Development of High-Density Genetic Linkage Maps and Identification of Loci for Chestnut Gall Wasp Resistance in Castanea spp.

 ,
,  ,
, 
Abstract
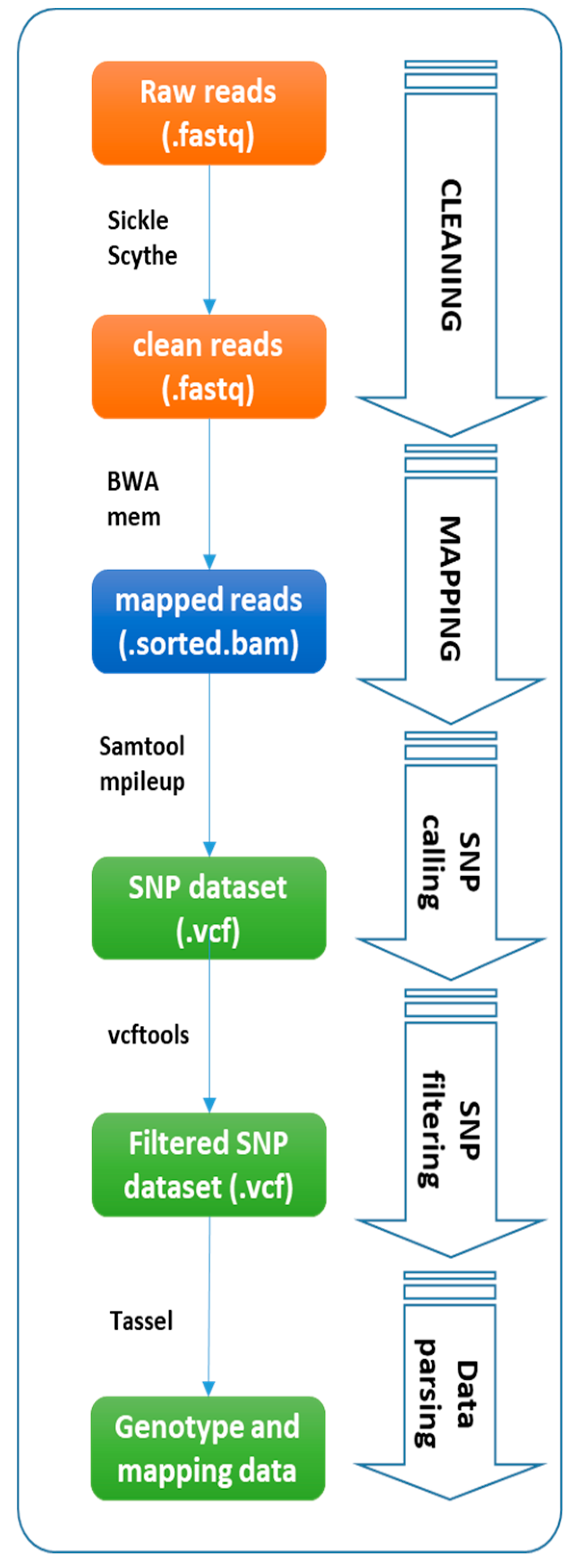
:1. Introduction
2. Results
2.1. ddRAD and SSR Genotyping
2.2. Genetic Linkage Maps
2.3. Phenotypic Distribution of Dryocosmus Kuriphilus Susceptibility
2.4. QTL Determining Resistance to Dryocosmus Kuriphilus (Rdk1 QTL Region)
2.5. Identification of Candidate Genes Within the Rdk1 QTL Region
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Simple Sequence Repeats Genotyping
4.3. ddRAD-Seq Library Construction and Sequencing
4.4. SNP Detection
4.5. Genetic Linkage Map Construction
4.6. Phenotypic Evaluation of Dryocosmus Kuriphilus Susceptibility
4.7. Quantitative Trait Loci (QTL) Detection
4.8. Expression Analysis of Genes Within the Rdk1 QTL Region
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bounous, G.; Marinoni, D.T. Chestnut: Botany, Horticulture, and Utilization. Hortic. Rev. 2010, 31, 291–347. [Google Scholar] [CrossRef]
- Pereira-Lorenzo, S.; Ballester, A.; Corredoira, E.; Viéitez, A.M.; Agnanostakis, S.; Costa, R.L.; Bounous, G.; Botta, R.; Beccaro, G.L.; Kubisiak, T.L.; et al. Chestnut. In Fruit Breeding; Badenes, M.L., Byrne, D.H., Eds.; Springer: New York, NY, USA, 2012; pp. 729–769. [Google Scholar]
- Torello-Marinoni, D.; Nishio, S.; Portis, E.; Valentini, N.; Sartor, C.; Dini, F.; Ruffa, P.; Oglietti, S.; Martino, G.; Akkak, A.; et al. Development of a genetic linkage map for molecular breeding of chestnut. Acta Hortic. 2018, 1220, 23–28. [Google Scholar] [CrossRef]
- Santos, C.; Nelson, C.D.; Zhebentyayeva, T.; Machado, H.; Gomes-Laranjo, J.; Costa, R.L. First interspecific genetic linkage map for Castanea sativa × Castanea crenata revealed QTLs for resistance to Phytophthora cinnamomi. PLoS ONE 2017, 12, e0184381. [Google Scholar] [CrossRef] [Green Version]
- Santos, C.; Zhebentyayeva, T.; Serrazina, S.; Nelson, C.D.; Costa, R.L. Development and characterization of EST-SSR markers for mapping reaction to Phytophthora cinnamomi in Castanea spp. Sci. Hortic. 2015, 194, 181–187. [Google Scholar] [CrossRef]
- Hebard, F.V. The backcross breeding program of the American Chestnut Foundation. J. Am. Chestnut Found 2006, 19, 55–77. [Google Scholar]
- Tanaka, K.; Kotobuki, K. Comparative Ease of Pellicle Removal among Japanese Chestnut (Castanea crenata Sieb. et Zucc.) and Chinese Chestnut (C. mollissima Blume) and Their Hybrids. J. Jpn. Soc. Hortic. Sci. 1992, 60, 811–819. [Google Scholar] [CrossRef] [Green Version]
- Shimura, I. Chestnut breeding history. Agric. Hortic. 2003, 58, 30–32. (In Japanese) [Google Scholar]
- Moriya, S.; Inoue, K.; Otake, A.; Shiga, M.; Mabuchi, M. Decline of the chestnut gallwasp population, Dryocosmus kuriphilus Yasumatsu (Hymenoptera: Cynipidae) after the establishment of Torymus sinensis Kajimo (Hymenoptera: Torymidae). Appl. Entomol. Zool. 1989, 24, 231–233. [Google Scholar] [CrossRef] [Green Version]
- Aebi, A.; Schönrogge, K.; Melika, G.; Alma, A.; Bosio, G.; Quacchia, A.; Picciau, L.; Abe, Y.; Moriya, S.; Yara, K.; et al. Parasitoid Recruitment to the Globally Invasive Chestnut Gall Wasp Dryocosmus kuriphilus. In Galling Arthropods and Their Associates; Ozaki, K., Yukwa, J., Ohgushi, T., Price, P.W., Eds.; Springer: Tokyo, Japan, 2007; pp. 103–121. [Google Scholar]
- Sartor, C.; Dini, F.; Marinoni, D.T.; Mellano, M.G.; Beccaro, G.L.; Alma, A.; Quacchia, A.; Botta, R. Impact of the Asian wasp Dryocosmus kuriphilus (Yasumatsu) on cultivated chestnut: Yield loss and cultivar susceptibility. Sci. Hortic. 2015, 197, 454–460. [Google Scholar] [CrossRef]
- Dini, F.; Sartor, C.; Botta, R. Detection of a hypersensitive reaction in the chestnut hybrid ‘Bouche de Bétizac’ infested by Dryocosmus kuriphilus Yasumatsu. Plant Physiol. Biochem. 2012, 60, 67–73. [Google Scholar] [CrossRef]
- Kubisiak, T.L.; Hebard, F.V.; Nelson, C.D.; Zhang, J.; Bernatzky, R.; Huang, H.; Anagnostakis, S.L.; Doudrick, R.L. Molecular Mapping of Resistance to Blight in an Interspecific Cross in the genus Castanea. Phytopathology 1997, 87, 751–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barreneche, T.; Casasoli, M.; Russell, K.; Akkak, A.; Meddour, H.; Plomion, C.; Villani, F.; Kremer, A. Comparative mapping between Quercus and Castanea using simple-sequence repeats (SSRs). Theor. Appl. Genet. 2003, 108, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Casasoli, M.; Mattioni, C.; Cherubini, M.; Villani, F. A genetic linkage map of European chestnut (Castanea sativa Mill.) based on RAPD, ISSR and isozyme markers. Theor. Appl. Genet. 2001, 102, 1190–1199. [Google Scholar] [CrossRef]
- Casasoli, M.; Derory, J.; Morera-Dutrey, C.; Brendel, O.; Porth, I.; Guehl, J.-M.; Villani, F.; Kremer, A. Comparison of Quantitative Trait Loci for Adaptive Traits Between Oak and Chestnut Based on an Expressed Sequence Tag Consensus Map. Genetics 2005, 172, 533–546. [Google Scholar] [CrossRef] [Green Version]
- HGW. Hardwood Genomic Project. Available online: https://hardwoodgenomics.org/Genome-assembly/1962958 (accessed on 20 November 2019).
- Kubisiak, T.L.; Nelson, C.D.; Staton, M.E.; Zhebentyayeva, T.; Smith, C.; Olukolu, B.A.; Fang, G.-C.; Hebard, F.V.; Anagnostakis, S.; Wheeler, N.; et al. A transcriptome-based genetic map of Chinese chestnut (Castanea mollissima) and identification of regions of segmental homology with peach (Prunus persica). Tree Genet. Genomes 2012, 9, 557–571. [Google Scholar] [CrossRef] [Green Version]
- Nishio, S.; Takada, N.; Yamamoto, T.; Terakami, S.; Hayashi, T.; Sawamura, Y.; Saito, T. Mapping and pedigree analysis of the gene that controls the easy peel pellicle trait in Japanese chestnut (Castanea crenata Sieb. et Zucc.). Tree Genet. Genomes 2013, 9, 723–730. [Google Scholar] [CrossRef]
- Nishio, S.; Terakami, S.; Matsumoto, T.; Yamamoto, T.; Takada, N.; Kato, H.; Katayose, Y.; Saito, T. Identification of QTLs for Agronomic Traits in the Japanese Chestnut (Castanea crenata Sieb. et Zucc.) Breeding. Hortic. J. 2018, 87, 43–54. [Google Scholar] [CrossRef] [Green Version]
- Casasoli, M.; Pot, D.; Plomion, C.; Monteverdi, M.C.; Barreneche, T.; Lauteri, M.; Villani, F. Identification of QTLs affecting adaptive traits in Castanea sativa Mill. Plant Cell Environ. 2004, 27, 1088–1101. [Google Scholar] [CrossRef]
- Elshire, R.J.; Glaubitz, J.C.; Sun, Q.; Poland, J.; Kawamoto, K.; Buckler, E.S.; Mitchell, S.E. A Robust, Simple Genotyping-by-Sequencing (GBS) Approach for High Diversity Species. PLoS ONE 2011, 6, e19379. [Google Scholar] [CrossRef] [Green Version]
- Peterson, B.K.; Weber, J.N.; Kay, E.H.; Fisher, H.S.; Hoekstra, H.E. Double Digest RADseq: An Inexpensive Method for De Novo SNP Discovery and Genotyping in Model and Non-Model Species. PLoS ONE 2012, 7, e37135. [Google Scholar] [CrossRef] [Green Version]
- Shirasawa, K.; Hirakawa, H.; Isobe, S. Analytical workflow of double-digest restriction site-associated DNA sequencing based on empirical andin silicooptimization in tomato. DNA Res. 2016, 23, 145–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Dijk, E.L.; Auger, H.; Jaszczyszyn, Y.; Thermes, C. Ten years of next-generation sequencing technology. Trends Genet. 2014, 30, 418–426. [Google Scholar] [CrossRef] [PubMed]
- Bai, B.; Wang, L.; Lee, M.; Zhang, Y.; Alfiko, Y.; Ye, B.Q.; Wan, Z.Y.; Lim, C.H.; Suwanto, A.; Chua, N.-H.; et al. Genome-wide identification of markers for selecting higher oil content in oil palm. BMC Plant Biol. 2017, 17, 93. [Google Scholar] [CrossRef] [PubMed]
- Nagano, S.; Shirasawa, K.; Hirakawa, H.; Maeda, F.; Ishikawa, M.; Isobe, S.N. Discrimination of candidate subgenome-specific loci by linkage map construction with an S1 population of octoploid strawberry (Fragaria × Ananassa). BMC Genom. 2017, 18, 374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yagi, M.; Shirasawa, K.; Waki, T.; Kume, T.; Isobe, S.; Tanase, K.; Yamaguchi, H. Construction of an SSR and RAD marker-based benetic linkage map for Carnation (Dianthus caryophyllus L.). Plant Mol. Biol. Rep. 2017, 35, 110–117. [Google Scholar] [CrossRef]
- Zhong, Y.-J.; Zhou, Y.-Y.; Li, J.-X.; Yu, T.; Wu, T.-Q.; Luo, J.-N.; Luo, S.-B.; Huang, H. A high-density linkage map and QTL mapping of fruit-related traits in pumpkin (Cucurbita moschata Duch.). Sci. Rep. 2017, 7, 12785. [Google Scholar] [CrossRef] [Green Version]
- UNIPROT. Available online: https://www.uniprot.org/uniprot/Q9LRR4 (accessed on 25 February 2020).
- Acquadro, A.; Marinoni, D.T.; Sartor, C.; Dini, F.; Macchio, M.; Botta, R. Transcriptome characterization and expression profiling in chestnut cultivars resistant or susceptible to the gall wasp Dryocosmus kuriphilus. Mol. Genet. Genom. 2019, 295, 107–120. [Google Scholar] [CrossRef]
- Bonierbale, M.W.; Plaisted, R.L.; Tanksley, S.D. RFLP Maps Based on a Common Set of Clones Reveal Modes of Chromosomal Evolution in Potato and Tomato. Genetics 1988, 120, 1095–1103. [Google Scholar]
- Causse, M.A.; Fulton, T.M.; Cho, Y.G.; Ahn, S.N.; Chunwongse, J.; Wu, K.; Xiao, J.; Yu, Z.; Ronald, P.C.; Harrington, S.E.; et al. Saturated Molecular Map of the Rice Genome Based on an Interspecific Backcross Population. Genetics 1994, 138, 1251–1274. [Google Scholar]
- Gebhardt, C.; Ritter, E.; Barone, A.; Debener, T.; Walkemeier, B.; Schachtschabel, U.; Kaufmann, H.; Thompson, R.D.; Bonierbale, M.W.; Ganal, M.W.; et al. FFLP maps of potato and their alignment with the homologous tomato genome. Theor. Appl. Genet. 1991, 83, 49–57. [Google Scholar] [CrossRef]
- Ky, C.L.; Barre, P.; Lorieux, M.; Trouslot, P.; Akaffou, S.; Louarn, J.; Charrier, A.; Hamon, S.; Noirot, M. Interspecific genetic linkage map, segregation distortion and genetic conversion in coffee (Coffea sp.). Theor. Appl. Genet. 2000, 101, 669–676. [Google Scholar] [CrossRef]
- Lawrence, E.J.; Griffin, C.H.; Henderson, I.R. Modification of meiotic recombination by natural variation in plants. J. Exp. Bot. 2017, 68, 5471–5483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coll, N.S.; Vercammen, D.; Smidler, A.; Clover, C.; Van Breusegem, F.; Dangl, J.L.; Epple, P. Arabidopsis Type I Metacaspases Control Cell Death. Science 2010, 330, 1393–1397. [Google Scholar] [CrossRef] [PubMed]
- Bittner-Eddy, P.D.; Beynon, J. The Arabidopsis Downy Mildew Resistance Gene, RPP13-Nd, Functions Independently of NDR1 and EDS1 and Does Not Require the Accumulation of Salicylic Acid. Mol. Plant Microbe Interact. 2001, 14, 416–421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferracini, C.; Gonella, E.; Ferrari, E.; Saladini, M.A.; Picciau, L.; Tota, F.; Pontini, M.; Alma, A. Novel insight in the life cycle of Torymus sinensis, biocontrol agent of the chestnut gall wasp. BioControl 2014, 60, 169–177. [Google Scholar] [CrossRef]
- Kotobuki, K.; Saito, T.; Kashimura, Y.; Shoda, M. Chestnut breeding program in national institute of fruit tree science, Japan. Acta Hortic. 1999, 494, 323–326. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure from small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Aldrich, P.R.; Michler, C.H.; Sun, W.; Romero-Severson, J. Microsatellite markers for northern red oak (Fagaceae: Quercus rubra). Mol. Ecol. Notes 2002, 2, 472–474. [Google Scholar] [CrossRef]
- Buck, E.; Hadonou, M.; James, C.J.; Blakesley, D.; Russell, K. Isolation and characterization of polymorphic microsatellites in European chestnut (Castanea sativa Mill.). Mol. Ecol. Notes 2003, 3, 239–241. [Google Scholar] [CrossRef]
- Kampfer, S.; Lexer, C.; Glössl, J.; Steinkellner, H. Characterization of (GA) n Microsatellite Loci from Quercus Robur. Hereditas 2004, 129, 183–186. [Google Scholar] [CrossRef]
- Marinoni, D.; Akkak, A.; Bounous, G.; Edwards, K.J.; Botta, R. Development and characterization of microsatellite markers in Castanea sativa (Mill.). Mol. Breed. 2003, 11, 127–136. [Google Scholar] [CrossRef]
- Nishio, S.; Yamamoto, T.; Terakami, S.; Sawamura, Y.; Takada, N.; Nishitani, C.; Saito, T. Novel genomic and EST-derived SSR markers in Japanese chestnuts. Sci. Hortic. 2011, 130, 838–846. [Google Scholar] [CrossRef]
- Steinkellner, H.; Fluch, S.; Turetschek, E.; Lexer, C.; Streiff, R.; Kremer, A.; Burg, K.; Glössl, J. Identification and characterization of (GA/CT) n-microsatellite loci from Quercus petraea. Plant Mol. Biol. 1997, 33, 1093–1096. [Google Scholar] [CrossRef]
- Acquadro, A.; Barchi, L.; Gramazio, P.; Portis, E.; Vilanova, S.; Comino, C.; Plazas, M.; Prohens, J.; Lanteri, S. Coding SNPs analysis highlights genetic relationships and evolution pattern in eggplant complexes. PLoS ONE 2017, 12, e0180774. [Google Scholar] [CrossRef]
- Li, H.; Durbin, R. Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 2009, 25, 1754–1760. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.R.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Van Ooijen, J. JoinMap® 4, Software for the Calculation of Genetic Linkage Maps in Experimental Populations; Kyazma BV: Wageningen, The Netherlands, 2006. [Google Scholar]
- Grattapaglia, D.; Sederoff, R. Genetic Linkage Maps of Eucalyptus Grandis and Eucalyptus Urophylla using a pseudo-testcross: Mapping Strategy and Rapd Markers. Genetics 1994, 137, 1121–1137. [Google Scholar]
- Kosambi, D.D. The estimation of map distances from recombination values. Ann. Eugen. 1943, 12, 172–175. [Google Scholar] [CrossRef]
- Voorrips, R.E. MapChart: Software for the graphical presentation of linkage maps and QTLs. J. Hered. 2002, 93, 77–78. [Google Scholar] [CrossRef] [Green Version]
- Lander, E.S.; Botstein, D. Mapping Mendelian Factors Underlying Quantitative Traits Using RFLP Linkage Maps. Genetics 1989, 121, 185–199. [Google Scholar]
- Jansen, R.C.; Stam, P. High resolution of quantitative traits into multiple loci via interval mapping. Genetics 1994, 136, 1447–1455. [Google Scholar]
- Van Ooijen, J.W.; Boer, M.P.; Jansen, R.C.; Maliepaard, C. MapQTL 4.0: Software for the Calculation of QTL Positions on Genetic Maps; Plant Research International: Wageningen, The Netherlands, 2002. [Google Scholar]
- Churchill, G.A.; Doerge, R.W. Empirical Threshold Values for Quantitative Trait Mapping. Genetics 1994, 138, 963–971. [Google Scholar]



{kind=link}
{kind=link}
| Linkage Group | Number of Loci | Length of Linkage Groups (cM) | Number of Markers | Average Loci Interval (cM/Loci) | Mapped Scaffolds | Total Length of Scaffolds (bp) |
|---|---|---|---|---|---|---|
| Bouche_A | 217 | 95.6 | 335 | 0.44 | 322 | 20,080,944 |
| Bouche_B | 123 | 59.9 | 236 | 0.49 | 227 | 12,693,145 |
| Bouche_C | 119 | 63.3 | 186 | 0.53 | 179 | 13,168,399 |
| Bouche_D | 110 | 66.2 | 169 | 0.60 | 157 | 11,638,867 |
| Bouche_E | 156 | 72.9 | 223 | 0.47 | 216 | 9,764,018 |
| Bouche_F | 163 | 73.8 | 238 | 0.45 | 229 | 14,708,920 |
| Bouche_G | 86 | 59.6 | 161 | 0.69 | 150 | 7,498,312 |
| Bouche_H | 47 | 58.8 | 82 | 1.25 | 71 | 4,308,112 |
| Bouche_I | 103 | 59.5 | 172 | 0.58 | 166 | 8,554,610 |
| Bouche_J | 119 | 59.0 | 201 | 0.50 | 189 | 10,514,064 |
| Bouche_K | 131 | 83.6 | 195 | 0.64 | 182 | 8,501,984 |
| Bouche_L | 85 | 57.4 | 123 | 0.68 | 114 | 5,773,207 |
| Bouche total | 1459 | 809.6 | 2321 | 0.55 | 2202 | 127,204,582 |
| Madonna_A | 171 | 91.7 | 257 | 0.54 | 249 | 15,914,736 |
| Madonna_B | 118 | 62.9 | 148 | 0.53 | 143 | 7,642,270 |
| Madonna_C | 97 | 57.5 | 126 | 0.59 | 121 | 9,479,624 |
| Madonna_D | 49 | 57.2 | 57 | 1.17 | 49 | 3,338,321 |
| Madonna_E | 105 | 60.7 | 142 | 0.58 | 138 | 6,422,502 |
| Madonna_F | 66 | 68.3 | 85 | 1.03 | 78 | 4,119,360 |
| Madonna_G | 72 | 56.5 | 89 | 0.78 | 80 | 3,607,147 |
| Madonna_H | 99 | 68.2 | 119 | 0.69 | 109 | 6,640,418 |
| Madonna_I | 107 | 56.5 | 132 | 0.53 | 126 | 6,420,027 |
| Madonna_J | 60 | 54.9 | 67 | 0.91 | 59 | 3,508,456 |
| Madonna_K | 90 | 64.1 | 110 | 0.71 | 101 | 4,308,112 |
| Madonna_L | 55 | 54.9 | 65 | 1.00 | 60 | 3,621,485 |
| Madonna total | 1089 | 753.3 | 1397 | 0.69 | 1313 | 75,022,458 |
| Scaffold | Scaffold Position (bp) | LG Position (cM) | First Season | Second Season | ||||
|---|---|---|---|---|---|---|---|---|
| GW | LOD | PVE | GW | LOD | PVE | |||
| scaffold03566 | 22494 | 45.41 | 3.2 | 3.55 | 67.2 | 3.3 | 3.62 | 69.4 |
| scaffold07739 | 7819 | 45.41 | ||||||
| scaffold10655 | 2698 | 45.41 | ||||||
| scaffold00261 | 32530 | 45.41 | ||||||
| scaffold07319 | 12557 | 45.60 | 3.40 | 3.55 | ||||
| scaffold18444 | 371 | 45.83 | 3.32 | 3.40 | ||||
| scaffold06031 | 22490 | 46.09 | 3.22 | 3.38 | ||||
| scaffold04450 | 24357 | 46.38 | 3.22 | 3.38 | ||||
| scaffold05894 | 16508 | 46.45 | 3.22 | 3.38 | ||||
| scaffold06906 | 12722 | 46.45 | ||||||
| scaffold10721 | 6288 | 46.45 | ||||||
| Scaffold (C. mollissima Genome) | Name of Gene (C. mollissima Genome) | A. thaliana Orthologues (C. mollissima Genome) | A. thaliana Orthologues (Bouche Transcriptome) | MI_vs_MNI (GFOLD) | BI_vs_BNI (GFOLD) | BI_vs_MI (GFOLD) | BNI_vs_MNI (GFOLD) |
|---|---|---|---|---|---|---|---|
| scaffold00261 | maker-scaffold00261-augustus-gene-0.29-mRNA-1 | AT2G44900; AT3G60350.1 | AT2G44900.1 | 0.25 | 0.53 | 0.87 | 0.59 |
| scaffold00261 | maker-scaffold00261-snap-gene-0.32-mRNA-1 | AT2G44910.1; AT4G37790; AT2G22800.1; AT3G60390.1 | AT3G60390.1 | 1.35 | 0.30 | 0.00 | 0.87 |
| scaffold00261 | maker-scaffold00261-snap-gene-0.33-mRNA-1 | AT3G60370.1 | AT3G60370.1 | 0.14 | 0.06 | −0.20 | −0.25 |
| scaffold00261 | maker-scaffold00261-augustus-gene-0.30-mRNA-1 | AT5G65274.1; AT4G01710.1 | - | 0.71 | 0.00 | 0.14 | 1.51 |
| scaffold00261 | maker-scaffold00261-snap-gene-0.36-mRNA-1 | AT1G05940.1 | AT1G05940.1 | 0.00 | 0.35 | 0.62 | 0.10 |
| scaffold00261 | maker-scaffold00261-snap-gene-1.35-mRNA-1 | AT3G59670.1 | AT3G59670.1 | −0.05 | 0.00 | 0.00 | −0.11 |
| scaffold00261 | maker-scaffold00261-snap-gene-1.36-mRNA-1 | AT4G37340.1; AT4G37360.1; AT4G37330.1; AT3G53280.1 | AT4G37370.1 | −1.43 | 0.14 | 0.15 | −1.44 |
| scaffold00261 | maker-scaffold00261-augustus-gene-1.31 | AT3G60340.1 | AT3G60340.1 | 0.33 | −0.41 | −0.29 | 0.44 |
| scaffold00261 | augustus_masked-scaffold00261-abinit-gene-1.1-mRNA-1 | AT2G13600.1; AT2G22410; AT5G19020.1 | AT1G08070.1 | 0.00 | 0.00 | 0.23 | 0.19 |
| scaffold05894 | augustus_masked-scaffold05894-abinit-gene-0.0-mRNA-1 | AT3G60220.1 | AT3G60220.1 | 0.00 | −0.04 | 0.78 | 0.92 |
| scaffold03566 | maker-scaffold03566-snap-gene-0.12-mRNA-1 | AT2G44745.1; AT4G39410 | AT2G44745.1 | 0.00 | 0.00 | 0.51 | 0.89 |
| scaffold04450 | maker-scaffold04450-augustus-gene-0.14-mRNA-1 | AT2G44520.1 | AT2G44520.1 | 0.17 | 0.50 | 0.57 | 0.23 |
| scaffold04450 | maker-scaffold04450-snap-gene-0.17-mRNA-1 | AT2G44525.1 | AT3G16000.1 | 0.26 | 0.00 | −0.24 | −0.23 |
| scaffold05894 | maker-scaffold05894-snap-gene-0.11-mRNA-1 | AT1G02180.1 | AT1G02180.1 | 0.00 | 0.00 | 0.00 | 0.22 |
| scaffold06031 | maker-scaffold06031-augustus-gene-0.16-mRNA-1 | - | AT3G26330.1 | 0.00 | 0.00 | 0.69 | 0.54 |
| scaffold06906 | maker-scaffold06906-snap-gene-0.18-mRNA-1 | AT2G35170.1; AT1G21920.1 | AT4G17080.1 | 0.31 | 0.20 | 0.00 | 0.04 |
| scaffold06906 | maker-scaffold06906-snap-gene-0.19-mRNA-1 | AT4G25110.1; AT1G02170.1 | AT1G02170.1 | 0.00 | 0.01 | 0.79 | 0.58 |
| scaffold06906 | maker-scaffold06906-snap-gene-0.20-mRNA-1 | AT3G10950.1; AT3G60245.1 | AT3G10950.1 | 0.28 | 0.13 | 0.52 | 0.65 |
| scaffold07319 | maker-scaffold07319-augustus-gene-0.10-mRNA-1 | AT2G39445.1 | AT3G09080.1 | 0.00 | 0.00 | 0.37 | 0.75 |
| scaffold07319 | maker-scaffold07319-augustus-gene-0.8-mRNA-1 | AT1G80770.1 | AT1G80770.1 | 0.00 | 0.00 | 0.00 | 0.00 |
| scaffold07319 | maker-scaffold07319-augustus-gene-0.8-mRNA-2 | AT1G80770.1 | AT1G80770.1 | 0.00 | 0.00 | 0.00 | 0.00 |
| scaffold07739 | maker-scaffold07739-augustus-gene-0.7-mRNA-1 | AT3G48110.1 | AT3G48110.1 | 0.09 | 0.00 | 0.00 | 0.13 |
| scaffold07739 | maker-scaffold07739-augustus-gene-0.7-mRNA-2 | AT3G48110.1 | AT3G48110.1 | 0.09 | 0.00 | 0.00 | 0.13 |
| scaffold10721 | maker-scaffold10721-snap-gene-0.12-mRNA-1 | AT3G13080.1; AT3G60160.1; AT1G04120.1; AT3G60970.1 | AT3G60160.1 | 0.00 | 0.00 | 0.00 | 0.17 |
| scaffold10655 | snap_masked-scaffold10655-abinit-gene-0.6-mRNA-1 | - | AT2G44930.1 | −0.06 | 0.41 | 2.27 | 0.34 |
| scaffold18444 | snap_masked-scaffold18444-abinit-gene-0.3-mRNA-1 | - | AT3G14470.1 | −0.10 | -0.09 | 0.16 | 0.35 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torello Marinoni, D.; Nishio, S.; Valentini, N.; Shirasawa, K.; Acquadro, A.; Portis, E.; Alma, A.; Akkak, A.; Pavese, V.; Cavalet-Giorsa, E.; et al. Development of High-Density Genetic Linkage Maps and Identification of Loci for Chestnut Gall Wasp Resistance in Castanea spp. Plants 2020, 9, 1048. https://doi.org/10.3390/plants9081048
Torello Marinoni D, Nishio S, Valentini N, Shirasawa K, Acquadro A, Portis E, Alma A, Akkak A, Pavese V, Cavalet-Giorsa E, et al. Development of High-Density Genetic Linkage Maps and Identification of Loci for Chestnut Gall Wasp Resistance in Castanea spp. Plants. 2020; 9(8):1048. https://doi.org/10.3390/plants9081048
Chicago/Turabian StyleTorello Marinoni, Daniela, Sogo Nishio, Nadia Valentini, Kenta Shirasawa, Alberto Acquadro, Ezio Portis, Alberto Alma, Aziz Akkak, Vera Pavese, Emile Cavalet-Giorsa, and et al. 2020. "Development of High-Density Genetic Linkage Maps and Identification of Loci for Chestnut Gall Wasp Resistance in Castanea spp." Plants 9, no. 8: 1048. https://doi.org/10.3390/plants9081048