
Multifaceted Linkages among Eco-Climatic Factors, Plankton Abundance, and Gonadal Maturation of Hilsa Shad, Tenualosa ilisha, Populations in Bangladesh

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Study Area
2.3. Data Collection
2.3.1. Determination of Plankton Abundance in the Water of Six Diverse Habitats of T. ilisha
2.3.2. Determination of Gut Plankton Abundance and Selective Feeding Indices of T. ilisha
2.3.3. Collection of Climatic Variables, Water Quality Parameters, and GSI Data
2.4. Statistical Analysis
3. Results
3.1. Assessment of Climatic and Water Quality Parameters in the Six Diverse Habitats of T. ilisha
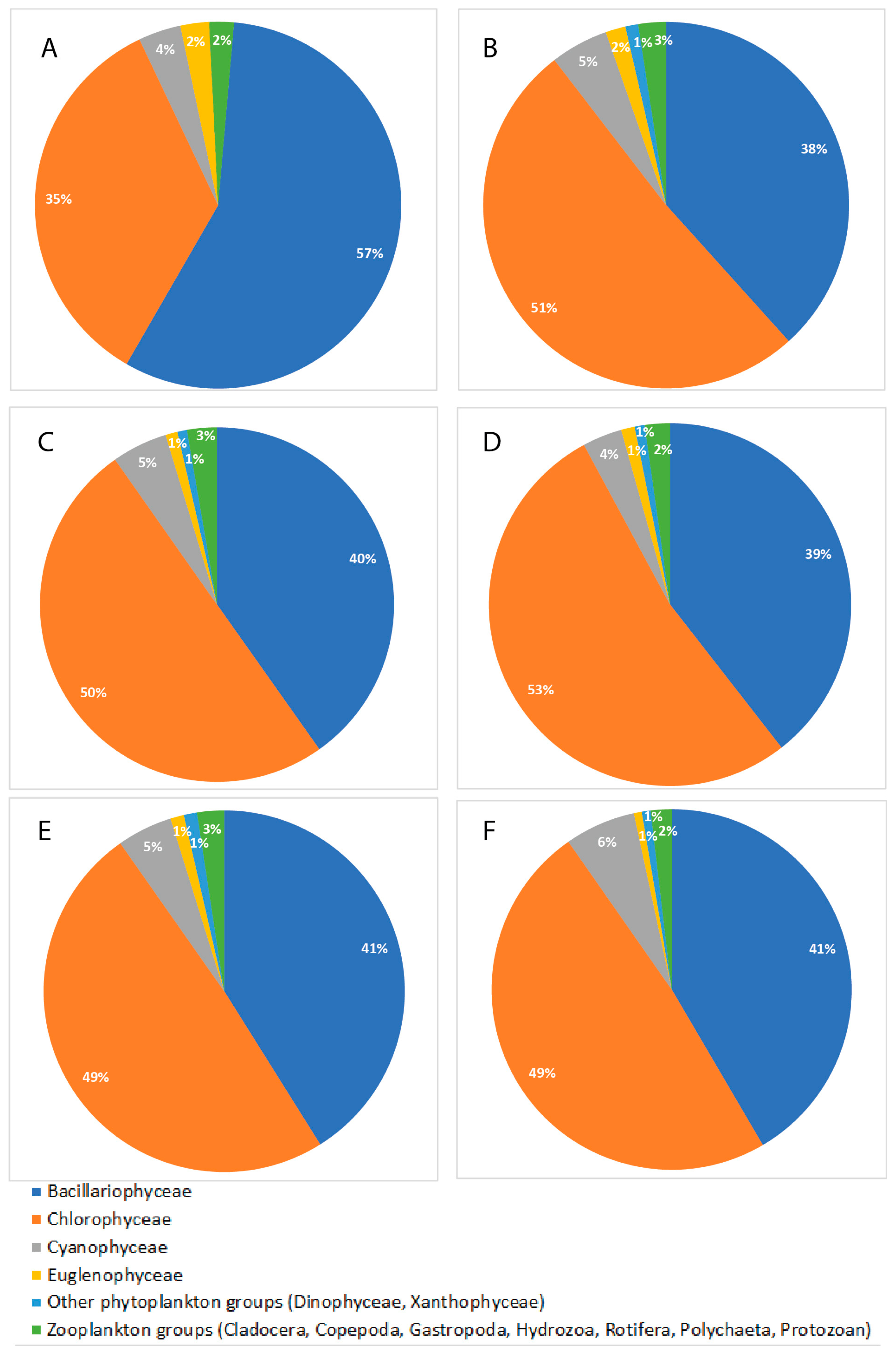
3.2. Determination of Plankton Abundance in the Six Diverse Habitats of T. ilisha
3.3. Determination of Gut Plankton Abundance and Selective Feeding Indices of T. ilisha
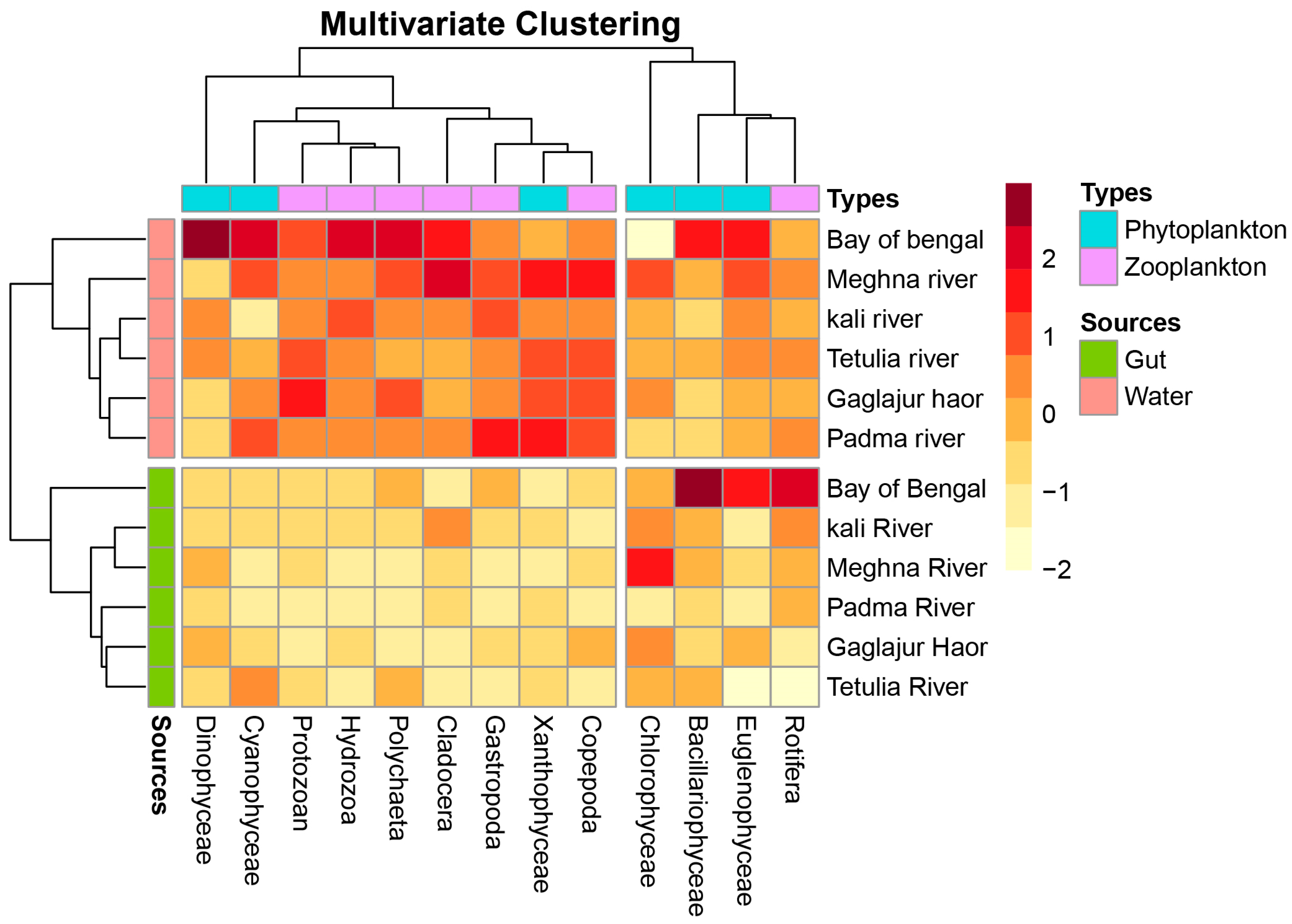
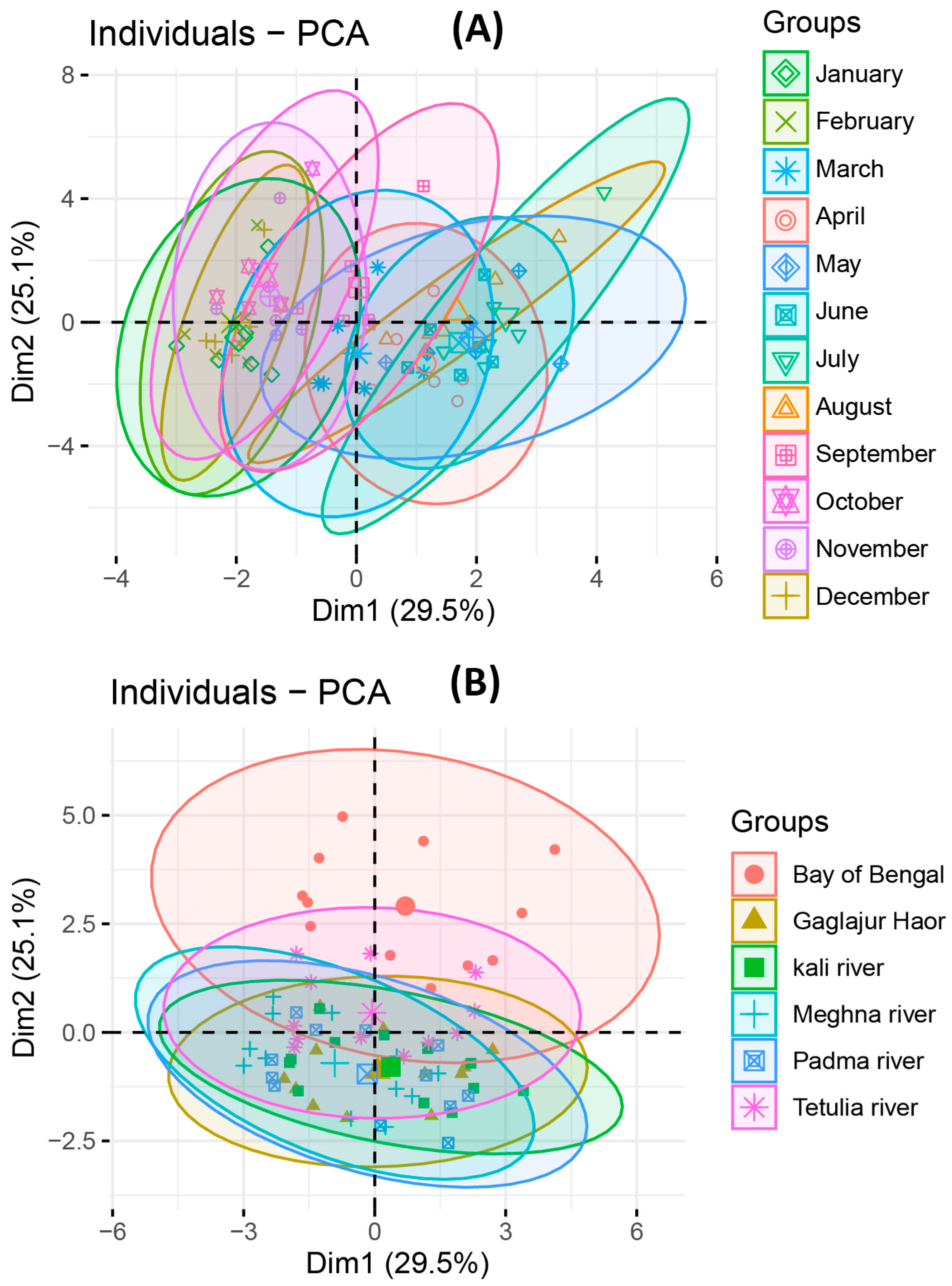
3.4. Multivariate Analysis among the Eco-Climatic Variables, Plankton Abundance in Water, Gut, and GSI of T. ilisha
3.5. Regression Analysis between Plankton Abundance in Gut and GSI of T. ilisha
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- DoF. National Fish Week Compendium; Department of Fisheries, Ministry of Fisheries and Livestock: Dhaka, Bangladesh, 2022; p. 160. Available online: https://ln.run/7GH6h (accessed on 19 July 2023). (In Bengali)
- Hossain, M.S.; Sarker, S.; Chowdhury, S.R.; Sharifuzzaman, S.M. Discovering spawning ground of Hilsa shad (Tenualosa ilisha) in the coastal waters of Bangladesh. Ecol. Model 2014, 282, 59–68. [Google Scholar] [CrossRef]
- Islam, M.M.; Islam, N.; Habib, A.; Mozumder, M.M.H. Climate change impacts on a tropical fishery ecosystem: Implications and societal responses. Sustainability 2020, 12, 7970. [Google Scholar] [CrossRef]
- Hossain, M.S.; Sharifuzzaman, S.M.; Rouf, M.A.; Pomeroy, R.S.; Hossain, M.D.; Chowdhury, S.R.; AftabUddin, S. Tropical hilsa shad (Tenualosa ilisha): Biology, fishery and management. Fish Fish. 2019, 20, 44–65. [Google Scholar] [CrossRef]
- Almukhtar, M.A.; Al-Mamun, M.A.; Rahman, M.M. Hilsa shad (Tenualosa ilisha): A review of its biology, fishery, and conservation status in Bangladesh. Rev. Fish. Sci. Aquac. 2016, 24, 265–276. [Google Scholar]
- Ahammad, A.K.S.; Hasan, N.A.; Haque, M.M.; Bashar, A.; Ahmed, M.B.U.; Alam, M.A.; Asaduzzaman, M.; Bashar, M.A.; Mahmud, Y. Environmental Factors and Genetic Diversity as Drivers of Early Gonadal Maturation: A Gonadosomatic Index Based Investigation on Indian Shad, Tenualosa ilisha Population of Bangladesh. Front. Mar. Sci. 2021, 8, 758868. [Google Scholar] [CrossRef]
- De, D.; Anand, P.S.S.; Sinha, S.; Suresh, V.R. Study on Preferred Food Items of Hilsa (Tenualosa Ilisha). Int. J. Agric. Food Sci. Technol. 2013, 4, 647–658. Available online: http://www.ripublication.com/ijafst.htm (accessed on 19 July 2023).
- Hasan, K.M.M.; Ahmed, Z.F.; Wahab, M.A.; Mohammed, E.Y. Food and Feeding Ecology of Hilsa (Tenualosa ilisha) in Bangladesh’s Meghna River Basin; IIED Working Paper; IIED: London, UK, 2016; Available online: https://shorturl.at/eyU27 (accessed on 19 July 2023).
- Bhaumik, U. Review of global studies on food, growth and maturity profile of Indian shad (Tenualosa ilisha). Int. J. Curr. Res. Acad. Rev. 2015, 3, 127–139. [Google Scholar] [CrossRef]
- Sarker, M.J.; Sarker, P.K.; Cahoon, L.B.; Dipty, A.K.; Bashar, M.A.; Hasan, M.M.; Mahmud, Y.; Sarker, M.M. Seasonal Variation in the Epibenthic Feeding Habits of Hilsa Shad (Tenualosa ilisha) in the Upper Meghna River Estuary, Bangladesh. Fishes 2023, 8, 335. [Google Scholar] [CrossRef]
- Mozumder, M.M.H.; Schneider, P.; Islam, M.M.; Deb, D.; Hasan, M.; Monzer, M.A.; Nur, A.A.U. Climate change adaptation strategies for small-scale Hilsa fishers in the coastal area of Bangladesh: Social, economic, and ecological perspectives. Front. Mar. Sci. 2023, 10, 1151875. [Google Scholar] [CrossRef]
- Barange, M.; Holsman, K.; Hollowed, A.; Ito, S.; Bograd, S.; Hazen, E.; King, J.; Mueter, F.; Perry, R.I. Climate change impacts, vulnerabilities and adaptations: North Pacific and Pacific Arctic marine fisheries. In Impacts of Climate Change on Fisheries and Aquaculture: Synthesis of Current Knowledge, Adaptation and Mitigation Options; Technical Paper 627; United Nations’ Food and Agriculture Organization: Rome, Italy, 2018. [Google Scholar]
- Dasgupta, S. Essential Fatty Acids. Br. Med. J. 2015, 2, 256–257. [Google Scholar] [CrossRef]
- Dilower, M.A.H.; Rahman, M.M.; Alam, M.J. Nutritional status of hilsa shad, Tenualosa ilisha (Hamilton, 1822) in the Bay of Bengal. Bangladesh J. Fish. 2023, 11, 1–6. [Google Scholar]
- Best of Farming. Hilsha Fish: The Best 10 Nutrition Facts. 2023. Available online: https://www.bestoffarming.com/hilsha-fish/ (accessed on 13 June 2023).
- Zhou, J.; Feng, P.; Li, Y.; Ji, H.; Gisbert, E. Effects of Dietary Lipid Levels on Growth and Gonad Development of Onychostoma macrolepis Broodfish. Fishes 2022, 7, 291. [Google Scholar] [CrossRef]
- Ray, S.K.; Dutta, S.; Pailan, G.H.; Suresh, V.R.; Dasgupta, S. Hormonal signatures of gonad maturity and seasonality of spawning in migrating hilsa. Tenualosa ilisha. Environ. Biol. Fish 2022, 105, 37–53. [Google Scholar] [CrossRef]
- Maulu, S.; Hasimuna, O.J.; Haambiya, L.H.; Monde, C.; Musuka, C.G.; Makorwa, T.H.; Munganga, B.P.; Phiri, K.J.; Nsekanabo, J.D.M. Climate Change Effects on Aquaculture Production: Sustainability Implications, Mitigation, and Adaptations. Front. Sustain. Food Syst. 2021, 5, 609097. [Google Scholar] [CrossRef]
- Miah, M.S. Climatic and anthropogenic factors changing spawning pattern and production zone of hilsa fishery in the Bay of Bengal. Weather. Clim. Extrem. 2015, 7, 109–115. [Google Scholar] [CrossRef]
- Hussain, M.G. Freshwater fishes of bangladesh: Fisheries, biodiversity and habitat. Aquat. Ecosyst. Health Manag. 2010, 13, 85–93. [Google Scholar] [CrossRef]
- Khan, M.R. Hilsa Production Growth Rate Declining. The Daily Star. 2023. Available online: https://www.thedailystar.net/news/bangladesh/news/hilsa-production-growth-rate-declining-3375126 (accessed on 19 July 2023).
- Prada-Ramallal, G.; Roque, F.; Herdeiro, M.T.; Takkouche, B.; Figueiras, A. Primary versus secondary source of data in observational studies and heterogeneity in meta-analyses of drug effects: A survey of major medical journals. BMC Med. Res. Methodol. 2018, 18, 97. [Google Scholar] [CrossRef] [PubMed]
- Valdez, A.S.; Raza, T.; Farolan, M.I.; Mendoza, C.I.; Perez, L.Q.; Peralta, J.F.; Valencia, R.I.; Lim, H.A.M.P. Primary and Secondary Data Collection to Conduct Researches, Write Thesis and Dissertation Amidst COVID-19 Pandemic: A Guidepost. In Proceedings of the 2nd International Symposium on Disaster Resilience and Sustainable Development; Lecture Notes in Civil Engineering. Pal, I., Kolathayar, S., Tawhidul Islam, S., Mukhopadhyay, A., Ahmed, I., Eds.; Springer: Singapore, 2023; Volume 294. [Google Scholar] [CrossRef]
- Creswell, J.W.; Plano Clark, V.L. Designing and Conducting Mixed Methods Research; Sage Publications: Thousand Oaks, CA, USA, 2017. [Google Scholar]
- Tashakkori, A.; Teddlie, C. Putting the human back in ‘Human research methodology’: The researcher in mixed methods research. J. Mix. Methods Res. 2010, 8, 758868. [Google Scholar] [CrossRef]
- Ivankova, N.V. Mixed Methods Applications in Action Research: From Methods to Community Action; Sage Publications: Thousand Oaks, CA, USA, 2014. [Google Scholar] [CrossRef]
- Uddin Ahmed, M.B.; Ahammad, A.K.; Shahjahan, M.; Rabbi, M.F.; Ashraful Alam, M.; Sakib, M.N.; Bashar, M.A.; Rahman, M.A.; Hossain, M.Y.; Mahmud, Y. Age, growth and maturity of the Indian Shad, Tenualosa ilisha through otolith examination from different habitats in Bangladesh. Egypt. J. Aquat. Biol. Fish 2020, 24, 343–359. [Google Scholar] [CrossRef]
- APHA (American Public Health Association). Standard Methods for the Examination of Water and Wastewater, 18th ed.; American Public Health Association: Washington, DC, USA, 1992. [Google Scholar]
- Botes, L. A guide to the common pelagic tunicates from South African waters. Afr. J. Mar. Sci. 2003, 25, 409–426. [Google Scholar]
- Mitra, A.; Flynn, K.J.; Burkholder, J.M.; Berge, T.; Calbet, A.; Raven, J.A.; Granéli, E.; Glibert, P.M.; Hansen, P.J.; Stoecker, D.K.; et al. The role of mixotrophic protists in the biological carbon pump. Biogeosciences Discuss. 2013, 10, 13535–13562. [Google Scholar] [CrossRef]
- McGaraghan, K. A guide to the identification of the diatoms of the United States. J. Phycol. 2018, 54, 1–16. [Google Scholar]
- Al-Kandari, M.; Al-Yamani, F.; Al-Rifaie, K. Marine Phytoplankton Atlas of Kuwait’s Waters; Kuwait Institute for Scientific Research: Safat, Kuwait, 2009. [Google Scholar]
- Ivlev, V.S. Experimental Ecology of the Feeding of Fishes; Yale University Press: New York, NY, USA, 1961; p. 302. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.programmingr.com/examples/neat-tricks/r-citation/ (accessed on 25 July 2023).
- Dag, A.; Hachman, A.; Singer, Y. Approximate leave-one-out for fast parameter tuning in high-dimensional latent variable models. In Proceedings of the 34th International Conference on Machine Learning, Sydney, Australia, 6–11 August 2017; Volume 70, pp. 927–936. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression; Sage Publications: Thousand Oaks, CA, USA, 2011. [Google Scholar]
- Galili, T. Dendextend: An R package for visualizing, adjusting, and comparing trees of hierarchical clustering. Bioinformatics 2015, 31, 3718–3720. [Google Scholar] [CrossRef] [PubMed]
- Lê, S.; Josse, J.; Husson, F.; Pagès, J. FactoMineR: An R package for multivariate analysis. J. Stat. Softw. 2008, 25, 1–18. [Google Scholar] [CrossRef]
- Kassambara, A.; Mundt, F. Factoextra: Extract and Visualize the Results of Multivariate Data Analyses. R Package Version 1.0.7. 2020. Available online: https://CRAN.R-project.org/package=factoextra (accessed on 31 March 2023).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. [Google Scholar]
- Yang, J.; Molfese, P.J.; Yu, Y.; Handwerker, D.A.; Chen, G.; Taylor, P.A.; Ejima, Y.; Wu, J.; Bandettini, P.A. Different activation signatures in the primary sensorimotor and higher-level regions for haptic three-dimensional curved surface exploration. NeuroImage 2021, 242, 118414. [Google Scholar] [CrossRef] [PubMed]
- Christensen, E.A.F.; Norin, T.; Tabak, I.; Van Deurs, M.; Behrens, J.W. Effects of temperature on physiological performance and behavioral thermoregulation in an invasive fish, the round goby. J. Exp. Biol. 2021, 224, jeb237669. [Google Scholar] [CrossRef] [PubMed]
- Pimolrat, P.; Whangchai, N.; Chitmanat, C.; Promya, J.; Lebel, L. Survey of Climate-Related Risks to Tilapia Pond Farms in Northern Thailand. Int. J. Geosci. 2013, 4, 54–59. [Google Scholar] [CrossRef]
- Santos, M.D.; Dickson, J.O.; Velasco, P.E.L. Mitigating the impacts of climate change: Philippine fisheries in focus. Fish People 2011, 9, 101–110. [Google Scholar]
- Sarkar, U.K.; Naskar, M.; Srivastava, P.K.; Roy, K.; Das Sarkar, S.; Gupta, S.; Bose, A.K.; Nandy, S.K.; Verma, V.K.; Sudheesan, D.; et al. Climato-environmental influence on breeding phenology of native catfishes in River Ganga and modeling species response to climatic variability for their conservation. Int. J. Biometeorol. 2019, 63, 991–1004. [Google Scholar] [CrossRef]
- Vass, K.K.; Das, M.K.; Srivastava, P.K.; Dey, S. Assessing the impact of climate change on inland fisheries in River Ganga and its plains in India. Aquat. Ecosyst. Health Manag. 2009, 12, 138–151. [Google Scholar] [CrossRef]
- Faruque, M.H.; Kabir, M.A. Climate change effects on aquaculture: A case study from northwestern Bangladesh. Int. J. Fish. Aquat. Stud. 2016, 4, 550–556. [Google Scholar]
- Soto-Zarazúa, G.M.; Rico-García, E.; Toledano-Ayala, M. Temperature effect on fish culture tank facilities inside greenhouse. Int. J. Phys. Sci. 2011, 6, 1039–1044. [Google Scholar]
- Sumant, M.M. Effect of Overall Heat Loss Coefficient on Performance of Circular Solar Collector for Close Water Open Air (Cwoa) Humidification Dehumidification Process. Int. J. Adv. Eng. Res. Dev. 2015, 119, 150–163. [Google Scholar] [CrossRef]
- Saber, A.; James, D.E.; Hayes, D.F.; Moret, G.J.M. Effects of seasonal fluctuations of surface heat flux and wind stress on mixing and vertical diffusivity of water column in deep lakes. Adv. Water Resour. 2018, 119, 150–163. [Google Scholar] [CrossRef]
- Chapanov, D. The impact of wind speed on water temperature in the Black Sea. J. Environ. Prot. Ecol. 2023, 24, 1–10. [Google Scholar]
- Didiharyono, D.; Sari, R.K.; Sari, A.K. The effect of wind speed on water temperature in the Bengawan Solo River, Indonesia. J. Phys. Conf. Ser. 2022, 2037, 1–10. [Google Scholar]
- Hossain, M.S.; Sharifuzzaman, S.M.; Chowdhury, S.R.; Sarker, S. Habitats across the life cycle of hilsa shad (Tenualosa ilisha) in aquatic ecosystem of Bangladesh. Fish. Manag. Ecol. 2016, 23, 450–462. [Google Scholar] [CrossRef]
- Nahar, J.; Hertini, E. Principal component analysis (pca) of phytoplankton community relations based on physical-chemical structures for supply chain management in the waters of the Bangka Bay region of west bay. Int. J. Supply Chain Manag. 2020, 9, 223–229. [Google Scholar]
- Kim, J.Y.; Lee, J.B.; Suh, Y.S. Oceanographic indicators for the occurrence of anchovy eggs inferred from generalized additive models. Fish. Aquat. Sci. 2020, 23, 1–14. [Google Scholar] [CrossRef]
- Anifowose, F.; Ajayi, O.; Fagorite, V. Seasonal Variability of Dissolved Oxygen in Relation To Temperature and Salinity in Gulf of Mexico, South of Cocodrie, Louisiana. Am. J. Innov. Res. Appl. Sci. 2020, 10, 237–241. [Google Scholar]
- Hossain, S.; Mofizur Rahman, M.; Akter, M.; Bhowmik, S. Species Composition and Abundance of Zooplankton Population in Freshwater Pond of Noakhali District, Bangladesh. World J. Fish Mar. Sci. 2015, 7, 387–393. [Google Scholar] [CrossRef]
- Xu, J.; Sun, J.; Beardall, J.; Gao, K. Lower Salinity Leads to Improved Physiological Performance in the Coccolithophorid Emiliania huxleyi, Which Partly Ameliorates the Effects of Ocean Acidification. Front. Mar. Sci. 2020, 7, 704. [Google Scholar] [CrossRef]
- Raven, J.A.; Gobler, C.J.; Hansen, P.J. Dynamic CO2 and pH levels in coastal, estuarine, and inland waters: Theoretical and observed effects on harmful algal blooms. Harmful Algae 2020, 91, 101594. [Google Scholar] [CrossRef]
- Hosen, H.A.; Pervin, R.; Shahriar, S.I. Changes in growth performances, survival rate and water quality parameter of pond on different stocking density of Gulsha Tengra (Mystus cavasius) in a monoculture system. Int. J. Fish. Aquat. Stud. 2017, 5, 52–56. [Google Scholar]
- Saalidong, B.M.; Aram, S.A.; Otu, S.; Lartey, P.O. Examining the dynamics of the relationship between water pH and other water quality parameters in ground and surface water systems. PLoS ONE 2022, 17, e0262117. [Google Scholar] [CrossRef]
- Abdullah, S.N.F.; Desa, S.M.; Awang, S.; Sulaiman, A.H.; Ismail, A.; Juahir, H.; Jusoh, A.M.; Yahaya, N.K.E.M.; Hashim, N.; San, L.Y.; et al. Impact of Flood Mitigation Project on the River Water Salinity. Trends Sci. 2022, 19, 5382. [Google Scholar] [CrossRef]
- Behara, A.; Vinayachandran, P.N.; Shankar, D. Influence of Rainfall Over Eastern Arabian Sea on Its Salinity. J. Geophys. Res. Ocean. 2019, 124, 5003–5020. [Google Scholar] [CrossRef]
- Sultana, T.; Haque, M.; Salam, M.; Alam, M. Effect of aeration on growth and production of fish in intensive aquaculture system in earthen ponds. J. Bangladesh Agric. Univ. 2017, 15, 113–122. [Google Scholar] [CrossRef]
- Sun, Y.; Zhang, W.; Peng, H.; Zhou, F.; Jiang, A.; Chen, X.; Wang, H. The Impacts of Climate Change on the Hydrological Process and Water Quality in the Three Gorges Reservoir Area, China. Water 2023, 15, 1542. [Google Scholar] [CrossRef]
- Asaduzzaman, M.; Akter, S.; Hoque, N.F.; Shakil, A.; Noor, A.R.; Akter, M.N.; Rahman, M.M. Multifaceted linkages among eco-physiological factors, seasonal plankton dynamics and selective feeding behavior of the green mussel (Perna viridis) in the south-east coast of the Bay of Bengal. J. Sea Res. 2020, 164, 101933. [Google Scholar] [CrossRef]
- Brasil, J.; Santos, J.B.O.; Sousa, W.; Menezes, R.F.; Huszar, V.L.M.; Attayde, J.L. Rainfall leads to habitat homogenization and facilitates plankton dispersal in tropical semiarid lakes. Aquat. Ecol. 2020, 54, 225–241. [Google Scholar] [CrossRef]
- Sallam, G.A.H.; Elsayed, E.A. Estimating the impact of air temperature and relative humidity change on the water quality of Lake Manzala, Egypt. J. Nat. Resour. Dev. 2015, 5, 76–87. [Google Scholar] [CrossRef]
- Sarkar, U.K.; Roy, K.; Karnatak, G.; Naskar, M.; Puthiyottil, M.; Baksi, S.; Lianthuamluaia, L.; Kumari, S.; Ghosh, B.D.a.s.; Das, B.K. Reproductive environment of the decreasing Indian river shad in Asian inland waters: Disentangling the climate change and indiscriminative fishing threats. Environ. Sci. Pollut. Res. 2021, 28, 30207–30218. [Google Scholar] [CrossRef] [PubMed]
- Alfonso, M.B.; Zunino, J.; Piccolo, M.C. Impact of water input on plankton temporal dynamics from a managed shallow saline lake. Ann. De Limnol. 2017, 53, 391–400. [Google Scholar] [CrossRef]
- Kürten, B.; Zarokanellos, N.D.; Devassy, R.P.; El-Sherbiny, M.M.; Struck, U.; Capone, D.G.; Schulz, I.K.; Al-Aidaroos, A.M.; Irigoien, X.; Jones, B.H. Seasonal modulation of mesoscale processes alters nutrient availability and plankton communities in the Red Sea. Prog. Oceanogr. 2019, 173, 238–255. [Google Scholar] [CrossRef]
- Khoshnevis Yazdi, S.; Shakouri, B. The Effects of Climate Change on Aquaculture. Int. J. Environ. Sci. Dev. 2010, 1, 378–382. [Google Scholar] [CrossRef]
- Cabrerizo, M.J.; Marañón, E.; Fernández-González, C.; Alonso-Núñez, A.; Larsson, H.; Aranguren-Gassis, M. Temperature Fluctuation Attenuates the Effects of Warming in Estuarine Microbial Plankton Communities. Front. Mar. Sci. 2021, 8, 656282. [Google Scholar] [CrossRef]
- Hastuti, A.W.; Pancawati, Y.; Surana, I.N. The abundance and spatial distribution of plankton communities in Perancak Estuary, Bali. IOP Conf. Ser. Earth Environ. Sci. 2018, 176, 012042. [Google Scholar] [CrossRef]
- Dupuis, A.P.; Hann, B.J. Warm spring and summer water temperatures in small eutrophic lakes of the Canadian prairies: Potential implications for phytoplankton and zooplankton. J. Plankton Res. 2009, 31, 489–502. [Google Scholar] [CrossRef]
- Ahmed, K.K.U.; Ahamed, S.U.; Hossain, M.R.A.; Ahmed, T.; Barman, S. Quantitative and qualitative assessment of plankton: Some ecological aspect and water quality parameters of the river Meghna, Bangladesh. Bangldesh J. Fish. Res 2003, 7, 131–140. [Google Scholar]
- Ahmed, M.; Islam, R.; Hadiuzzaman, M. Emergency response during disastrous situations in densely populated urban areas: A GIS-based approach. Hydrology 2018, 10, 86–95. [Google Scholar] [CrossRef]
- Huang, X.; Lu, D.; Ricciuto, D.M.; Hanson, P.J.; Richardson, A.D.; Lu, X.; Weng, E.; Nie, S.; Jiang, L.; Hou, E.; et al. A model-independent data assimilation (MIDA) module and its applications in ecology. Geosci. Model Dev. 2021, 14, 5217–5238. [Google Scholar] [CrossRef]
- Mozumder, M.S.I.; De Wever, H.; Volcke, E.I.; Garcia-Gonzalez, L. A robust fed-batch feeding strategy independent of the carbon source for optimal polyhydroxybutyrate production. Process Biochem. 2014, 49, 365–373. [Google Scholar] [CrossRef]
- De los Ríos, P. Richness and distribution of zooplanktonic crustacean species in Chilean Andes mountains and southern Patagonia shallow ponds. Pol. J. Environ. 2005, 14, 817–822. [Google Scholar]
- Vignatti, A.M.; Paggi, J.C.; Cabrera, G.C.; Echaniz, S.A. Zooplankton diversity and its relationship with environmental changes after the filling of a temporary saline lake in the semi-arid region of La Pampa, Argentina. Lat. Am. J. Aquat. Res. 2012, 40, 1005–1016. [Google Scholar] [CrossRef]
- Starks, E.; Cooper, R.; Leavitt, P.R.; Wissel, B. Effects of drought and pluvial periods on fish and zooplankton communities in prairie lakes: Systematic and a systematic responses. Glob. Chang. Biol. 2014, 20, 1032–1042. [Google Scholar] [CrossRef]
- Echaniz, S.A.; Vignatti, A.M. Seasonal variation and influence of turbidity and salinity on the zooplankton of a saline lake in central Argentina. Lat. Am. J. Aquat. Res. 2011, 39, 306–315. [Google Scholar] [CrossRef]
- Echaniz, S.A.; Cabrera, G.C.; Vignatti, A.M. Limnological parameters and population structure of Artemia persimilis Piccinelli and Prosdocimi, 1968 (Crustacea, Anostraca) in La Amarga, a Hypersaline Lake of La Pampa (Argentina). Res. Zool. 2015, 5, 25–31. [Google Scholar]
- Iglesias, C.; Mazzeo, N.; Meerhoff, M.; Lacerot, G.; Clemente, J.M.; Scasso, F.; Kruk, C.; Goyenola, G.; García-Alonso, J.; Amsinck, S.L.; et al. High predation is ofkey importance for dominance of small-bodied zooplankton in warm shallow lakes: Evidence from lakes, fish exclosures and surface sediments. Hydrobiologia 2011, 667, 133–147. [Google Scholar] [CrossRef]
- Vignatti, L.; Garcia, A.M.; Garcia, V.; Garcia, J.; Garcia, R. Patterns of Plankton Response to Predation Pressure from Omnivorous Planktivorous Fish in Eutrophic Environments. J. Aquat. Ecol. 2012, 42, 201–215. [Google Scholar]
- Borics, G.; Tóthmérész, B.; Lukács, B.A.; Várbíró, G. Functional groups of phytoplankton shaping diversity of shallow lake ecosystems. Hydrobiologia 2012, 698, 251–262. [Google Scholar] [CrossRef]
- Hossain, A.; Knorr, G.; Jokat, W.; Lohmann, G. Opening of the Fram Strait led to the establishment of a modern-like three-layer stratification in the Arctic Ocean during the Miocene. Arktos—J. Arct. Geosci. 2021, 1, 2477–2499. [Google Scholar] [CrossRef]
- Harrod, C.; Griffiths, A.M.; Limburg, K.E. Using otolith chemistry to reconstruct habitat use and migration patterns in fishes. Fishes Estuaries 2018, 101, 191–214. [Google Scholar]
- Sriyasak, P.; Chitmanat, C.; Whangchai, N.; Promya, J.; Lebel, L. Effects of Temperature upon Water Turnover in Fish Ponds in Northern Thailand. Int. J. Geosci. 2013, 4, 18–23. [Google Scholar] [CrossRef]
- Nair, N.B.; Nancollas, G.H. Observations on the biology of the Hilsa, Hilsa ilisha (Hamilton) with special reference to its migration and spawning in the river Hooghly. Indian J. Fish. 1958, 5, 68–116. [Google Scholar]
- Mondal, P. The breeding biology of Hilsa shad, Tenualosa ilisha (Hamilton) in the Ganga River, West Bengal, India. Iran. J. Fish. Sci. 2012, 11, 1–13. [Google Scholar]
- Kay, S.; Caesar, J.; Wolf, J.; Bricheno, L.; Nicholls, R.J.; Islam, A.S.; Haque, A.; Pardaens, A.; Lowe, J.A. Modelling the increased frequency of extreme sea levels in the Ganges-Brahmaputra-Meghna delta due to sea level rise and other effects of climate change. Environ. Sci. Process. Impacts 2018, 20, 1312–1325. [Google Scholar] [CrossRef]
- Rahman, M.S. The Advantages and Disadvantages of Using Qualitative and Quantitative Approaches and Methods in Language “Testing and Assessment” Research: A Literature Review. J. Educ. Learn. 2017, 6, 102–112. [Google Scholar] [CrossRef]
- Mahdi, S.S. Climate Change and Agriculture in India: Impact and Adaptation; Springer: Berlin/Heidelberg, Germany, 2019; pp. 1–262. [Google Scholar] [CrossRef]
- Li, X.; Wei, P.; Liu, S.; Tian, Y.; Ma, H.; Liu, Y. Photoperiods affect growth, food intake and physiological metabolism of juvenile European Sea Bass (Dicentrachus labrax L.). Aquac. Rep. 2021, 20, 100656. [Google Scholar] [CrossRef]
- Castro, J.S.; Braz-Mota, S.; Campos, D.F.; Souza, S.S.; Val, A.L. High Temperature, pH, and Hypoxia Cause Oxidative Stress and Impair the Spermatic Performance of the Amazon Fish Colossoma macropomum. Front. Physiol. 2020, 11, 772. [Google Scholar] [CrossRef]
- Muslim, M.; Fitrani, M.; Afrianto, A.M. The Effect of Water Temperature on Incubation Period, Hatching Rate, Normalities of The Larvae and Survival Rate of Snakehead Fish Channa striata. Aquac. Indones. 2018, 19, 90. [Google Scholar] [CrossRef]
- Bladon, A.; Myint, K.T.; Ei, T.; Khine, M.; Aye, P.T.; Thwe, T.L.; Leemans, K.; Soe, K.M.; Akester, M.; Merayo, E. Spawning seasonality of hilsa (Tenualosa ilisha) in Myanmar’s Ayeyarwady Delta; International Institute for Environment and Development (IIED): London, UK, 2019. [Google Scholar]
- Villalejo-Fuerte, M.; Muñetón-Gómez, M.D.S.; Gárate-Lizárraga, I.; García-Domínguez, F. Gut content, phytoplankton abundance and reproductive season in the black oyster (Hyotissa hyotis, Linné, 1758) at Isla Espíritu Santo, Gulf of California. J. Shellfish Res. 2005, 24, 185–190. [Google Scholar] [CrossRef]
- Kimbonguila, A.; Matos, L.; Petit, J.; Scher, J.; Nzikou, J.-M. Effect of Physical Treatment on the Physicochemical, Rheological and Functional Properties of Yam Meal of the Cultivar “Ngumvu” From Dioscorea Alata L. of Congo. Int. J. Recent Sci. Res. 2019, 10, 30693–30695. [Google Scholar] [CrossRef]
- Ahsan, D.A.; Kabir, A.N.; Rahman, M.M.; Mahabub, S.; Yesmin, R.; Faruque, M.H.; Naser, M.N. Plankton composition, abundance and diversity in hilsa (Tenualosa ilisha) migratory rivers of Bangladesh during spawning season. Dhaka Univ. J. Biol. Sci. 2012, 21, 177–189. [Google Scholar] [CrossRef]
- Pinto-Coelho, R.M.; Giani, A.; Morais, C.A., Jr.; Carvalho, E.R., Jr.; Bezerra-Neto, J.F. The nutritional status of zooplankton in a tropical reservoir: Effects of food quality and community structure. Braz. J. Biol. (Rev. Brasleira Biol.) 2005, 65, 313–324. [Google Scholar] [CrossRef]
- Meghna, B. The Biophysical Assessments of the Hilsa Fish (Tenualosa ilisha) Habitat in the Lower. October 2015. Available online: https://www.iied.org/sites/default/files/pdfs/migrate/16605IIED.pdf (accessed on 20 July 2023).
- Yoahida, T.; Liong, C.F.; Majid, A.M.; Toda, T.; Othman, B.H.J. Temperature effects on the egg development time and hatching success of three Acartia species (Copepoda: Calanoida) from the Strait of Malacca. Zool. Stud. 2012, 51, 644–654. [Google Scholar]
- Haque, M.F.; Begum, N.; Islam, M.S. Seasonal Variations in Phytoplankton and Zooplankton Population in Relation To Some Environmental Factors in the Tidal Sangu River in Chittagong in Bangladesh. J. Sylhet Agril. Univ. 2015, 2, 209–219. [Google Scholar]
- Jerin, S.; Gani, M.A.; Khondker, M. Dynamics of Environmental Factors in Relation to Phytoplankton Species in a Pond of Old Dhaka, Bangladesh. J. Asiat. Soc. Bangladesh Sci. 2016, 42, 169–175. [Google Scholar] [CrossRef]
- Claude, E. Boyd and Frank Lichtkoppler. Water Quality Management In Pond Fish Culture, International Center for Aquaculture Agricultural Experiment Station. Res. Dev. Ser. 1979, 22, 3–4. [Google Scholar]
- Puccinelli, E.; Sardenne, F.; Pecquerie, L.; Fawcett, S.E.; Machu, E.; Soudant, P. Omega-3 Pathways in Upwelling Systems: The Link to Nitrogen Supply. Front. Mar. Sci. 2021, 8, 664601. [Google Scholar] [CrossRef]
- Rajendran, R.; Ramesh, S.; Roy, S.D.; Haq, M.A.B.; Ramalingam, V. Abundance of anemone fishes in North Bay Island and mass culture of live food organisms for their larval rearing. J. Fish. 2014, 2, 173. [Google Scholar] [CrossRef]
- Jayalakshmi, T.; Manoharan, N.; Santhanam, P.; Ananth, S.; Dinesh Kumar, S.; Nandakumar, R.; Kaviyarasan, M. Water quality and proximate composition of zooplankton of Muthukkuda coast of Palk Bay, southeastern India. J. Mar. Biol. Assoc. India 2017, 59, 25–30. [Google Scholar] [CrossRef]
- Meynier, L.; Morel, P.C.H.; Mackenzie, D.D.S.; Macgibbon, A.; Chilvers, B.L.; Duignan, P.J. Proximate composition, energy content, and fatty acid composition of marine species from Campbell Plateau, New Zealand. N. Z. J. Mar. Freshw. Res. 2008, 42, 425–437. [Google Scholar] [CrossRef]
- Arfiati, D.; Kertikasari, F.S.; Cokrowati, N.; Puspitasari, A.W. Gut content analysis of tilapia (Oreochromis niloticus) from Jagir River, Surabaya City, East java. AIP Conf. Proc. 2019, 2120, 040007. [Google Scholar] [CrossRef]
- Hasan, K.; Wahab, A.; Ahmed, Z.F.; Mohammed, E.Y. Food and Feeding Ecology with Special Reference to Food Preference of Hilsa Tenualosa ilisha in Meghna River Basin of Bangladesh; Working Paper; International Institute for Environment and Development (IIED): London, UK, 2016. [Google Scholar] [CrossRef]
- De, S.; Chakraborty, S.K.; Ghosh, A. Plankton composition in the water and gut of hilsa (Tenualosa ilisha) and its feeding preferences. J. Aquat. Ecol. 2013, 42, 201–215. [Google Scholar]
- Rollwagen-Bollens, G.; Connelly, K.A.; Bollens, S.M.; Zimmerman, J.; Coker, A. Nutrient Control of Phytoplankton Abundance and Biomass, and Microplankton Assemblage Structure in the Lower Columbia River (Vancouver, Washington, USA). Water 2022, 14, 1599. [Google Scholar] [CrossRef]
- Shaha, D.C.; Hasan, J.; Kundu, S.R.; Yusoff, F.M.; Salam, M.A.; Khan, M.; Haque, F.; Ahmed, M.; Rahman, M.J.; Wahab, M.A. Dominant phytoplankton groups as the major source of polyunsaturated fatty acids for hilsa (Tenualosa ilisha) in the Meghna estuary Bangladesh. Sci. Rep. 2022, 12, 20980. [Google Scholar] [CrossRef]
- Segner, H. Reproductive and Developmental Toxicity in Fishes. In Reproductive and Developmental Toxicology; Elsevier: Amsterdam, The Netherlands, 2011. [Google Scholar] [CrossRef]
- Jones, J. The effects of environmental factors on hilsa fish reproduction in the rivers of Bangladesh. J. Fish. Aquat. Sci. 2012, 7, 1–10. [Google Scholar]
- Holmgren, T. Environmental factors affecting hilsa fish reproduction in the rivers of Bangladesh. J. Fish Biol. 1994, 45, 1–10. [Google Scholar]
- Mohammed, E.Y.; Hasan, K.M.M.; Wahab, A.; Ahmed, Z.F. Physical, chemical and biological assessments of the Hilsa fish (Tenualosa ilisha) habitat in the lower Meghna, Bangladesh. J. Fish Biol. 2015; Submitted. Available online: https://www.iied.org/sites/default/files/pdfs/migrate/16609IIED.pdf(accessed on 19 July 2023).
- Liu, G.; He, W.; Cai, S. Seasonal variation of dissolved oxygen in the southeast of the Pearl River Estuary. Water 2020, 12, 2475. [Google Scholar] [CrossRef]
- Brander, K. Impacts of climate change on fisheries. J. Mar. Syst. 2010, 79, 389–402. [Google Scholar] [CrossRef]
- Moniruzzaman, M.; Flura; Alam, M.A.; Rahman, M.H.; Hasan, K.R.; Lima, R.A.; Mahmud, Y. Nutrient and Plankton Abundance in the Major Nursery Grounds of Hilsa (Tenualosa ilisha) in Perspective of Water Quality Factors. Am. J. Biosci. Bioinf. 2023, 2, 1–13. [Google Scholar] [CrossRef]
- Volkoff, H.; Rønnestad, I. Effects of temperature on feeding and digestive processes in fish. Temperature 2020, 7, 307–320. [Google Scholar] [CrossRef]
- Abedin, M.Z.; Rahman, M.M.; Alam, M.J. Effect of water quality on the growth and survival of Indian major carp, Labeo rohita (Hamilton). Int. J. Fish. Aquat. Stud. 2017, 5, 1–6. [Google Scholar]
- Adeogun, O.; Fafioye, O.O.; Olaleye, B.A.; Ngobili, G.O. The relationship between some physicochemical parameters and plankton composition on fish production in ponds. In Proceedings of the 19th Annual Conference of the Fisheries Society of Nigeria (FISON), Lagos, Ilorin, Nigeria, 29 November–3 December 2005; Volume 4. Available online: http://hdl.handle.net/1834/21775 (accessed on 19 July 2023).
- Pilkaitytë, R.; Razinkovas-Baziukas, A.; Olenin, S. The impact of salinity increase on the zooplankton community of a coastal lake undergoing restoration measures (Curonian Lagoon, Baltic Sea). Oceanologia 2016, 58, 1–10. [Google Scholar]
- Rahman, M.M.; Islam, M.A.; Azam, M.S. Relationship between phytoplankton abundance and gonadal maturation of Hilsa fish (Tenualosa ilisha) in Bangladesh. J. Fish. Aquat. Sci. 2020, 15, 223–230. [Google Scholar]
- Ahmed, M.K.; Islam, M.M.; Akhtaruzzaman, M. Influence of phytoplankton abundance on gonadal maturation of Hilsa fish (Tenualosa ilisha) in the Meghna River, Bangladesh. J. Appl. Fish. Aquat. Res. 2021, 9, 24–32. [Google Scholar]
- Hossain, M.Y.; Nahiduzzaman, M.; Hasanuzzaman, A.F.M. Conservation of Hilsa (Tenualosa ilisha) in Bangladesh: Challenges and strategies. Int. J. Fish. Aquat. Stud. 2018, 6, 57–65. [Google Scholar]
- Kabir, M.R.; Khan, M.M.R.; Islam, M.A. Conservation of Hilsa (Tenualosa ilisha) in Bangladesh: Progress, challenges, and future prospects. J. Fish. 2020, 8, 647–656. [Google Scholar]
- Sarker, M.S.; Uddin, M.J.; Rahman, M.S. Implications of climate change on phytoplankton dynamics and fish reproductive success in the Bay of Bengal. Clim. Chang. Environ. Sustain. 2022, 10, 179–191. [Google Scholar]
- Siddique, M.A.B.; Ahammad, A.K.S.; Mahalder, B.; Alam, M.M.; Hasan, N.A.; Bashar, A.; Biswas, J.C.; Haque, M.M. Perceptions of the Impact of Climate Change on Performance of Fish Hatcheries in Bangladesh: An Empirical Study. Fishes 2022, 7, 270. [Google Scholar] [CrossRef]
- Siddique, M.A.B.; Ahammad, A.K.S.; Bashar, A.; Hasan, N.A.; Mahalder, B.; Alam, M.M.; Biswas, J.C.; Haque, M.M. Impacts of climate change on fish hatchery productivity in Bangladesh: A critical review. Heliyon 2022, 8, e11951. [Google Scholar] [CrossRef]
- Akhter, S.; Islam, M.R.; Hossain, M.Y. Nutritional composition of phytoplankton and its effects on reproductive hormones and gamete development in Hilsa fish (Tenualosa ilisha). Aquaculture 2017, 468, 316–324. [Google Scholar]
- Nahar, T.; Sarker, M.S.; Rahman, M.S. Influence of phytoplankton species on gonadal maturation of Hilsa fish (Tenualosa ilisha) in the Meghna River, Bangladesh. Aquat. Ecosyst. Health Manag. 2019, 22, 29–40. [Google Scholar]
- Rahman, M.A.; Hague, M.M.; Islam, M.S. Nutritional composition of phytoplankton and its role in gonadal maturation of Hilsa fish (Tenualosa ilisha) in the Padma River, Bangladesh. Aquat. Biol. 2023, 31, 45–56. [Google Scholar]
- Siddique, M.A.B.; Mahalder, B.; Haque, M.M.; Bashar, A.; Hasan, M.M.; Shohan, M.H.; Talukdar, M.M.N.; Biswas, J.C.; Ahammad, A.K.S. Assessment of Embryonic and Larval Development of Nile Tilapia under the Traditional and Re-Circulatory Thermostatic System in Relation to Climatic and Water Quality Variations. Aquac. J. 2023, 3, 70–89. [Google Scholar] [CrossRef]
- Mahalder, B.; Haque, M.M.; Siddique, M.A.B.; Hasan, N.A.; Alam, M.M.; Talukdar, M.M.N.; Shohan, M.H.; Ahasan, N.; Hasan, M.M.; Ahammad, A.K.S. Embryonic and Larval Development of Stinging Catfish, Heteropneustes fossilis, in Relation to Climatic and Water Quality Parameters. Life 2023, 13, 583. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | GPS Coordinates | ||
|---|---|---|---|
| Site 1 | Site 2 | Site 3 | |
| Gaglajur Haor, Netrokona | 24.803643, 91.107437 | 24.804359, 91.107201 | 24.802772, 91.107636 |
| Tetulia River, Barisal | 22.132731, 90.490980 | 22.148945, 90.501136 | 22.160998, 90.503075 |
| Kali River, Kishoregonj | 24.129192, 90.931914 | 24.130202, 90.931757 | 24.130536, 90.931705 |
| Meghna River, Chandpur | 24.154072, 90.935592 | 23.314200, 90.614044 | 23.285916, 90.636677 |
| Padma River | 23.926921, 89.257314 | 23.920214, 89.309277 | 23.942873, 89.321573 |
| Bay of Bengal, Cox’s Bazar | 21.520980, 91.829054 | 21.496385, 91.855833 | 21.428648, 91.948873 |
| Parameters | Bay of Bengal | Gaglajur Haor | Kali River | Meghna River | Padma River | Tetulia River | F Value | p Value | Level of Sig. |
|---|---|---|---|---|---|---|---|---|---|
| Air temperature (°C) | 26.24 ± 3.01 | 24.87 ± 4.00 | 25.60 ± 4.16 | 25.62 ± 3.95 | 25.78 ± 4.34 | 25.65 ± 3.85 | 0.152 | 0.979 | NS |
| Relative humidity (%) | 79.31 ± 6.89 | 77.69 ± 12.19 | 76.43 ± 14.18 | 78.87 ± 11.44 | 75.89 ± 14.82 | 80.34 ± 9.21 | 0.258 | 0.934 | NS |
| Rainfall (mm) | 26.00 ± 30.88 | 15.29 ± 16.44 | 8.18 ± 7.55 | 9.98 ± 9.72 | 7.43 ± 6.14 | 14.24 ± 14.74 | 2.09 | 0.076 | NS |
| Wind speed (km/h) | 7.97 ± 2.16 | 6.24 ± 1.66 | 7.17 ± 1.78 | 3.08 ± 0.87 | 7.41 ± 1.79 | 8.13 ± 2.31 | 12.84 | 0.000 | ** |
| Water temperature (°C) | 27.46 ± 0.84 | 27.47 ± 0.83 | 28.15 ± 1.32 | 27.24 ± 0.58 | 27.60 ± 0.74 | 27.61 ± 1.11 | 1.29 | 0.278 | NS |
| DO (ppm) | 6.43 ± 0.10 | 8.43 ± 0.15 | 7.56 ± 0.12 | 11.57 ± 0.43 | 12.58 ± 0.30 | 10.56 ± 0.14 | 1211.94 | 0.000 | ** |
| pH | 6.62 ± 0.09 | 9.34 ± 0.17 | 9.41 ± 0.12 | 8.21 ± 0.20 | 8.43 ± 0.10 | 7.59 ± 0.18 | 615.07 | 0.000 | ** |
| Salinity (ppm) | 29.67 ± 0.48 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.00 ± 0.00 | 0.43 ± 0.17 | 40,536.02 | 0.000 | ** |
| Class | |
|---|---|
| Phytoplankton | Genus/Species |
| Bacillariophyceae (24 spp.) | Amphora sp., Anomoeoneis sp., Asterionella sp., Bacillaria sp., Coscinodiscus sp., Cyclotella sp., Cymbella sp., Diatoma sp., Diatomella sp., Ditylum sp., Eunotia sp., Eucampia sp., Fragillaria sp., Gomphonema sp., Gyrosigma sp., Navicula sp., Nitzschia sp., Pleurosigma sp., Rhizosolenia sp., Surirella sp., Synedra sp., Skeletonema sp., Tabellaria sp., Thalassionema sp. |
| Chlorophyceae (26 spp.) | Actinastrus sp., Ankistrodesmus sp., Botryococcus sp., Chlorella sp., Closterium sp., Coelastrum sp., Micractinium sp., Microspora sp., Muogeotia sp., Oedogonium sp., Oocystis sp., Palmella sp., Pediastrum sp., Pleorococcus sp., Pteromonas sp., Scenedesmus sp., Selenestrum sp., Spirogyra sp., Staurastrum sp., Stichococcus sp., Tetraedron sp., Triceratium sp., Ulothrix sp., Uroglena sp., Volvox sp., Zygnema sp. |
| Cyanophyceae (15 spp.) | Anabaena sp., Aphanizomenon sp., Aphanocapsa sp., Chroococcus sp., Cosmarium sp., Closterium sp., Dictyophimus sp., Desmidium sp., Gomphosphaeria sp., Merismopedium sp., Microcystis sp., Microspora sp., Nostoc sp., Oscillatoria sp., Spirulina sp. |
| Euglenophyceae (2 spp.) | Euglena sp., Phacus sp. |
| Xanthophyceae (2 spp.) | Botrydium sp., Tribonema sp. |
| Dinophyceae (4 spp.) | Ceratium sp., Gyrodinium sp., Dinophysis sp., Peridinium sp |
| Zooplankton | |
| Copepoda (8 spp.) | Acartia sp., Calanus sp., Calanopia sp., Cyclops sp., Diaptomus sp., Laptodora sp., Naupleus sp., Paracartia sp. |
| Cladocera (5 spp.) | Bosmina sp., Diaphanosoma sp., Daphnia sp., Moina sp., Sida sp. |
| Rotifera (7 spp.) | Asplanchna sp., Brachionus sp., Filinia sp., Hexarthra sp., Keratilla sp., Poliarthra sp., Trichocerca sp. |
| Protozoa (2 spp.) | Difflugia sp., Favella sp. |
| Polychaeta (1 spp.) | Pedinosoma sp. |
| Gastropoda (1 spp.) | Notobranchaea sp. |
| Hydrozoa (3 spp.) | Aglaura sp., Chelophyes sp., Liriope sp. |
| Groups | Bay of Bengal | Gaglajur Haor | Kali River | Meghna River | Padma River | Tetulia River |
|---|---|---|---|---|---|---|
| Bacillariophyceae | 0.04 | 0.04 | 0.02 | 0.02 | 0.09 | 0.02 |
| Chlorophyceae | 0.05 | 0.01 | 0.02 | 0.06 | 0.01 | 0.03 |
| Cyanophyceae | −0.32 | −0.13 | 0.09 | −0.30 | −0.22 | 0.03 |
| Euglenophyceae | −0.06 | 0.01 | −0.39 | −0.36 | −0.29 | −0.50 |
| Dinophyceae | −0.59 | 0.20 | −0.24 | 0.24 | 0.02 | −0.27 |
| Xanthophyceae | −1.00 | −0.56 | −0.57 | −0.69 | −0.50 | −0.58 |
| Cladocera | −0.46 | −0.28 | −0.01 | −0.32 | −0.18 | −0.27 |
| Copepoda | −0.28 | −0.17 | −0.24 | −0.27 | −0.28 | −0.33 |
| Gastropoda | −0.24 | −0.37 | −0.41 | −0.46 | −0.67 | −0.69 |
| Hydrozoa | −0.80 | −0.60 | −0.59 | −0.76 | −0.63 | −0.65 |
| Rotifera | 0.25 | −0.34 | 0.05 | −0.11 | −0.04 | −0.52 |
| Polychaeta | −0.33 | −0.45 | −0.23 | −0.41 | −0.23 | −0.07 |
| Protozoan | −0.48 | −0.64 | −0.32 | −0.38 | −0.60 | −0.44 |
| Variable | PC1 | PC2 | PC3 |
|---|---|---|---|
| Plankton abundance in gut (Number/L) | −0.27 | 0.57 | −0.08 |
| Plankton abundance in water (Number/L) | −0.19 | 0.26 | −0.52 |
| Water temperature (°C) | 0.59 | −0.06 | −0.17 |
| DO (ppm) | −0.13 | −0.29 | −0.31 |
| pH | −0.02 | −0.43 | −0.26 |
| Salinity (ppm) | 0.09 | 0.58 | 0.4 |
| Relative humidity (%) | 0.25 | 0.18 | −0.43 |
| Rainfall (mm) | 0.36 | 0.27 | −0.15 |
| Wind speed (Km/h) | 0.38 | 0.14 | −0.02 |
| Air temperature (°C) | 0.54 | 0.03 | −0.21 |
| GSI | −0.38 | 0.27 | −0.34 |
| Eigenvalue | 3.24 | 2.76 | 1.78 |
| Variance (%) | 29.52 | 25.11 | 16.16 |
| Cumulative variance (%) | 29.52 | 54.64 | 70.79 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shohan, M.H.; Siddique, M.A.B.; Mahalder, B.; Haque, M.M.; Goswami, C.; Ahmed, M.B.U.; Alam, M.A.; Bashar, M.A.; Mahmud, Y.; Chowdhury, M.A.; et al. Multifaceted Linkages among Eco-Climatic Factors, Plankton Abundance, and Gonadal Maturation of Hilsa Shad, Tenualosa ilisha, Populations in Bangladesh. Climate 2024, 12, 40. https://doi.org/10.3390/cli12030040
Shohan MH, Siddique MAB, Mahalder B, Haque MM, Goswami C, Ahmed MBU, Alam MA, Bashar MA, Mahmud Y, Chowdhury MA, et al. Multifaceted Linkages among Eco-Climatic Factors, Plankton Abundance, and Gonadal Maturation of Hilsa Shad, Tenualosa ilisha, Populations in Bangladesh. Climate. 2024; 12(3):40. https://doi.org/10.3390/cli12030040
Chicago/Turabian StyleShohan, Mobin Hossain, Mohammad Abu Baker Siddique, Balaram Mahalder, Mohammad Mahfujul Haque, Chayon Goswami, Md. Borhan Uddin Ahmed, Mohammad Ashraful Alam, Md. Abul Bashar, Yahia Mahmud, Mahamudul Alam Chowdhury, and et al. 2024. "Multifaceted Linkages among Eco-Climatic Factors, Plankton Abundance, and Gonadal Maturation of Hilsa Shad, Tenualosa ilisha, Populations in Bangladesh" Climate 12, no. 3: 40. https://doi.org/10.3390/cli12030040