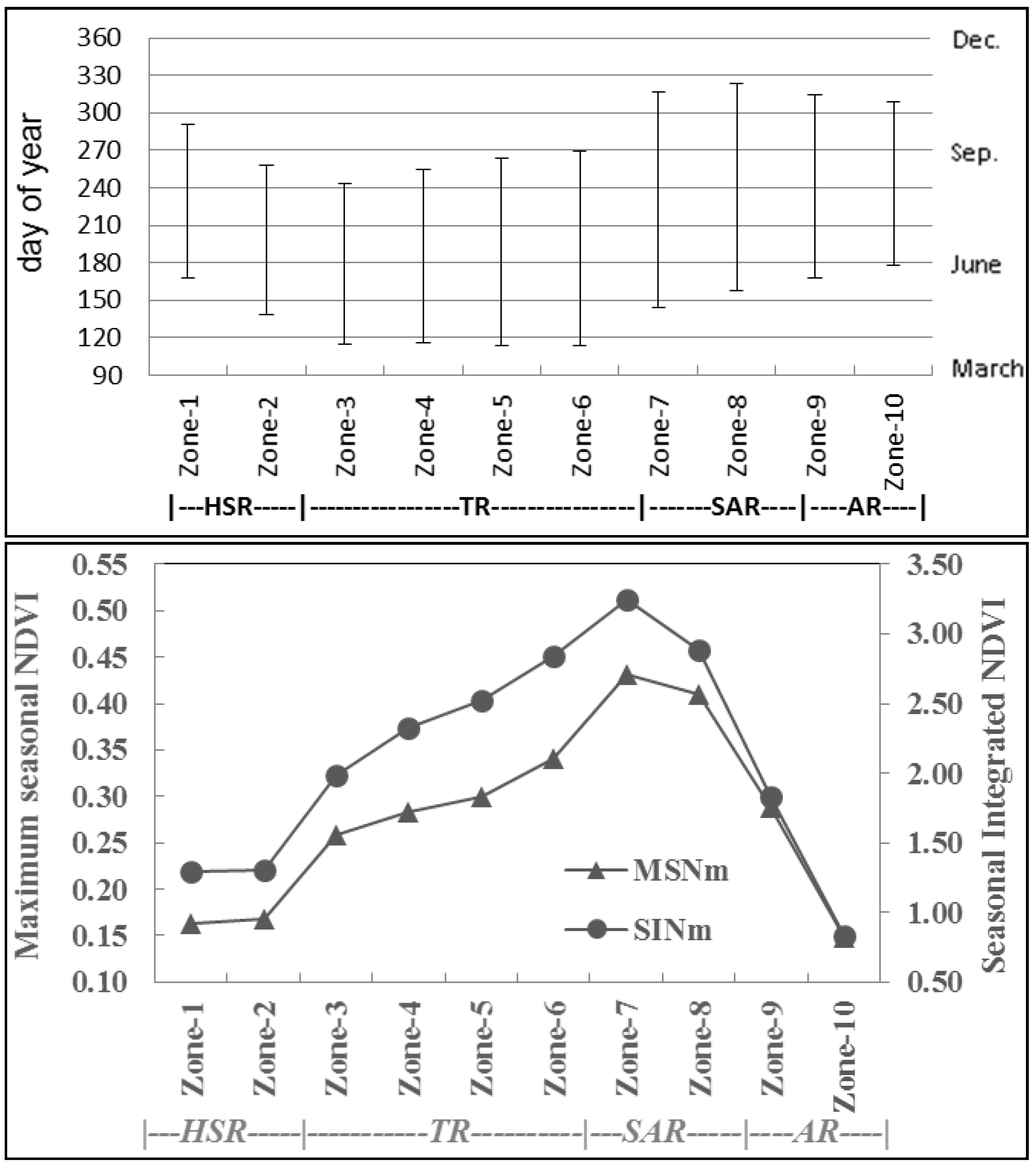
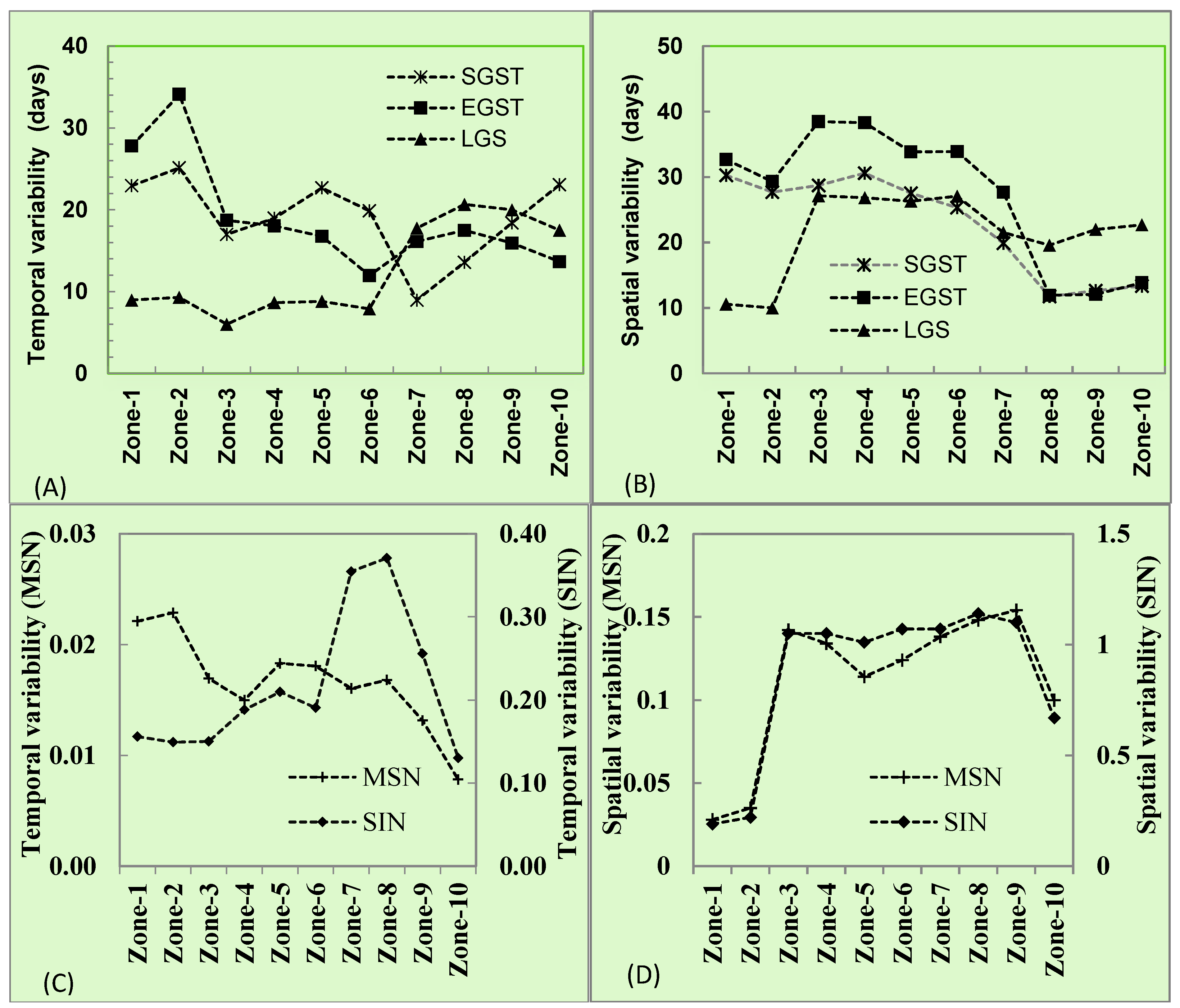
3.3. Phenology and Productivity Patterns


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elevation Zones | Temperature (°C) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DJFT | MAMT | JJAT | SONT | ANNT | |||||||||||
| µ | σs | σt | µ | σs | σt | µ | σs | σt | µ | σs | σt | µ | σs | σt | |
| Zone 1 | 21.6 | 1.7 | 1.4 | 40.1 | 2.1 | 1.8 | 37.9 | 1.9 | 1.5 | 35.8 | 1.6 | 0.9 | 33.8 | 1.8 | 0.7 |
| Zone 2 | 18.9 | 1.8 | 1.5 | 38.0 | 1.7 | 1.7 | 37.8 | 1.9 | 1.3 | 34.7 | 1.8 | 1.0 | 32.3 | 1.8 | 0.6 |
| Zone 3 | 11.6 | 4.4 | 1.3 | 31.9 | 4.0 | 1.9 | 36.6 | 4.0 | 0.8 | 30.3 | 3.5 | 0.8 | 27.6 | 4.0 | 0.7 |
| Zone 4 | 7.1 | 5.3 | 1.5 | 28.0 | 4.7 | 2.0 | 34.6 | 4.3 | 0.9 | 27.6 | 4.0 | 0.8 | 24.3 | 4.6 | 0.9 |
| Zone 5 | 2.2 | 4.7 | 1.7 | 24.4 | 5.0 | 2.3 | 32.8 | 4.6 | 1.3 | 24.8 | 4.4 | 0.9 | 21.1 | 4.7 | 1.0 |
| Zone-6 | −1.0 | 4.6 | 1.6 | 20.5 | 6.1 | 2.3 | 29.5 | 5.9 | 1.4 | 21.6 | 4.9 | 0.9 | 17.7 | 5.4 | 1.0 |
| Zone 7 | −8.0 | 4.4 | 1.2 | 9.6 | 6.3 | 1.9 | 19.2 | 6.0 | 1.3 | 13.5 | 4.7 | 0.8 | 8.6 | 5.3 | 0.9 |
| Zone 8 | −11.1 | 3.5 | 1.1 | 5.8 | 4.1 | 1.9 | 17.2 | 3.8 | 1.7 | 11.7 | 3.5 | 0.7 | 5.9 | 3.7 | 0.9 |
| Zone 9 | −12.9 | 2.8 | 1.1 | 4.2 | 3.8 | 1.8 | 16.9 | 3.5 | 2.0 | 10.7 | 3.3 | 0.8 | 4.7 | 3.4 | 0.9 |
| Zone 10 | −14.1 | 2.7 | 0.9 | 3.7 | 4.2 | 1.7 | 16.1 | 3.7 | 2.0 | 8.9 | 3.3 | 0.8 | 3.7 | 3.5 | 0.9 |
| Elevation Zones | Rainfall (mm) | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| DJFR | MAMR | JJAR | SONR | ANNR | |||||||||||
| µ | σs | σt | µ | σs | σt | µ | σs | σt | µ | σs | σt | µ | σs | σt | |
| Zone 1 | 30.2 | 8.9 | 18.3 | 32.7 | 8.8 | 14 | 66.2 | 9.3 | 37.7 | 16.4 | 4.4 | 9.6 | 145.6 | 7.8 | 79.6 |
| Zone 2 | 36.1 | 10 | 20.1 | 38.3 | 9.8 | 18.1 | 56 | 11.9 | 34.8 | 15.3 | 5.6 | 9.6 | 145.7 | 9.3 | 82.7 |
| Zone 3 | 43.7 | 17.2 | 19 | 43.6 | 16.7 | 18.8 | 45.7 | 20.9 | 26.7 | 15.5 | 7 | 9.8 | 148.6 | 15.4 | 74.3 |
| Zone 4 | 38.4 | 16.6 | 16.4 | 39.7 | 15.8 | 14.9 | 36.7 | 17.5 | 20.5 | 12.8 | 6 | 8.2 | 127.6 | 14 | 59.9 |
| Zone 5 | 34.1 | 13.7 | 15.2 | 36 | 12.7 | 15.4 | 27.7 | 11.6 | 20.5 | 10.6 | 4.1 | 7.5 | 108.3 | 10.5 | 58.6 |
| Zone 6 | 35.2 | 15.7 | 15.5 | 38.8 | 14.2 | 15.9 | 27.7 | 11.9 | 14.9 | 11.3 | 4.7 | 8.3 | 112.9 | 11.6 | 54.6 |
| Zone 7 | 47.8 | 20.5 | 19.3 | 53 | 19 | 19.5 | 46.1 | 23.8 | 19.5 | 16.8 | 7.2 | 11.9 | 163.7 | 17.6 | 70.2 |
| Zone 8 | 46.2 | 20 | 17.5 | 53 | 19.3 | 18.4 | 43.8 | 22.9 | 18.2 | 15.7 | 6.9 | 10.3 | 158.8 | 17.2 | 64.3 |
| Zone 9 | 37.4 | 16.3 | 14.1 | 45.7 | 17.1 | 17.3 | 34.6 | 17.3 | 14.8 | 12.5 | 5.1 | 7.8 | 130.2 | 14 | 54.1 |
| Zone 10 | 33 | 13.7 | 12.6 | 40.7 | 15.7 | 17.7 | 26.8 | 11.7 | 12.1 | 10.8 | 3.8 | 7 | 111.4 | 11.2 | 49.3 |

4. Discussion
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Harle, K.; Howden, S.; Hunt, L.; Dunlop, M. The potential impact of climate change on the Australian wool industry by 2030. Agric. Syst. 2007, 93, 61–89. [Google Scholar] [CrossRef]
- Paudel, K.P.; Andersen, P. Assessing rangeland degradation using multi temporal satellite images and grazing pressure surface model in Upper Mustang, Trans Himalaya, Nepal. Remote Sens. Environ. 2010, 114, 1845–1855. [Google Scholar] [CrossRef]
- Immerzeel, W.W.; Droogers, P.; de Jong, S.M.; Bierkens, M.F.P. Large-scale monitoring of snow cover and runoff simulation in Himalayan river basins using remote sensing. Remote Sens. Environ. 2009, 113, 40–49. [Google Scholar] [CrossRef]
- Vetter, S. Rangelands at equilibrium and non-equilibrium: Recent developments in the debate. J. Arid Environ. 2005, 62, 321–341. [Google Scholar] [CrossRef]
- Joshi, S.; Jasra, W.A.; Ismail, M.; Shrestha, R.M.; Yi, S.L.; Wu, N. Herders’ perceptions of and responses to climate change in northern pakistan. Environ. Manag. 2013, 52, 639–648. [Google Scholar] [CrossRef] [PubMed]
- Tucker, C.J.; Sellers, P.J. Satellite remote sensing of primary production. Int. J. Remote Sens. 1986, 7, 1395–1416. [Google Scholar] [CrossRef]
- Rigge, M.; Smart, A.; Wylie, B.; Gilmanov, T.; Johnson, P. Linking phenology and biomass productivity in South Dakota Mixed-Grass Prairie. Rangel. Ecol. Manag. 2013, 66, 579–587. [Google Scholar] [CrossRef]
- Wang, J.; Rich, P.M.; Price, K.P.; Kettle, W.D. Relations between NDVI and tree productivity in the central Great Plains. Int. J. Remote Sens. 2004, 25, 3127–3138. [Google Scholar] [CrossRef]
- Pettorelli, N.; Vik, J.O.; Mysterud, A.; Gaillard, J.M.; Tucker, C.J.; Stenseth, N.C. Using the satellite-derived NDVI to assess ecological responses to environmental change. Trends Ecol. Evol. 2005, 20, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.; Gao, X.; Ferreira, L. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar]
- Thein, T.R.; Watson, F.G.R.; Cornish, S.S.; Anderson, T.N.; Newman, W.B.; Lockwood, R.E. Chapter 7 vegetation dynamics of Yellowstone’s Grazing System. Terr. Ecol. 2009, 7, 113–133. [Google Scholar]
- Hangbin, Z.; Xiaoping, Y.; Jialin, L. MODIS data based NDVI Seasonal dynamics in agro-ecosystems of south bank Hangzhouwan bay. J. Agric. Res. 2011, 6, 4025–4033. [Google Scholar]
- Martínez, B.; Gilabert, M.A. Vegetation dynamics from NDVI time series analysis using the wavelet transform. Remote Sens. Environ. 2009, 113, 1823–1842. [Google Scholar] [CrossRef]
- Milesi, C.; Samanta, A.; Hashimoto, H.; Kumar, K.K.; Ganguly, S.; Thenkabail, P.S.; Srivastava, A.N.; Nemani, R.R.; Myneni, R.B. Decadal variations in NDVI and food production in India. Remote Sens. 2010, 2, 758–776. [Google Scholar] [CrossRef]
- Xu, L.; Myneni, R.B.; Chapin, F.S., III; Callaghan, T.V.; Pinzon, J.E.; Tucker, C.J.; Zhu, Z.; Bi, J.; Ciais, P.; Tømmervik, H.; et al. Temperature and vegetation seasonality diminishment over northern lands. Nat. Clim. Chang. 2013, 3, 581–586. [Google Scholar] [CrossRef]
- Bradley, B.A.; Jacob, R.W.; Hermance, J.F.; Mustard, J.F. A curve fitting procedure to derive inter-annual phenologies from time series of noisy satellite NDVI data. Remote Sens. Environ. 2007, 106, 137–145. [Google Scholar] [CrossRef]
- Nemani, R.R.; Keeling, C.D.; Hashimoto, H.; Jolly, W.M.; Piper, S.C.; Tucker, C.J.; Myneni, R.B.; Running, S.W. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science 2003, 300, 1560–1563. [Google Scholar] [CrossRef] [PubMed]
- Potter, C.S.; Klooster, S.; Brooks, V. Interannual Variability in Terrestrial Net Primary Production: Exploration of Trends and Controls on Regional to Global Scales. Ecosystems 1999, 2, 36–48. [Google Scholar] [CrossRef]
- Tucker, C.J.; Slayback, D.A.; Pinzon, J.E.; Los, S.O.; Myneni, R.B.; Taylor, M.G. Higher northern latitude normalized difference vegetation index and growing season trends from 1982 to 1999. Int. J. Biometeorol. 2001, 45, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Myneni, R.B.; Keeling, C.D.; Tucker, C.J.; Asrar, G.; Nemani, R.R. Increased plant growth in the northern high latitudes from 1981 to 1991. Nature 1997, 386, 698–702. [Google Scholar] [CrossRef]
- Reed, B.; Brown, J. Measuring phenological variability from satellite imagery. J. Veg. Sci. 1994, 5, 703–714. [Google Scholar] [CrossRef]
- Archer, D.R.; Fowler, H.J. Spatial and temporal variations in precipitation in the Upper Indus Basin, global teleconnections and hydrological implications. Hydrol. Earth Syst. Sci. 2004, 8, 47–61. [Google Scholar] [CrossRef]
- Champion, G.H.; Seth, S.K. Forest Types of Pakistan; Pakistani Forest Institute: Peshawar, Pakistan, 1965; p. 233. [Google Scholar]
- Butt, B.; Turner, M.D.; Singh, A.; Brottem, L. Use of MODIS NDVI to evaluate changing latitudinal gradients of rangeland phenology in Sudano-Sahelian West Africa. Remote Sens. Environ. 2011, 115, 3367–3376. [Google Scholar] [CrossRef]
- White, M.; Running, S.; Thornton, P. The impact of growing-season length variability on carbon assimilation and evapotranspiration over 88 years in the eastern US deciduous forest. Int. J. Biometeorol. 1999, 42, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Cleland, E.E.; Chuine, I.; Menzel, A.; Mooney, H.A.; Schwartz, M.D. Shifting plant phenology in response to global change. Trends Ecol. Evol. 2007, 22, 357–365. [Google Scholar] [CrossRef] [PubMed]
- Prince, S.D.; Goetz, S.J.; Goward, S.N. Monitoring primary production from Earth observing satellites. Water Air Soil Pollut. 1995, 82, 509–522. [Google Scholar] [CrossRef]
- Kariyeva, J.; van Leeuwen, W.J.D. Environmental Drivers of NDVI-Based Vegetation Phenology in Central Asia. Remote Sens. 2011, 3, 203–246. [Google Scholar] [CrossRef]
- Vrieling, A.; Beurs, K.M.; Brown, M.E. Variability of African farming systems from phenological analysis of NDVI time series. Clim. Chang. 2011, 109, 455–477. [Google Scholar] [CrossRef]
- Kreutzmann, H. Pastoral Practices and Their Transformation in the North-Western. 2004, 8, 54–88. [Google Scholar] [CrossRef]
- Nüsser, M.; Clemens, J. Impacts on mixed mountain agriculture in the Rupal Valley, Nanga Parbat, Northern Pakistan. Mountain Res. Dev. 2013, 16, 117–133. [Google Scholar] [CrossRef]
- Liu, J.; Tan, X.; J, W.; Ma, J.; Zhang, N. Comparative analysis between the 2010 severe drought in southwest China and typical drought disasters. China Water Resour. 2011, 9, 17–20. [Google Scholar]
- Palazzi, E.; Ahmad, T.; Paolo, C.; Elisa, V.; Antonello, P. Climatic characterization of Baltoro Glacier (Karakoram) and Northern Pakistan from in-situ stations. In Engineering Geology for Society and Territory; Lollino, G., Manconi, A., Clague, J., Shan, W., Chiarle, M., Eds.; Springer International Publishing: New York, NJ, USA, 2015; pp. 33–37. [Google Scholar]
- Xie, H.; Ringler, C.; Zhu, T.; Waqas, A. Droughts in Pakistan: A spatiotemporal variability analysis using the Standardized Precipitation Index. Water Int. 2013, 38, 620–631. [Google Scholar] [CrossRef]
- Fitter, A.; Fitter, R.; Harris, I.; Williamson, M. Relationships between first flowering date and temperature in the flora of a locality in central England. J. Func. Ecol. 1995, 296, 55–60. [Google Scholar] [CrossRef]
- Cook, B.I.; Wolkovich, E.M.; Parmesan, C. Divergent responses to spring and winter warming drive community level flowering trends. Proc. Natl. Acad. Sci. USA 2012, 109, 9000–9005. [Google Scholar] [CrossRef] [PubMed]
- Chuine, I. A unified model for budburst of trees. J. Theor. Biol. 2000, 207, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Laube, J.; Sparks, T.H.; Estrella, N.; Hofler, J.; Ankerst, D.P.; Menzel, A. Chilling outweighs photoperiod in preventing precocious spring development. Glob. Chang. Biol. 2014, 20, 170–182. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Pan, W. Relationships among phenological growing season, time-integrated normalized difference vegetation index and climate forcing in the temperate region of eastern China. Int. J. Climatol. 2002, 22, 1781–1792. [Google Scholar] [CrossRef]
- Higuchi, K.; Worthy, D.; Chan, D.; Shahshkov, A. Regional source/sink impact on the diurnal, seasonal and inter-annual variations in atmospheric CO2 at a boreal forest site in Canada. Tellus 2003, 55, 115–125. [Google Scholar] [CrossRef]
- Fowler, H.J.; Archer, D.R. Conflicting signals of climatic change in the upper Indus Basin. J. Clim. 2006, 19, 4276–4293. [Google Scholar] [CrossRef]
- Milly, P.C.D.; Betancourt, J.; Falkenmark, M.; Hirsch, R.M.; Zbigniew, W.; Lettenmaier, D.P.; Stouffer, R.J. Stationarity is dead: Whither water management? Science 2008, 319, 573–574. [Google Scholar] [CrossRef] [PubMed]
- Menzel, A.; Fabian, P. Growing season extended in Europe. Nature 1999. [Google Scholar] [CrossRef]
- Ding, M.; Zhang, Y.; Sun, X.; Liu, L.; Wang, Z.; Bai, W. Spatiotemporal variation in alpine grassland phenology in the Qinghai-Tibetan Plateau from 1999 to 2009. Chin. Sci. Bull. 2013, 58, 396–405. [Google Scholar] [CrossRef]
- Sardar, M.R. Indigenous Production and Utilization Systems in the High Altitude Alpine Pasture, Saif-ul- Maluk (NWFP), Pakistan; FAO and Pakistan Forest Institute: Peshawar, Pakistan, 1997. [Google Scholar]
- Seth, C.M. Biomass fluctuation in alpine pastures of Kashmir Himalaya. Ann. Arid Zone 1996, 35, 65–67. [Google Scholar]
- Chmielewski, F.-M.; Rötzer, T. Response of tree phenology to climate change across Europe. Agric. Forest Meteorol. 2001, 108, 101–112. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abbas, S.; Qamer, F.M.; Murthy, M.S.R.; Tripathi, N.K.; Ning, W.; Sharma, E.; Ali, G. Grassland Growth in Response to Climate Variability in the Upper Indus Basin, Pakistan. Climate 2015, 3, 697-714. https://doi.org/10.3390/cli3030697
Abbas S, Qamer FM, Murthy MSR, Tripathi NK, Ning W, Sharma E, Ali G. Grassland Growth in Response to Climate Variability in the Upper Indus Basin, Pakistan. Climate. 2015; 3(3):697-714. https://doi.org/10.3390/cli3030697
Chicago/Turabian StyleAbbas, Sawaid, Faisal M. Qamer, Manchiraju S.R. Murthy, Nitin K. Tripathi, Wu Ning, Eklabya Sharma, and Ghaffar Ali. 2015. "Grassland Growth in Response to Climate Variability in the Upper Indus Basin, Pakistan" Climate 3, no. 3: 697-714. https://doi.org/10.3390/cli3030697