Increasing Air Temperatures and Its Effects on Growth and Productivity of Tomato in South Florida


Abstract
:1. Introduction
2. Results
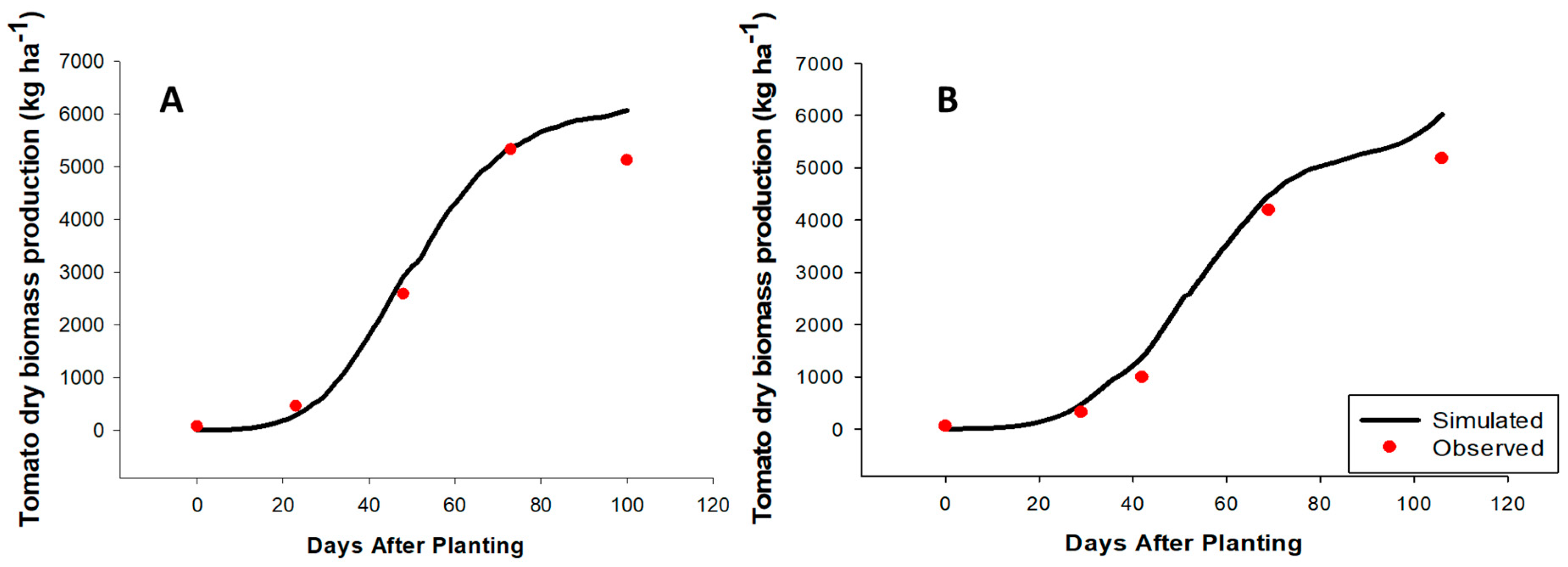
2.1. CROPGRO Tomato Model Calibration
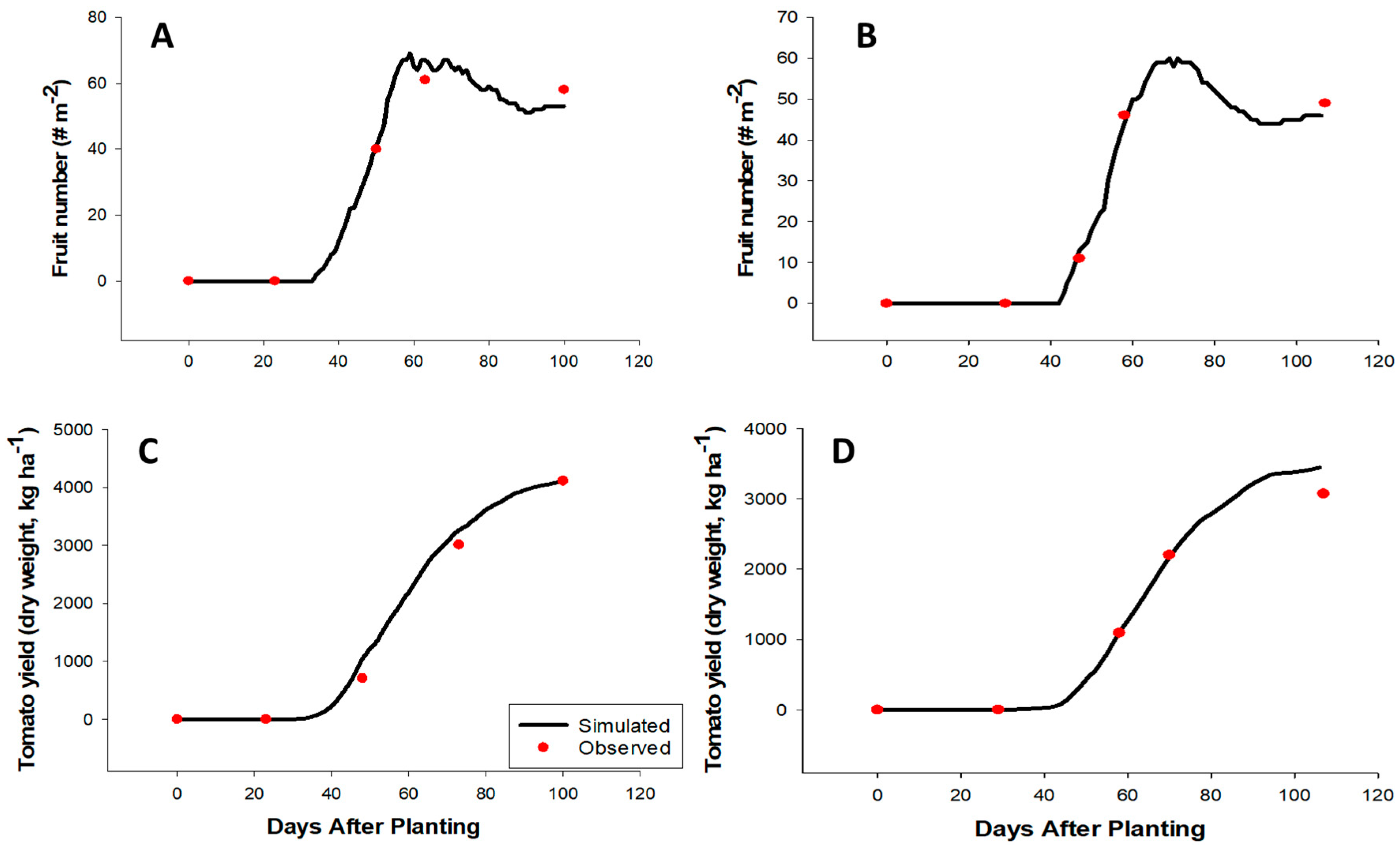
2.2. CROPGRO Tomato Model Evaluation
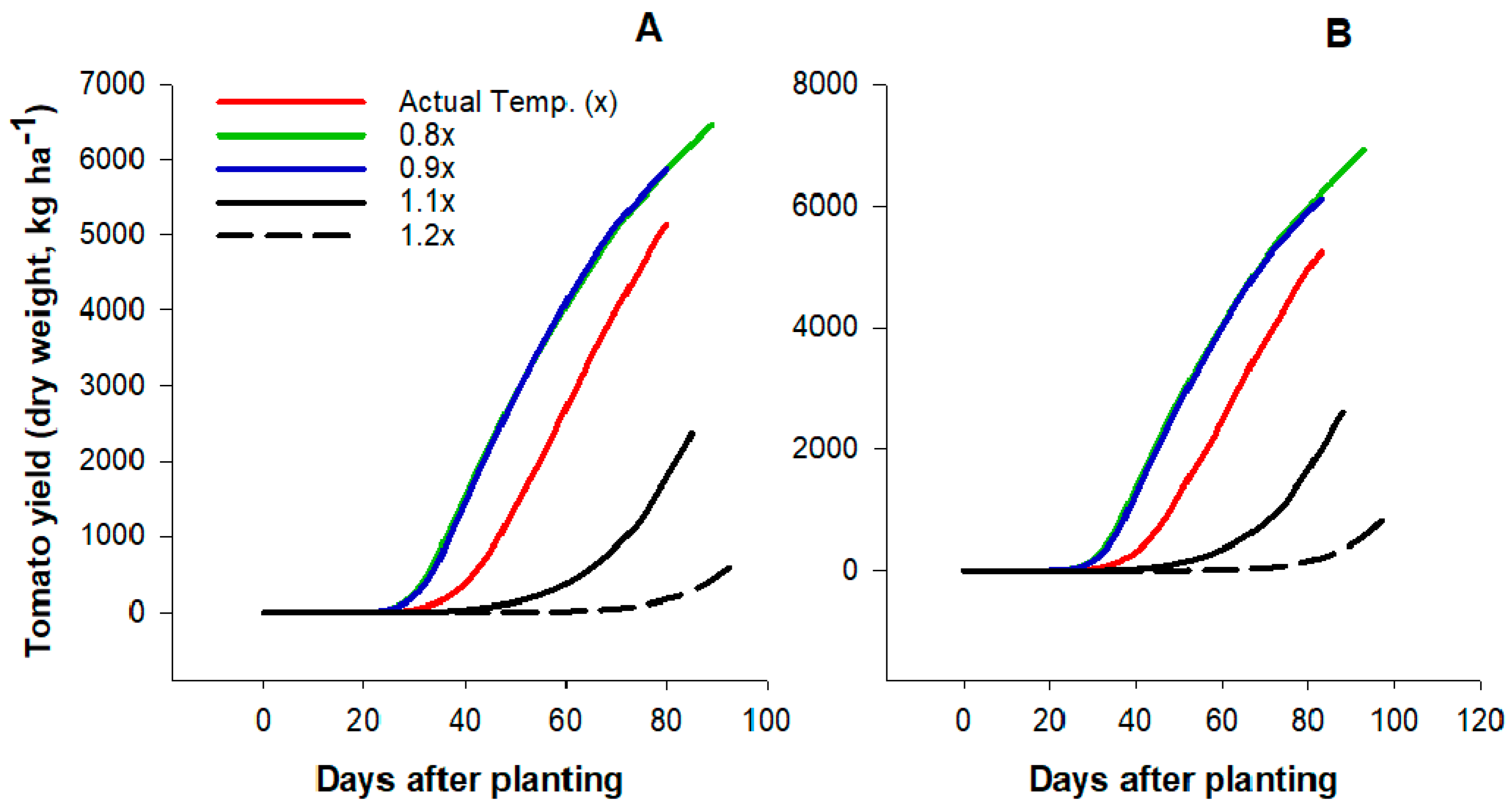
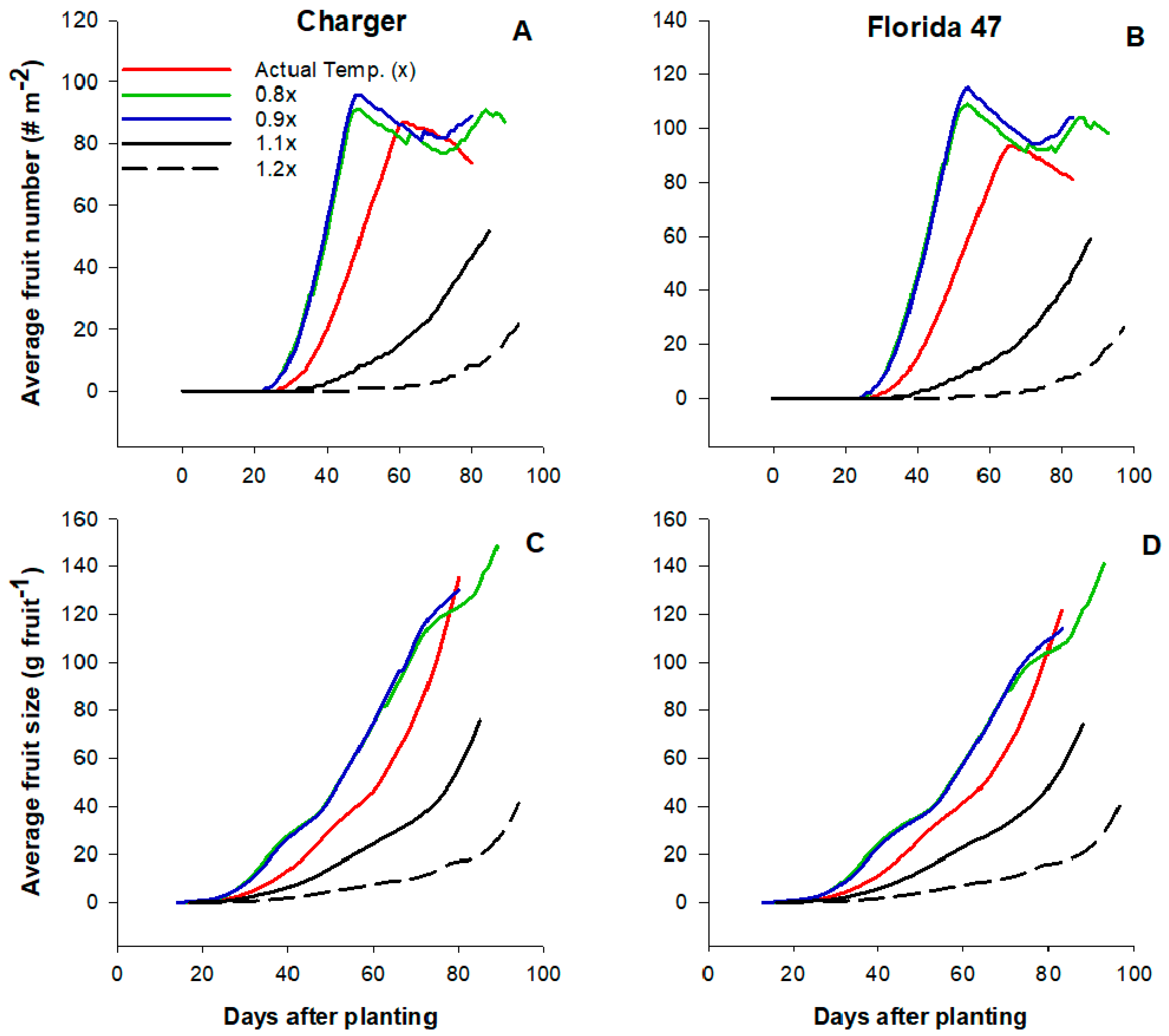
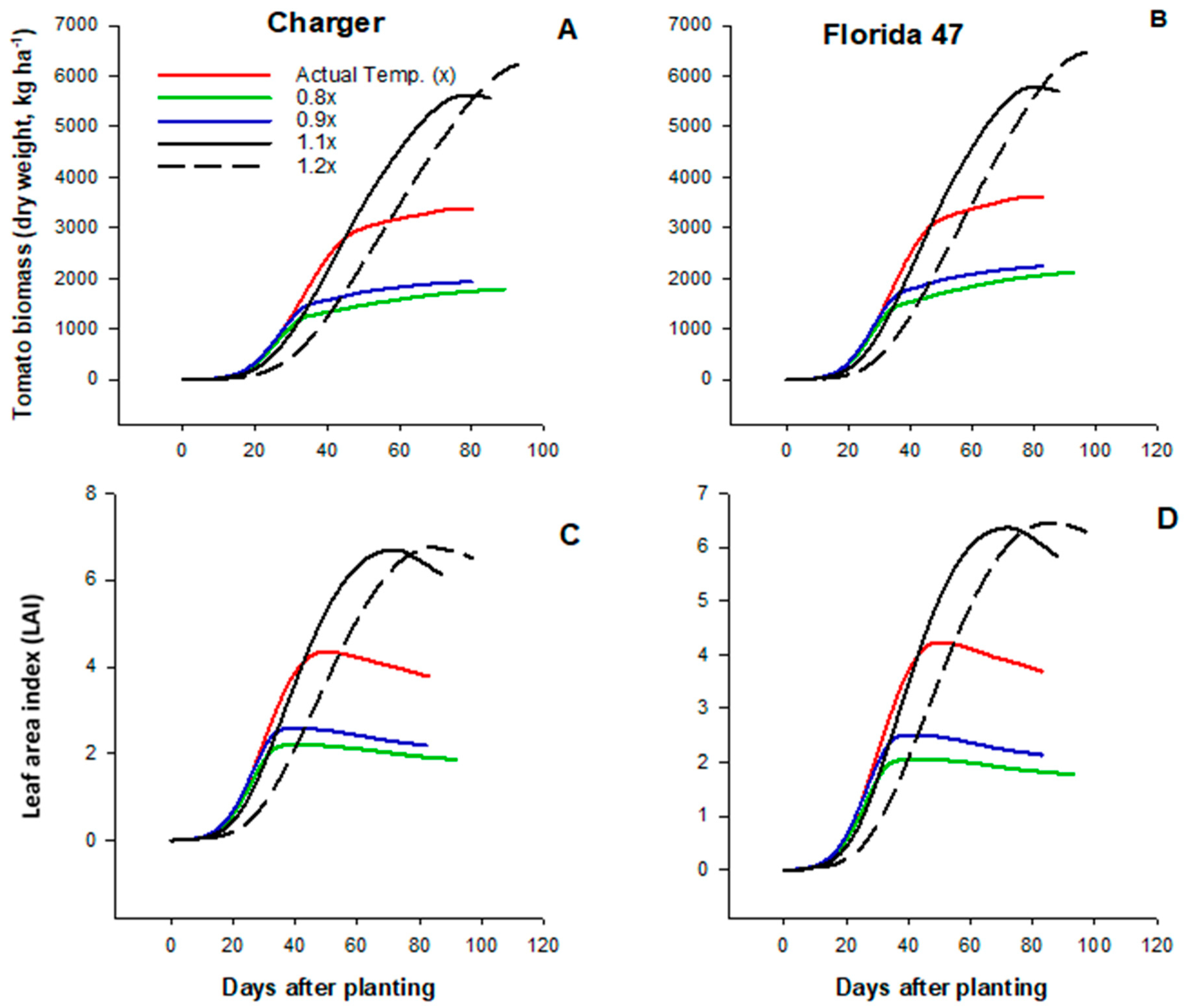
2.3. Effects of High Temperatures on Tomato Yield and Growth
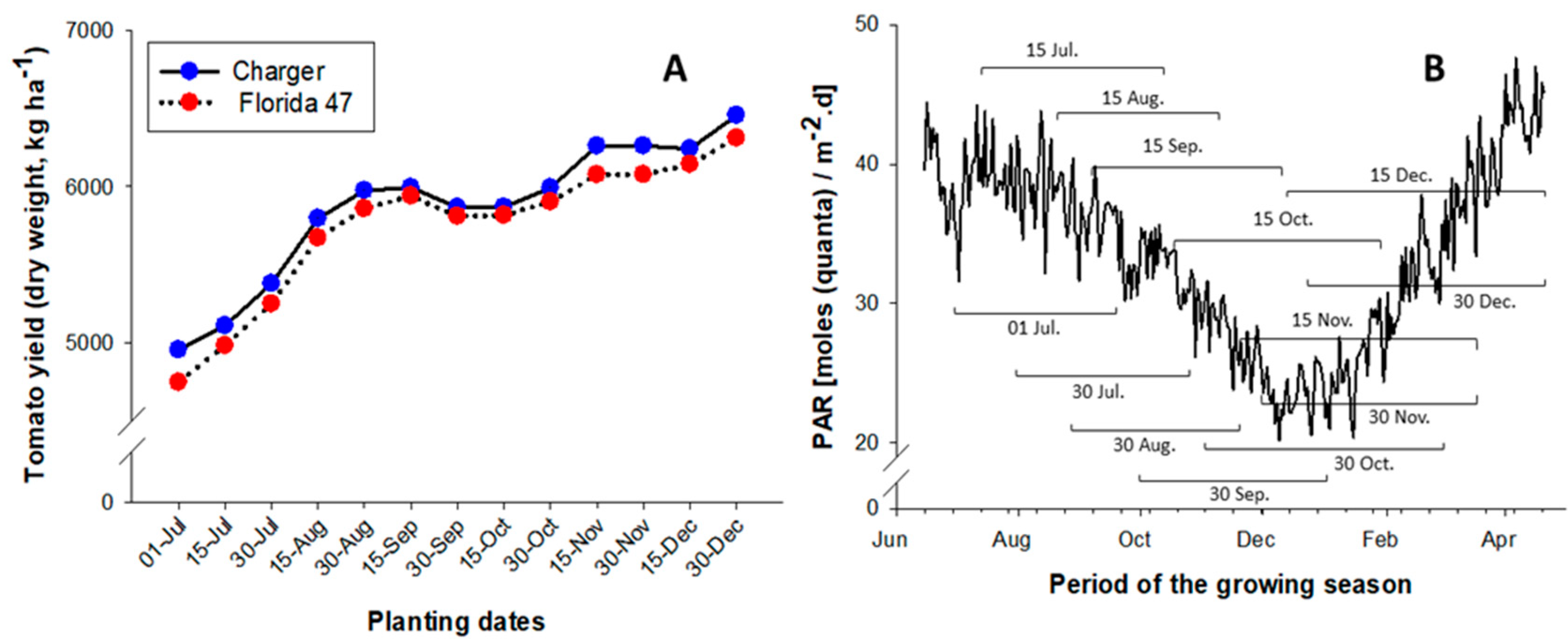
2.4. Effects of Planting Dates on Tomato Yield and Season Length
3. Discussions
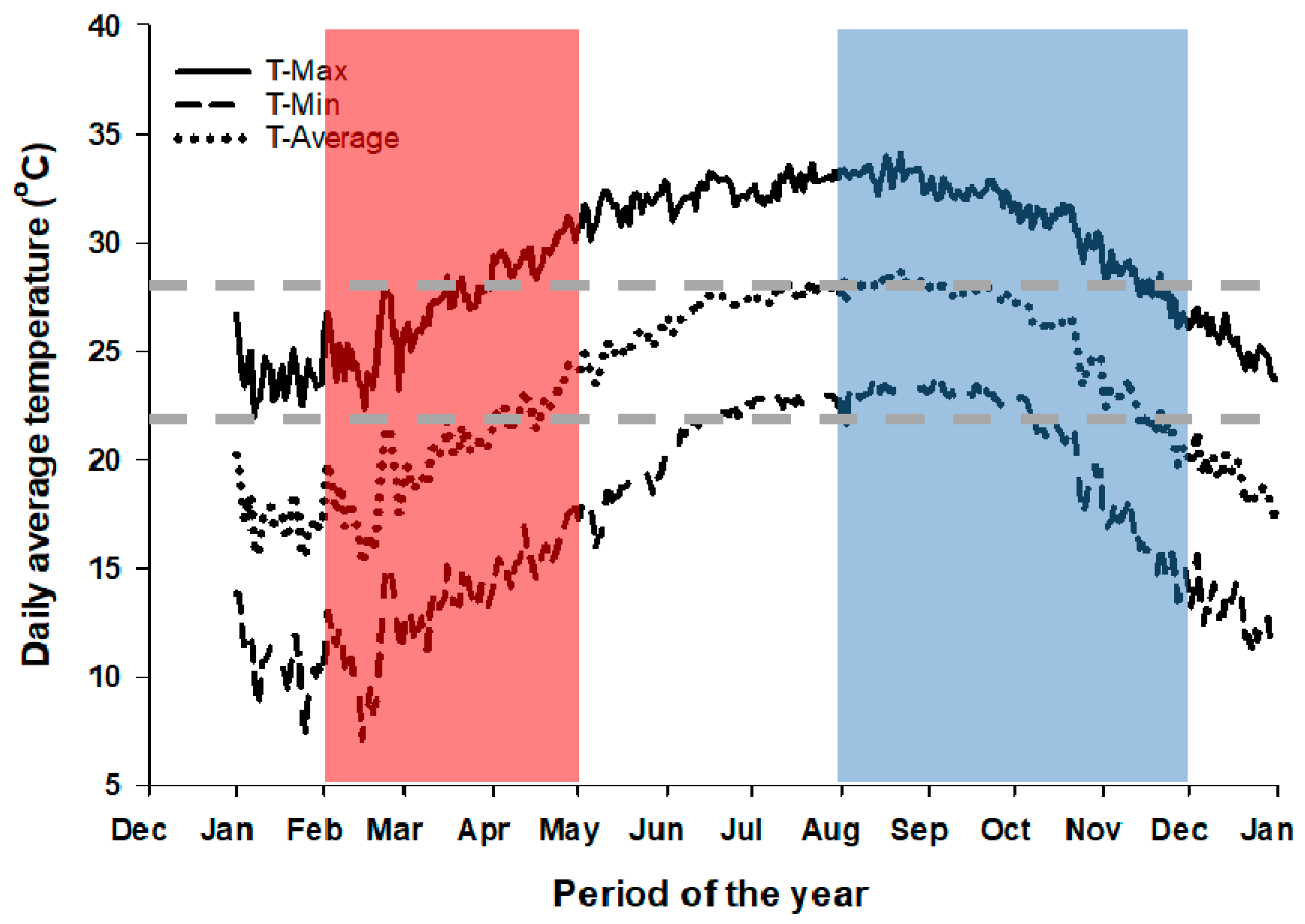
4. Materials and Methods
4.1. Study Location
4.2. DSSAT CROPGRO-Tomato Model
4.3. Climate, Soil, and Crop Data Collection
4.4. Model Calibration and Evaluation
4.5. Description of the Simulated Treatments
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- USDA/NASS. 2019 State Agriculture Overview for Florida. Available online: https://www.nass.usda.gov/Quick_Stats/Ag_Overview/stateOverview.php?state=FLORIDA (accessed on 22 June 2020).
- IPCC. Climate Change 2014: Synthesis Report; Plattner, G.-K., Ed.; IPCC: Copenhagen, Danmark, 2014. [Google Scholar]
- Singh, R.P.; Prasad, P.V.V.; Reddy, K.R. Impacts of Changing Climate and Climate Variability on Seed Production and Seed Industry. In Advances in Agronomy; Academic Press Inc.: Cambridge, MA, USA, 2013; Volume 118, pp. 49–110. [Google Scholar]
- Mittler, R.; Finka, A.; Goloubinoff, P. How do plants feel the heat? Trends Biochem. Sci. 2012, 37, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Bita, C.E.; Gerats, T. Plant tolerance to high temperature in a changing environment: Scientific fundamentals and production of heat stress-tolerant crops. Front. Plant Sci. 2013, 4, 273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazzeo, M.F.; Cacace, G.; Iovieno, P.; Massarelli, I.; Grillo, S.; Siciliano, R.A. Response mechanisms induced by exposure to high temperature in anthers from thermo-tolerant and thermo-sensitive tomato plants: A proteomic perspective. PLoS ONE 2018, 13, e201027. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mittler, R.; Blumwald, E. Genetic engineering for modern agriculture: Challenges and perspectives. Annu. Rev. Plant Biol. 2010, 61, 443–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozores-Hampton, F.; Kiran, M.; McAvoy, G. Blossom Drop, Reduced Fruit Set and Post-Pollination Disorders in Tomato. In Intitute of Food and Agricultural Sciences; IFAS Entension: Gainesville, FL, USA, 2012. [Google Scholar]
- Alam, M.; Sultana, N.; Ahmad, S.; Hossain, M.; Islam, A. Performance of heat tolerant tomato hybrid lines under hot, humid conditions. Bangladesh J. Agric. Res. 2010, 35, 367–373. [Google Scholar] [CrossRef]
- Berry, S.; Uddin, M. Effect of high temperature on fruit set in tomato cultivars and selected germplasm. HortScience 1988, 23, 606–608. [Google Scholar]
- Geisenberg, C.; Stewart, K. Field crop management. In The Tomato Crop; Atherton, J.G., Rudich, J., Eds.; Springer: Heidelberg, The Netherlands, 1986; pp. 511–557. [Google Scholar]
- Boote, K.J.; Rybak, M.R.; Scholberg, J.M.; Jones, J.W. Improving the CROPGRO-tomato model for predicting growth and yield response to temperature. HortScience 2012, 47, 1038–1049. [Google Scholar] [CrossRef]
- De Koning, A.N.M. Development and Dry Matter Distribution in Glasshouse Tomato: A Quantitative Approach; Wageningen Agricultural University: Wageningen, The Netherlands, 1994. [Google Scholar]
- Peet, M.M.; Willits, D.; Gardner, R. Response of ovule development and post-pollen production processes in male-sterile tomatoes to chronic, sub-acute high temperature stress. J. Exp. Bot. 1997, 48, 101–111. [Google Scholar] [CrossRef]
- Boote, K.J.; Pickering, N.B.; Allen, L.H.; Kirkham, M.B.; Olszyk, D.M.; Whitman, C.E. Plant Modeling: Advances and Gaps in Our Capability to Predict Future Crop Growth and Yield in Response to global Climate Change. In Advances in Carbon Dioxide Effects Research; Allen, L.H., Jr., Kirkham, M.B., Olszyk, D.M., Whitman, C.E., Eds.; John Wiley & Sons: Madison, WI, USA, 1997; Volume 61, pp. 179–228. [Google Scholar]
- Olson, S.M.; Simonne, E.H. Tomato Production Botany and Planting. In Vegetable Production Handbook of Florida, 23rd ed.; Dittmar, P., Freeman, J., Paret, M., Smith, H., Eds.; IFAS Entension: Gainesville, FL, USA, 2019; pp. 348–392. [Google Scholar]
- McAvoy, G.; University of Florida, Gainesville, FL, USA. Personal communication, 2019.
- Young, L.W.; Wilen, R.W.; Bonham-Smith, P.C. High temperature stress of Brassica napus during fowering reduces micro-and megagametophyte fertility, induces fruit abortion, and disrupts seed production. J. Exp. Bot. 2004, 55, 485–495. [Google Scholar] [CrossRef] [Green Version]
- Prasad, P.; Craufurd, P.Q.; Summerfield, R. Sensitivity of peanut to timing of heat stress during reproductive development. Crop Sci. 1999, 39, 1352–1357. [Google Scholar] [CrossRef]
- Sato, S.; Peet, M.M.; Thomas, J.F. Physiological factors limit fruit set of tomato (Lycopersicon esculentum Mill.) under chronic, mild heat stress. Plant Cell Environ. 2000, 23, 719–726. [Google Scholar] [CrossRef]
- USDA. Web Soil Survey. 2015. Available online: https://websoilsurvey.sc.egov.usda.gov/App/WebSoilSurvey.aspx (accessed on 22 June 2020).
- Kadyampakeni, D.M.; Morgan, K.T.; Schumann, A.W.; Nkedi-Kizza, P.; Obreza, T.A. Water use in drip-and microsprinkler-irrigated citrus trees soil & water management & conservation. Soil Sci. Soc. Am. J. 2014, 78, 1351–1361. [Google Scholar]
- Boote, K.J.; Jones, J.W.; Hoogenboom, G. Simulation of Crop Growth: CROPGRO Model; Marcel Dekker: New York, NY, USA, 1998. [Google Scholar]
- Jones, J.; Hoogenboom, G.; Porter, C.; Boote, K.J.; Batchelor, W.; Hunt, L.; Wilkens, P.; Singh, U.; Gijsman, A.; Ritchie, J. The DSSAT cropping system model. Eur. J. Agron. 2003, 18, 235–265. [Google Scholar] [CrossRef]
- Jones, J.W.; Tsuji, G.Y.; Hoogenboom, G.; Hunt, L.A.; Thornton, P.K.; Wilkens, P.W.; Imamura, D.T.; Bowen, W.T.; Singh, U. Decision Support System for Agrotechnology Transfer (DSSAT); Unversity of Fllorida: Gainesville, FL, USA, 2012. [Google Scholar]
- Adhikari, P.; Ale, S.; Bordovsky, J.P.; Thorp, K.R.; Modala, N.R.; Rajan, N.; Barnes, E.M. Simulating future climate change impacts on seed cotton yield in the Texas High Plains using the CSM-CROPGRO-Cotton model. Agric. Water Manag. 2016, 164, 317–330. [Google Scholar] [CrossRef]
- Modala, N.R.; Ale, S.; Rajan, N.; Munster, C.L.; Delaune, P.B.; Thorp, K.R.; Nair, S.S.; Barnes, E.M. Evaluation of the CSM-CROPGRO-cotton model for the texas rolling plains region and simulation of deficit irrigation strategies for increasing water use efficiency. Trans. ASABE 2015, 58, 685–696. [Google Scholar]
- Nagender, T.; Reddy, D.; Sreenivas, G. Evaluation of CSM-CROPGRO-Cotton model under varied plant densities and nitrogen levels for simulating crop growth, development and seed cotton. Int. J. Pure App. Biosci. 2017, 5, 1738–1754. [Google Scholar] [CrossRef]
- Thorp, K.R.; Hunsaker, D.J.; Bronson, K.F.; Barnes, E.M. Cotton irrigation scheduling using a crop growth model and FAO-56 methods: Field and simulation studies. Trans. ASABE 2017, 60, 2023–2039. [Google Scholar] [CrossRef] [Green Version]
- Amouzou, K.A.; Naab, J.B.; Lamers, J.P.; Borgemeister, C.; Becker, M.; Vlek, P.L. CROPGRO-Cotton model for determining climate change impacts on yield, water- and N- use efficiencies of cotton in the Dry Savanna of West Africa. Agric. Syst. 2018, 165, 85–96. [Google Scholar] [CrossRef]
- Rahman, M.H.U.; Ahmad, A.; Wajid, A.; Hussain, M.; Rasul, F.; Ishaque, W.; Islam, A.; Shelia, V.; Awais, M.; Ullah, A.; et al. Application of CSM-CROPGRO-Cotton model for cultivars and optimum planting dates: Evaluation in changing semi-arid climate. F. Crop. Res. 2019, 238, 139–152. [Google Scholar] [CrossRef]
- Ayankojo, I.T.; Morgan, K.T.; Ozores-Hampton, M.; Migliaccio, K.W. Effects of real-time location-specific drip irrigation scheduling on water use, plant growth, nutrient accumulation, and yield of Florida fresh-market tomato. HortScience 2018, 53, 1372–1378. [Google Scholar] [CrossRef] [Green Version]
- Allen, R.G.; Pereira, L.S.; Raes, D. Crop evapotranspiration-Guidelines for computing crop water requirements-FAO Irrigation and drainage paper 56 Table of Contents. FAO Rome 1998, 300, D5109. [Google Scholar]
- Migliaccio, K.; Morgan, K.T.; Vellidis, G.; Zotarelli, L.; Fraisse, C.; Zurweller, B.A.; Andreis, J.H.; Crane, J.H.; Rowland, D.L. Smartphone apps for irrigation scheduling. Trans. ASABE 2016, 59, 291–301. [Google Scholar]
- Ayankojo, I.T.; Morgan, K.T.; Mahmoud, K. Evaluation of soil water and nitrogen distribution by site-specific irrigation scheduling method in tomato crop grown on sandy soil. Soil Sci. Soc. Am. J. 2019, 83, 761–771. [Google Scholar] [CrossRef] [Green Version]
- Miller, L.; Vellidis, G.; Coolong, T. Comparing a smartphone irrigation scheduling application with water balance and soil moisture-based irrigation methods: Part I—plasticulture-grown tomato. Horttechnology 2018, 28, 354–361. [Google Scholar] [CrossRef] [Green Version]
- Miller, L.; Vellidis, G.; Mohawesh, O.; Coolong, T. Comparing a smartphone irrigation scheduling application with water balance and soil moisture-based irrigation methods: Part II—plasticulture-grown watermelon. Horttechnology 2018, 28, 362–369. [Google Scholar] [CrossRef] [Green Version]
- Vellidis, G.; Liakos, V.; Perry, C.; Tucker, M.; Collins, G.; Snider, J. A smartphone app for scheduling irrigation on cotton. In Proceedings of the 2014 Beltwide Cotton Conference, New Orleans, LA, USA, 6–8 January 2014. [Google Scholar]
- Liu, G.D.; Simonne, E.H.; Morgan, K.T.; Hochmuth, G.J.; Agehara, S.; Mylavarapu, R. Fertilizer Management for Vegetable Production in Florida. In Florida Vegetable Production Handbook, 23rd ed.; Dittmar, P., Freeman, J., Paret, M., Smith, H., Eds.; IFAS Entension: Gainesville, FL, USA, 2019; pp. 3–9. [Google Scholar]
- Jones, J.W.; He, J.; Boote, K.J.; Wilkens, P.; Porter, C.; Hu, Z.; Ahuja, L.R.; Ma, L. Estimating DSSAT cropping system cultivar-specific parameters using bayesian techniques. In Methods of Introducing System Models into Agricultural Research; John Wiley & Sons: Madison, WI, USA, 2011. [Google Scholar]
- Willmott, C.J.; Ackleson, S.G.; Davis, R.E.; Feddema, J.J.; Klink, K.; LeGates, D.R.; O’Donnell, J.; Rowe, C.M. Statistics for the evaluation and comparison of models. J. Geophys. Res. 1985, 90, 8995. [Google Scholar] [CrossRef] [Green Version]
- Loague, K.; Green, R.E. Statistical and graphical methods for evaluating solute transport models: Overview and application. J. Contam. Hydrol. 1991, 7, 51–73. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Description | Charger | FL.47 |
|---|---|---|---|
| EM.FL | Time between plant emergence and flower appearance (R1) (photothermal days) | 24.60 | 24.4 |
| FL.SH | Time between first flower and first pod (R3) (photothermal days) | 2.50 | 2.20 |
| FL.SD | Time between first flower and first seed (R5) (photothermal days) | 16.00 | 19.00 |
| SD.PM | Time between first seed (R5) and physiological maturity (R7) (photothermal days) | 45.01 | 45.20 |
| FL.LF | Time between first flower (R1) and end of leaf expansion (photothermal days) | 52.00 | 52.00 |
| LFMAX | Maximum leaf photosynthesis rate at 30 °C, 350 vpm CO2, and high light (mg CO2/m2-s) | 1.36 | 1.36 |
| SLAVR | Specific leaf area of cultivar under standard growth conditions (cm2/g) | 300.0 | 300.0 |
| SIZLF | Maximum size of full leaf (three leaflets) (cm2) | 300.0 | 300.0 |
| XFRT | Maximum fraction of daily growth that is partitioned to seed + shell | 0.80 | 0.78 |
| WTPSD | Maximum weight per seed (g) | 0.004 | 0.004 |
| SFDUR | Seed filling duration for pod cohort at standard growth conditions (photothermal days) | 26.00 | 26.00 |
| SDPDV | Average seed per pod under standard growing conditions (#/pod) | 300.0 | 300.0 |
| PODUR | Time required for cultivar to reach final pod load under optimal conditions (photothermal days) | 55.00 | 55.00 |
| THRSH | Threshing percentage. The maximum ratio of (seed/(seed+shell)) at maturity. Causes seed to stop growing as their dry weight increases until the shells are filled in a cohort. | 8.50 | 8.50 |
| SDPRO | Fraction protein in seeds (g(protein)/g(seed)) | 0.30 | 0.30 |
| SDLIP | Fraction oil in seeds (g(oil)/g(seed)) | 0.05 | 0.05 |
| Parameter | Simulated | Observed | RMSE | d-Stat |
|---|---|---|---|---|
| Days to anthesis | 24 | 25 | 0.71 | 0.98 |
| Days to physiological maturity | 103 | 102 | 0.71 | 0.99 |
| Dry biomass production (kg ha−1) | 6635 | 6419 | 217 | 0.97 |
| Fruit yield (kg ha−1) | 4310 | 4231 | 124 | 0.99 |
| Treatment | Simulated | Measured | RMSE | d-Stat |
|---|---|---|---|---|
| Anthesis date | ||||
| Fall 2016 | 22 | 23 | 0.71 | 0.98 |
| Spring 2017 | 27 | 27 | ||
| Physiological maturity date | ||||
| Fall 2016 | 100 | 100 | 1.17 | 0.97 |
| Spring 2017 | 106 | 105 | ||
| Season | Aboveground Biomass | Total Fruit Weight | Fruit Number | |||
|---|---|---|---|---|---|---|
| RMSE | d-Stat | RMSE | d-Stat | RMSE | d-Stat | |
| Fall 2016 | 456 | 0.99 | 248 | 0.99 | 15.12 | 0.78 |
| Spring 2017 | 440 | 0.96 | 112 | 0.98 | 2.00 | 0.99 |
| Planting Dates | Change in Yield (%) Z | Change in Season Length (days) y | Change in Yield (%) | Change in Season Length (days) |
|---|---|---|---|---|
| Charger | Florida 47 | |||
| 01-July | −23.3 | 0 | −24.8 | 0 |
| 15-July | −20.9 | 0 | −21.1 | 0 |
| 30-July | −16.7 | 0 | −16.8 | 0 |
| 15-August | −10.2 | +2 | −10.1 | +2 |
| 30-August | −7.5 | +5 | −7.2 | +5 |
| 15-September | −7.2 | +10 | −5.9 | +10 |
| 30-September | −9.1 | +16 | −7.9 | +16 |
| 15-October | −9.1 | +25 | −7.8 | +25 |
| 30-October | −7.2 | +32 | −6.5 | +31 |
| 15-November | −3.0 | +34 | −3.7 | +33 |
| 30-November | −3.0 | +34 | −3.7 | +33 |
| 15-December | −3.3 | +32 | −2.7 | +32 |
| 30-December | 0 | +29 | 0 | +28 |
| Stage | Description |
|---|---|
| 1 | From transplant until nine or more leaves on the main shoot unfolded |
| 2 | From first flower open until the eighth inflorescence first flower opened |
| 3 | From the ninth inflorescence first flower opening until its fruits reach typical size but no color changes |
| 4 | Fruit maturity until first harvest |
| 5 | After first harvest until last harvest |
| Treatment # | Treatment Description 1 | Avg. Daily Max. Temp. | Avg. Daily Min. Temp. |
|---|---|---|---|
| °C | |||
| T1 | Actual Temperature (AT) | 32 | 23 |
| T2 | AT*0.8 (0.8x) | 26 | 18 |
| T3 | AT*0.9 (0.9x) | 29 | 20 |
| T4 | AT*1.1 (1.1x) | 34 | 25 |
| T5 | AT*1.2 (1.2x) | 38 | 28 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayankojo, I.T.; Morgan, K.T. Increasing Air Temperatures and Its Effects on Growth and Productivity of Tomato in South Florida. Plants 2020, 9, 1245. https://doi.org/10.3390/plants9091245
Ayankojo IT, Morgan KT. Increasing Air Temperatures and Its Effects on Growth and Productivity of Tomato in South Florida. Plants. 2020; 9(9):1245. https://doi.org/10.3390/plants9091245
Chicago/Turabian StyleAyankojo, Ibukun T., and Kelly T. Morgan. 2020. "Increasing Air Temperatures and Its Effects on Growth and Productivity of Tomato in South Florida" Plants 9, no. 9: 1245. https://doi.org/10.3390/plants9091245