Framework Phylogeny, Evolution and Complex Diversification of Chinese Oaks

 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Sequence Information of the Chinese Oaks
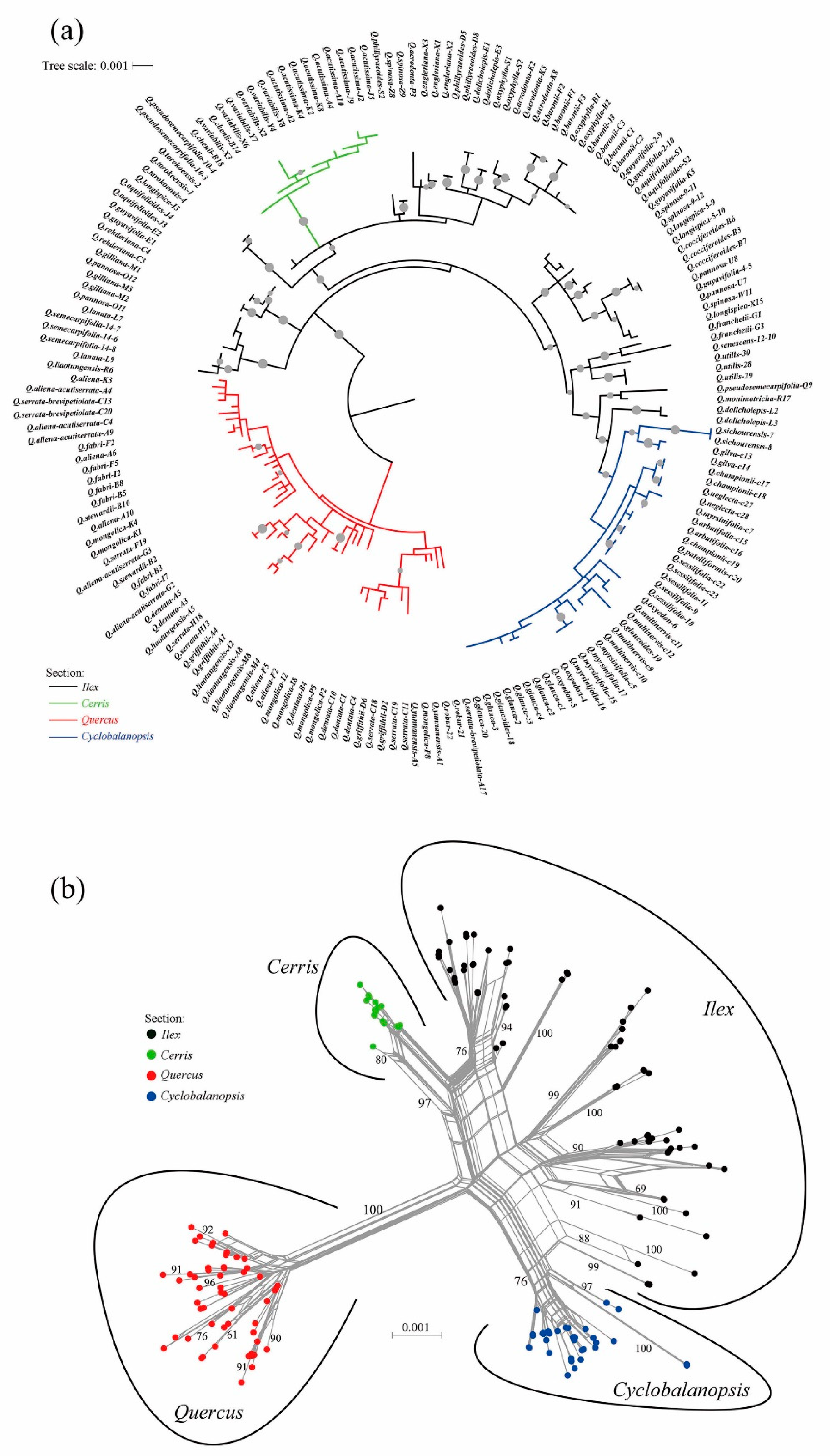
2.2. Infrageneric Phylogeny of the Chinese Oak’s Lineage
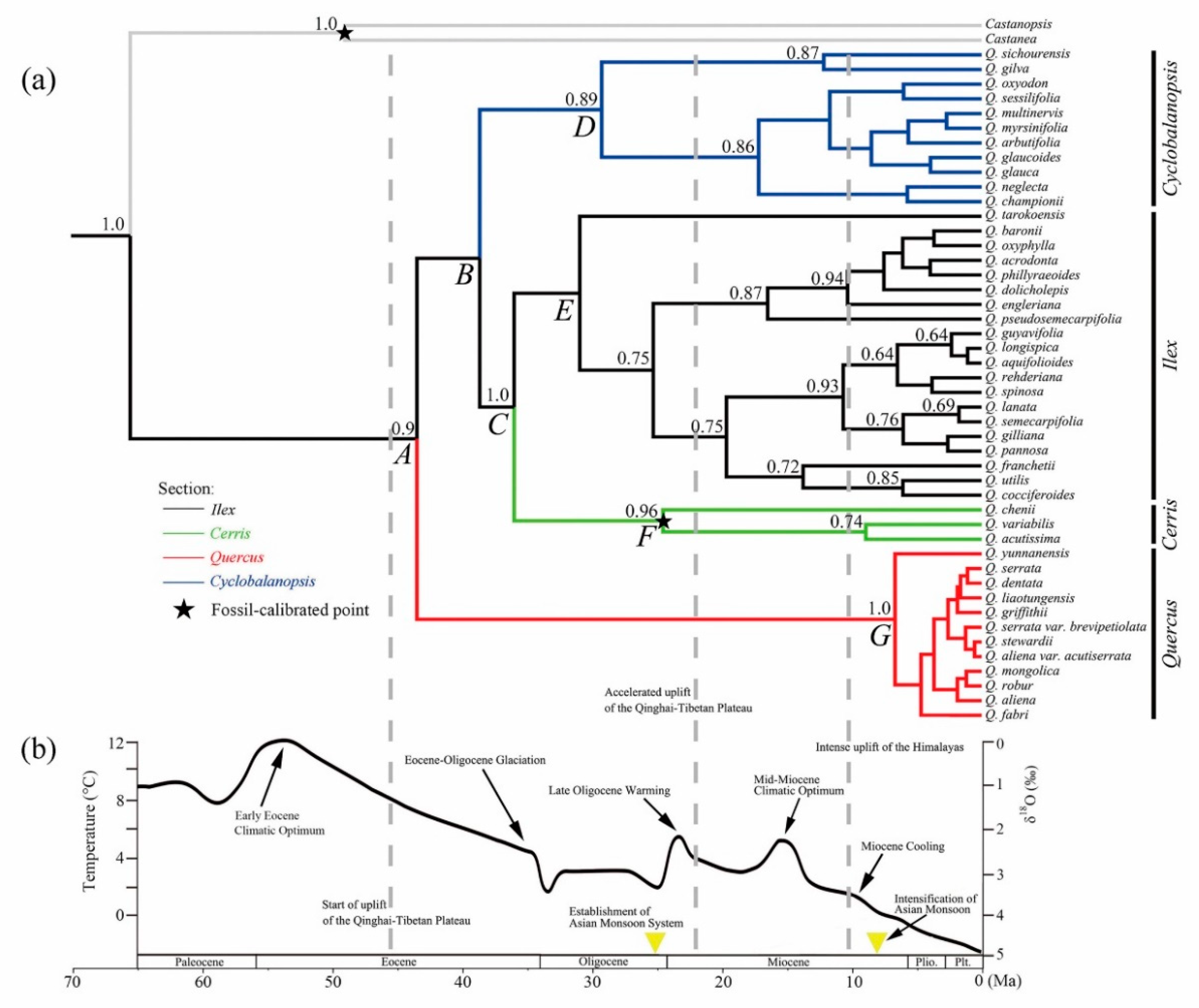
2.3. Evolutionary History of the Chinese Oaks
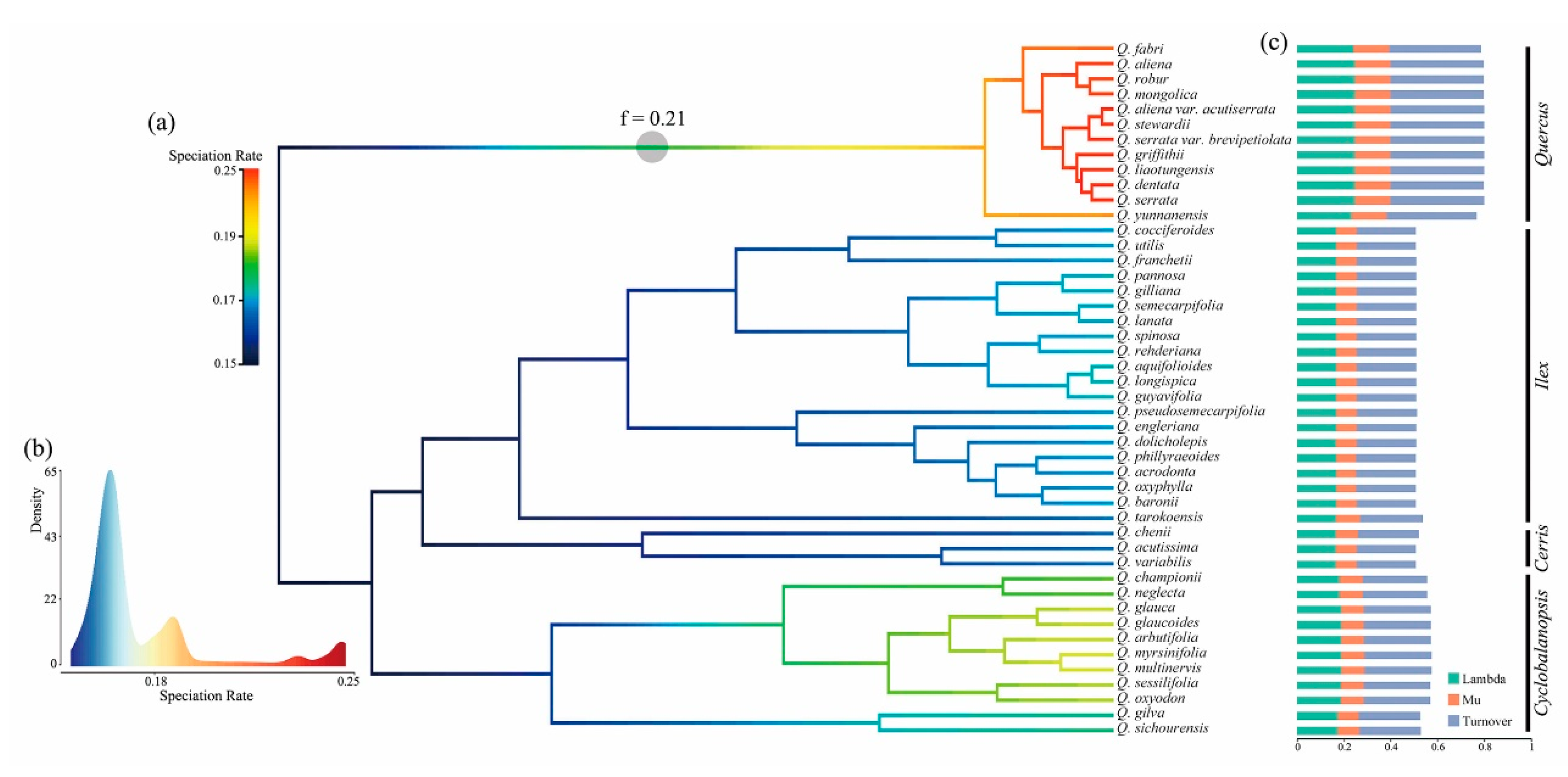
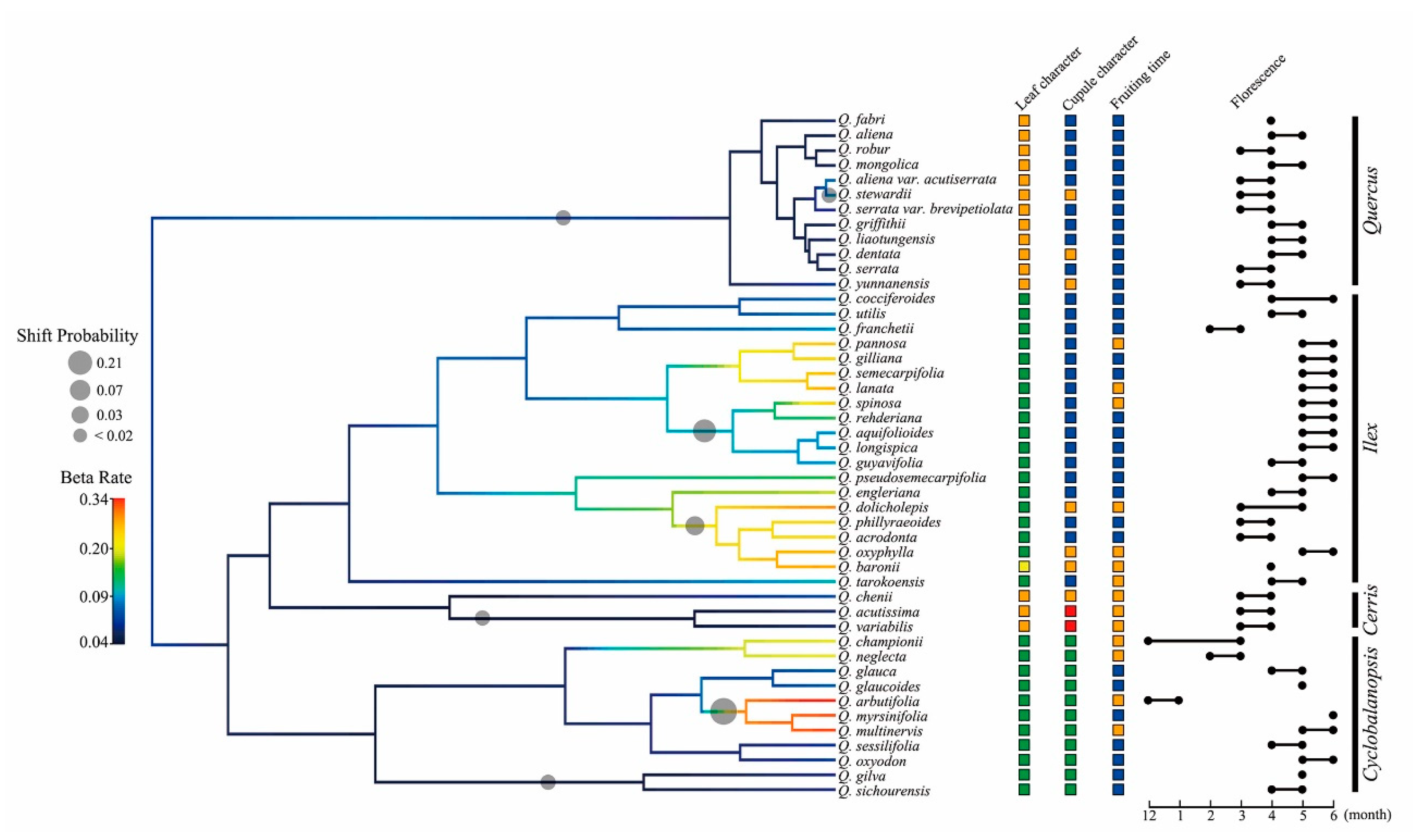
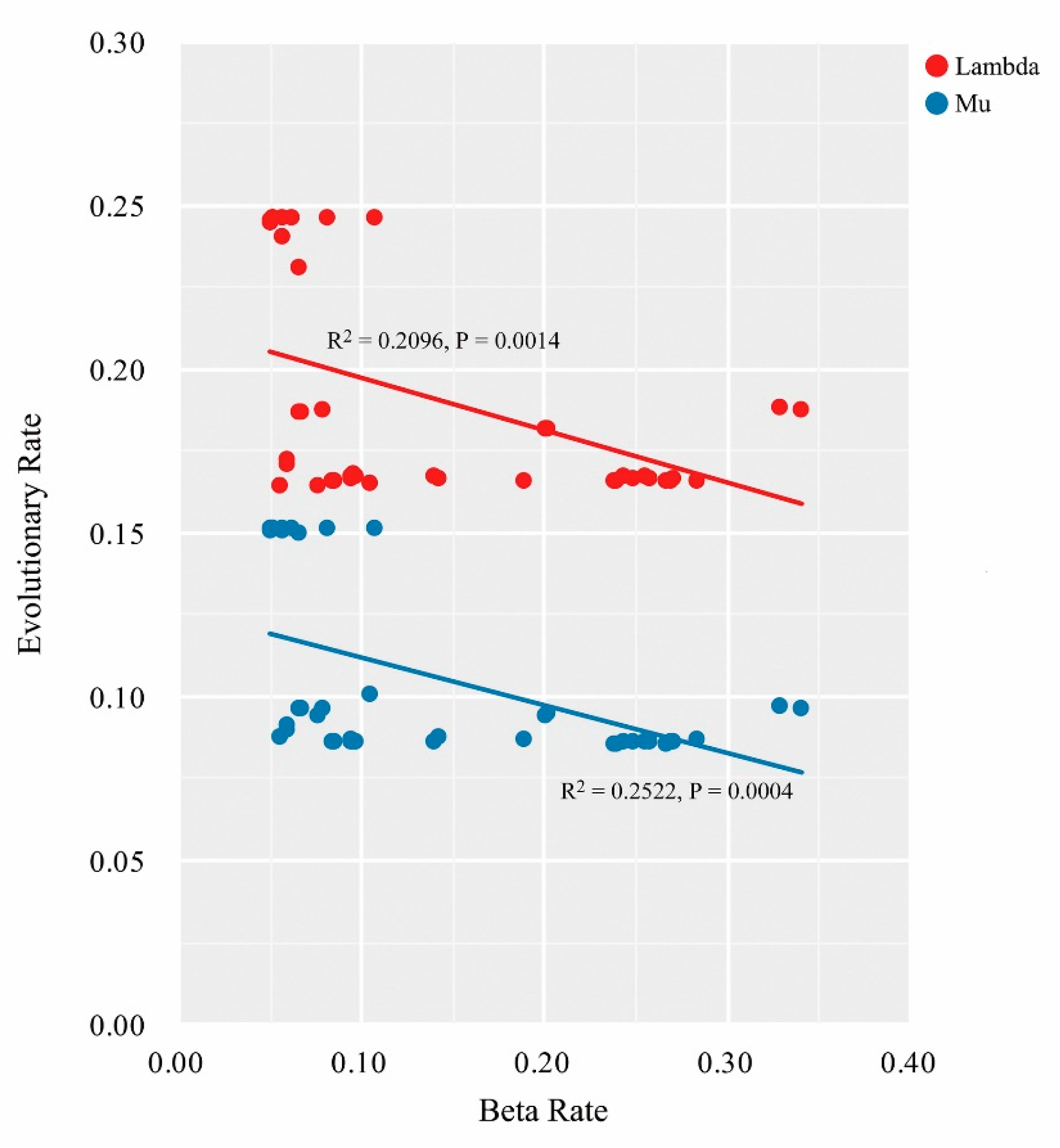
2.4. Lineage Diversification and Phenotypic Evolution of the Chinese Oak Species
3. Discussion
3.1. Infrageneric Phylogeny of Chinese Oaks
3.2. Evolutionary History of the Chinese Oaks
3.3. Complex Diversification of the Chinese Oak’s Lineage
4. Material and Methods
4.1. Species Information and DNA Extraction
4.2. PCR Amplification and Molecular Data Processing
4.3. Reconstruction of Infrageneric Phylogeny
4.4. Molecular Dating
4.5. Diversification Simulation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lortie, C.J.; Svenning, J.C. The diversity of diversity studies: Retrospectives and future directions. Ecography 2015, 38, 330–334. [Google Scholar] [CrossRef]
- Testo, W.L.; Sessa, E.; Barrington, D.S. The rise of the Andes promoted rapid diversification in Neotropical Phlegmariurus (Lycopodiaceae). New Phytol. 2019, 222, 604–613. [Google Scholar] [CrossRef]
- Ricklefs, R.E. Evolutionary diversification and the origin of the diversity-environment relationship. Ecology 2006, 87, S3–S13. [Google Scholar] [CrossRef]
- Svenning, J.C.; Borchsenius, F.; Bjorholm, S.; Balslev, H. High tropical net diversification drives the New World latitudinal gradient in palm (Arecaceae) species richness. J. Biogeogr. 2008, 35, 394–406. [Google Scholar] [CrossRef]
- Eaton, D.A.R.; Fenster, C.B.; Hereford, J.; Huang, S.Q.; Ree, R.H. Floral diversity and community structure in Pedicularis (Orobanchaceae). Ecology 2012, 93, S182–S194. [Google Scholar] [CrossRef]
- Donoghue, M.J. A phylogenetic perspective on the distribution of plant diversity. Proc. Natl. Acad. Sci. USA 2008, 105, 11549–11555. [Google Scholar] [CrossRef] [Green Version]
- Eaton, D.A.R.; Hipp, A.L.; González-Rodriguez, A.; Cavender-Bares, J. Historical introgression among the American live oaks and the comparative nature of tests for introgression. Evolution 2015, 69, 2587–2601. [Google Scholar] [CrossRef]
- Ortego, J.; Noguerales, V.; Gugger, P.F.; Sork, V.L. Evolutionary and demographic history of the Californian scrub white oak species complex: An integrative approach. Mol. Ecol. 2015, 24, 6188–6208. [Google Scholar] [CrossRef] [Green Version]
- Qian, H.; Wiens, J.J.; Zhang, J.; Zhang, Y.J. Evolutionary and ecological causes of species richness patterns in North American angiosperm trees. Ecography 2015, 38, 241–250. [Google Scholar] [CrossRef]
- McVay, J.D.; Hipp, A.L.; Manos, P.S. A genetic legacy of introgression confounds phylogeny and biogeography in oaks. Proc. R. Soc. B 2017, 284, 20170300. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.; Suzuki, N.; Tojo, K. Parallel evolution of an alpine type ecomorph in a scorpionfly: Independent adaptation to high-altitude environments in multiple mountain locations. Mol. Ecol. 2019, 28, 3225–3240. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, B.D.; Moreau, C.S. Defensive traits exhibit an evolutionary trade-off and drive diversification in ants. Evolution 2017, 71, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Manos, P.S.; Doyle, J.J.; Nixon, K.C. Phylogeny, biogeography, and processes of molecular differentiation in Quercus subgenus Quercus (Fagaceae). Mol. Phylogenet. Evol. 1999, 12, 333–349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kremer, A.; Abbott, A.G.; Carlson, J.E.; Manos, P.S.; Plomion, C.; Sisco, P.; Staton, M.E.; Ueno, S.; Vendramin, G.G. Genomics of Fagaceae. Tree Genet. Genomes 2012, 8, 583–610. [Google Scholar] [CrossRef] [Green Version]
- Cavender-Bares, J. Diversity, distribution and ecosystem services of the North American oaks. Int. Oaks 2016, 27, 37–48. [Google Scholar]
- Hipp, A.L.; Manos, P.S.; Gonzalez-Rodriguez, A.; Hahn, M.; Kaproth, M.; McVay, J.D.; Avalos, S.V.; Cavender-Bares, J. Sympatric parallel diversification of major oak clades in the Americas and the origins of Mexican species diversity. New Phytol. 2018, 217, 439–452. [Google Scholar] [CrossRef] [Green Version]
- Chasse, B. Updated classification of oaks: A summary. Int. Oaks 2018, 29, 11–18. [Google Scholar]
- Burger, W.C. The species concept in Quercus. TAXON 1975, 24, 45–50. [Google Scholar] [CrossRef]
- Nixon, K.C. Infrageneric classification of Quercus (Fagaceae) and typification of sectional names. Ann. Sci. For. 1993, 50, 25s–34s. [Google Scholar] [CrossRef] [Green Version]
- Denk, T.; Grimm, G.W. The oaks of western Eurasia: Traditional classifications and evidence from two nuclear markers. TAXON 2010, 59, 351–366. [Google Scholar] [CrossRef]
- Simeone, M.C.; Piredda, R.; Papini, A.; Vessella, F.; Schirone, B. Application of plastid and nuclear markers to DNA barcoding of Euro-Mediterranean oaks (Quercus, Fagaceae): Problems, prospects and phylogenetic implications. Bot. J. Linn. Soc. 2013, 172, 478–499. [Google Scholar] [CrossRef] [Green Version]
- Crowl, A.A.; Manos, P.S.; McVay, J.D.; Lemmon, A.R.; Lemmon, E.M.; Hipp, A.L. Uncovering the genomic signature of ancient introgression between white oak lineages (Quercus). New Phytol. 2020, 226, 1158–1170. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.K. Origin, phylogeny and dispersal of Quercus from China. Acta Bot. Yunnanica 1992, 14, 227–236. [Google Scholar]
- Pu, C.X.; Zhou, Z.K.; Luo, Y. A cladistic analysis of Quercus (Fagaceae) in China based on leaf epidermis and architecture. Acta Bot. Yunnanica 2002, 24, 689–698. [Google Scholar]
- Deng, M.; Hipp, A.L.; Song, Y.G.; Li, Q.S.; Coombes, A.; Cotton, A. Leaf epidermal features of Quercus subgenus Cyclobalanopsis (Fagaceae) and their systematic significance. Bot. J. Linn. Soc. 2014, 176, 224–259. [Google Scholar] [CrossRef] [Green Version]
- Hipp, A.L.; Manos, P.S.; Hahn, M.; Avishai, M.; Bodenes, C.; Cavender-Bares, J.; Crowl, A.A.; Deng, M.; Denk, T.; Fitz-Gibbon, S.; et al. Genomic landscape of the global oak phylogeny. New Phytol. 2020, 226, 1198–1212. [Google Scholar] [CrossRef]
- Denk, T.; Grimm, G.W.; Manos, P.S.; Deng, M.; Hipp, A.L. An updated infrageneric classification of the oaks: Review of previous taxonomic schemes and synthesis of evolutionary patterns. In Tree Physiology. Oaks Physiological Ecology. Exploring the Functional Diversity of Genus Quercus L.; Gil-Pelegrin, E., Peguero-Pina, J.J., Sancho-Knapik, D., Eds.; Springer: Cham, Switzerland, 2017; pp. 13–38. [Google Scholar]
- Yang, J.; Vazquez, L.; Chen, X.D.; Li, H.M.; Zhang, H.; Liu, Z.L.; Zhao, G.F. Development of chloroplast and nuclear DNA markers for Chinese oaks (Quercus subgenus Quercus) and assessment of their utility as DNA barcodes. Front. Plant Sci. 2017, 8, 816. [Google Scholar] [CrossRef]
- Yan, M.X.; Xiong, Y.S.; Liu, R.B.; Deng, M.; Song, J.J. The application and limitation of universal chloroplast markers in discriminating East Asian evergreen oaks. Front. Plant Sci. 2018, 9, 569. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.K. The fossil history of Quercus. Acta Bot. Yunnanica 1993, 15, 21–33. [Google Scholar]
- Deng, M.; Jiang, X.L.; Hipp, A.L.; Manos, P.S.; Hahn, M. Phylogeny and biogeography of East Asian evergreen oaks (Quercus section Cyclobalanopsis; Fagaceae): Insights into the Cenozoic history of evergreen broad-leaved forests in subtropical Asia. Mol. Phylogenet. Evol. 2018, 119, 170–181. [Google Scholar] [CrossRef]
- Jiang, X.L.; Hipp, A.L.; Deng, M.; Su, T.; Zhou, Z.K.; Yan, M.X. East Asian origins of European holly oaks (Quercus section Ilex Loudon) via the Tibet-Himalaya. J. Biogeogr. 2019, 46, 2203–2214. [Google Scholar] [CrossRef]
- Xu, X.T.; Wang, Z.H.; Rahbek, C.; Lessard, J.P.; Fang, J.Y. Evolutionary history influences the effects of water-energy dynamics on oak diversity in Asia. J. Biogeogr. 2013, 40, 2146–2155. [Google Scholar] [CrossRef]
- Zeng, Y.F.; Liao, W.J.; Petit, R.J.; Zhang, D.Y. Geographic variation in the structure of oak hybrid zones provides insights into the dynamics of speciation. Mol. Ecol. 2011, 20, 4995–5011. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.F.; Wang, W.T.; Liao, W.J.; Wang, H.F.; Zhang, D.Y. Multiple glacial refugia for cool-temperate deciduous trees in northern East Asia: The Mongolian oak as a case study. Mol. Ecol. 2015, 24, 5676–5691. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.M.; Zhang, X.X.; Kang, H.Z.; Sun, X.; Yin, S.; Du, H.M.; Yamanaka, N.; Gapare, W.; Wu, H.X.; Liu, C.J. Phylogeography of Quercus variabilis based on chloroplast DNA sequence in East Asia: Multiple glacial refugia and mainland-migrated island populations. PLoS ONE 2012, 7, e47268. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Deng, M.; Jiang, X.L.; Westwood, M.; Song, Y.G.; Turkington, R. Phylogeography of Quercus glauca (Fagaceae), a dominant tree of East Asian subtropical evergreen forests, based on three chloroplast DNA interspace sequences. Tree Genet. Genomes 2014, 11, 805. [Google Scholar] [CrossRef]
- Du, F.K.; Hou, M.; Wang, W.T.; Mao, K.S.; Hampe, A. Phylogeography of Quercus aquifolioides provides novel insights into the Neogene history of a major global hotspot of plant diversity in south-west China. J. Biogeogr. 2017, 44, 294–307. [Google Scholar] [CrossRef]
- Jiang, X.L.; Gardner, E.M.; Meng, H.H.; Deng, M.; Xu, G.B. Land bridges in the Pleistocene contributed to flora assembly on the continental islands of South China: Insights from the evolutionary history of Quercus championii. Mol. Phylogenet. Evol. 2019, 132, 36–45. [Google Scholar] [CrossRef]
- Ju, M.M.; Feng, L.; Yang, J.; Yang, Y.C.; Chen, X.D.; Zhao, G.F. Evaluating population genetic structure and demographic history of Quercus spinosa (Fagaceae) based on specific length amplified fragment sequencing. Front. Genet. 2019, 10, 965. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.D.; Yang, J.; Feng, L.; Zhou, T.; Zhang, H.; Li, H.M.; Bai, G.Q.; Meng, X.; Li, Z.H.; Zhao, G.F. Phylogeography and population dynamics of an endemic oak (Quercus fabri Hance) in subtropical China revealed by molecular data and ecological niche modeling. Tree Genet. Genomes 2020, 16, 2. [Google Scholar] [CrossRef]
- Yang, J.; Vazquez, L.; Feng, L.; Liu, Z.L.; Zhao, G.F. Climatic and soil factors shape the demographical history and genetic diversity of a deciduous oak (Quercus liaotungensis) in Northern China. Front. Plant Sci. 2018, 9, 1534. [Google Scholar] [CrossRef] [PubMed]
- Favre, A.; Packert, M.; Pauls, S.U.; Jahnig, S.C.; Uhl, D.; Michalak, I.; Muellner-Riehl, A.N. The role of the uplift of the Qinghai-Tibetan Plateau for the evolution of Tibetan biotas. Biol. Rev. 2015, 90, 236–253. [Google Scholar] [CrossRef]
- Zachos, J.; Pagani, M.; Sloan, I.; Thomas, E.; Billups, K. Trends, rhythms, and aberrations in global climate 65 Ma to present. Science 2001, 292, 686–693. [Google Scholar] [CrossRef] [PubMed]
- Simeone, M.C.; Grimm, G.W.; Papini, A.; Vessella, F.; Cardoni, S.; Tordoni, E.; Piredda, R.; Franc, A.; Denk, T. Plastome data reveal multiple geographic origins of Quercus Group Ilex. PeerJ 2016, 4, e1897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pham, K.K.; Hipp, A.L.; Manos, P.S.; Cronn, R.C. A time and a place for everything: Phylogenetic history and geography as joint predictors of oak plastome phylogeny. Genome 2017, 60, 720–732. [Google Scholar] [CrossRef] [Green Version]
- Manos, P.S.; Stanford, A.M. The historical biogeography of Fagaceae: Tracking the Tertiary history of temperate and subtropical forests of the Northern Hemisphere. Int. J. Plant Sci. 2001, 162, S77–S93. [Google Scholar] [CrossRef]
- Hubert, F.; Grimm, G.W.; Jousselin, E.; Berry, V.; Franc, A.; Kremer, A. Multiple nuclear genes stabilize the phylogenetic backbone of the genus Quercus. Syst. Biodivers. 2014, 12, 405–423. [Google Scholar] [CrossRef]
- Feng, L.; Zheng, Q.J.; Qian, Z.Q.; Yang, J.; Zhang, Y.P.; Li, Z.H.; Zhao, G.F. Genetic structure and evolutionary history of three alpine sclerophyllous oaks in east Himalaya-Hengduan Mountains and adjacent regions. Front. Plant Sci. 2016, 7, 1688. [Google Scholar] [CrossRef] [Green Version]
- An, M.; Deng, M.; Zheng, S.S.; Jiang, X.L.; Song, Y.G. Introgression threatens the genetic diversity of Quercus austrocochinchinensis (Fagaceae), an endangered oak: A case inferred by molecular markers. Front. Plant Sci. 2017, 8, 229. [Google Scholar] [CrossRef] [Green Version]
- Cavender-Bares, J.; Gonzalez-Rodriguez, A.; Eaton, D.A.R.; Hipp, A.L.; Beulke, A.; Manos, P.S. Phylogeny and biogeography of the American live oaks (Quercus subsection Virentes): A genomic and population genetics approach. Mol. Ecol. 2015, 24, 3668–3687. [Google Scholar] [CrossRef]
- Meng, H.H.; Su, T.; Gao, X.Y.; Li, J.; Jiang, X.L.; Sun, H.; Zhou, Z.K. Warm-cold colonization: Response of oaks to uplift of the Himalaya-Hengduan Mountains. Mol. Ecol. 2017, 26, 3276–3294. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.Q.; Duan, Y.W.; Hao, G.; Ge, X.J.; Sun, H. Evolutionary history and underlying adaptation of alpine plants on the Qinghai-Tibet Plateau. J. Syst. Evol. 2014, 52, 241–249. [Google Scholar] [CrossRef]
- Renner, S.S. Available data point to a 4-km-high Tibetan Plateau by 40 Ma, but 100 molecular-clock papers have linked supposed recent uplift to young node ages. J. Biogeogr. 2016, 43, 1479–1487. [Google Scholar] [CrossRef]
- Su, T.; Spicer, R.A.; Li, S.H.; Xu, H.; Huang, J.; Sherlock, S.; Huang, Y.J.; Li, S.F.; Wang, L.; Jia, L.B.; et al. Uplift, climate and biotic changes at the Eocene-Oligocene transition in south-eastern Tibet. Natl. Sci. Rev. 2019, 6, 495–504. [Google Scholar] [CrossRef] [Green Version]
- Simeone, M.C.; Cardoni, S.; Piredda, R.; Imperatori, F.; Avishai, M.; Grimm, G.W.; Denk, T. Comparative systematics and phylogeography of Quercus Section Cerris in western Eurasia: Inferences from plastid and nuclear DNA variation. PeerJ 2018, 6, e5793. [Google Scholar] [CrossRef] [Green Version]
- Cavender-Bares, J.; Kothari, S.; Meireles, J.E.; Kaproth, M.A.; Manos, P.S.; Hipp, A.L. The role of diversification in community assembly of the oaks (Quercus L.) across the continental U.S. Am. J. Bot. 2018, 105, 565–586. [Google Scholar] [CrossRef] [Green Version]
- Leroy, T.; Louvet, J.M.; Lalanne, C.; Provost, G.L.; Labadie, K.; Aury, J.M.; Delzon, S.; Plomion, C.; Kremer, A. Adaptive introgression as a driver of local adaptation to climate in European white oaks. New Phytol. 2020, 226, 1171–1182. [Google Scholar] [CrossRef]
- Rabosky, D.L.; Adams, D.C. Rates of morphological evolution are correlated with species richness in salamanders. Evolution 2012, 66, 1807–1818. [Google Scholar] [CrossRef]
- Moore, B.R.; Donoghue, M.J. Correlates of diversification in the plant clade Dipsacales: Geographic movement and evolutionary innovations. Am. Nat. 2007, 170, S28–S35. [Google Scholar] [CrossRef]
- Wellenreuther, M.; Merot, C.; Berdan, E.; Bernatchez, L. Going beyond SNPs: The role of structural genomic variants in adaptive evolution and species diversification. Mol. Ecol. 2019, 28, 1203–1209. [Google Scholar] [CrossRef] [Green Version]
- Nagamitsu, T.; Uchiyama, K.; Izuno, A.; Shimizu, H.; Nakanishi, A. Environment-dependent introgression from Quercus dentata to a coastal ecotype of Quercus mongolica var. crispula in northern Japan. New Phytol. 2020, 226, 1018–1028. [Google Scholar] [CrossRef] [PubMed]
- Koehler, K.; Center, A.; Cavender-Bares, J. Evidence for a freezing tolerance-growth rate trade-off in the live oaks (Quercus series Virentes) across the tropical-temperate divide. New Phytol. 2012, 193, 730–744. [Google Scholar] [CrossRef]
- Cannon, C.H.; Petit, R.J. The oak syngameon: More than the sum of its parts. New Phytol. 2020, 226, 978–983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Di, X.Y.; Meng, X.; Feng, L.; Liu, Z.L.; Zhao, G.F. Phylogeography and evolution of two closely related oak species (Quercus) from north and northeast China. Tree Genet. Genomes 2016, 12, 89. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Li, H.P. A new test for detecting recent positive selection that is free from the confounding impacts of demography. Mol. Biol. Evol. 2011, 28, 365–375. [Google Scholar] [CrossRef] [Green Version]
- Rozas, J.; Sanchez-Delbarrio, J.C.; Messeguer, X.; Rozas, R. DnaSP, DNA polymorphism analyses by the coalescent and other methods. Bioinformatics 2003, 19, 2496–2497. [Google Scholar] [CrossRef]
- Minh, B.Q.; Nguyen, M.A.T.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2014, 32, 268–274. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) V4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [Green Version]
- Bryant, D.; Moulton, V. Neighbor-Net: An agglomerative method for the construction of phylogenetic networks. Mol. Biol. Evol. 2004, 21, 255–265. [Google Scholar] [CrossRef]
- Suchard, M.A.; Lemey, P.; Baele, G.; Ayres, D.L.; Drummond, A.J.; Rambaut, A. Bayesian phylogenetic and phylodynamic data integration using BEAST 1.10. Virus Evol. 2018, 4, vey016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Z.K. Fossils of the Fagaceae and their implications in systematics and biogeography. Acta Phytotaxon. Sin. 1999, 37, 369–385. [Google Scholar]
- Rabosky, D.L. Automatic detection of key innovations, rate shifts, and diversity-dependence on phylogenetic trees. PLoS ONE 2014, 9, e89543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rabosky, D.L.; Grundler, M.; Anderson, C.; Title, P.; Shi, J.J.; Brown, J.W.; Huang, H.T.; Larson, J.G. BAMMtools: An R package for the analysis of evolutionary dynamics on phylogenetic trees. Methods Ecol. Evol. 2014, 5, 701–707. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2011. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Section | Taxon | Samples | Leaf Character | Cupule Character | Florescence Range | Fruiting Period with Florescence |
|---|---|---|---|---|---|---|
| Quercus | Quercus aliena Blume | 9 | Deciduous | Ovate bract | Apr.–May | Present year |
| Q. aliena var. acutiserrata Maximowicz ex Wenzig | 9 | Deciduous | Ovate bract | Mar.–Apr. | Present year | |
| Q. dentata Thunb. | 9 | Deciduous | Lanceolate bract | Apr.–May | Present year | |
| Q. fabri Hance | 9 | Deciduous | Ovate bract | Apr. | Present year | |
| Q. griffithii Hooker and Thomson ex Miquel | 6 | Deciduous | Ovate bract | Apr.–May | Present year | |
| Q. liaotungensis Koidz. (or Q. wutaishanica Blume) | 9 | Deciduous | Ovate bract | Apr.–May | Present year | |
| Q. mongolica Fischer ex Ledebour | 9 | Deciduous | Ovate bract | Apr.–May | Present year | |
| Q. robur L. † | 2 | Deciduous | Ovate bract | Mar.–Apr. | Present year | |
| Q. serrata Murray | 8 | Deciduous | Ovate bract | Mar.–Apr. | Present year | |
| Q. serrata var. brevipetiolata (A. DC.) Nakai | 6 | Deciduous | Ovate bract | Mar.–Apr. | Present year | |
| Q. stewardii Rehd. | 3 | Deciduous | Lanceolate bract | Mar.–Apr. | Present year | |
| Q. yunnanensis Franchet | 3 | Deciduous | Lanceolate bract | Mar.–Apr. | Present year | |
| Ilex | Q. acrodonta Seemen | 6 | Evergreen | Ovate bract | Mar.–Apr. | Present year |
| Q. aquifolioides Rehd. and Wils. | 8 | Evergreen | Ovate bract | May–Jun. | Present year | |
| Q. baronii Skan | 9 | Semievergreen | Lanceolate bract | Apr. | Following year | |
| Q. cocciferoides Hand.-Mazz. | 3 | Evergreen | Ovate bract | Apr.–Jun. | Present year | |
| Q. dolicholepis A. Camus | 6 | Evergreen | Lanceolate bract | Mar.–May | Following year | |
| Q. engleriana Seemen | 3 | Evergreen | Ovate bract | Apr.–May | Present year | |
| Q. franchetii Skan | 3 | Evergreen | Ovate bract | Feb.–Mar. | Present year | |
| Q. gilliana Rehd. and Wils. | 3 | Evergreen | Ovate bract | May–Jun. | Present year | |
| Q. guyavifolia H. Leveille | 8 | Evergreen | Ovate bract | Apr.–May | Present year | |
| Q. lanata Smith | 3 | Evergreen | Ovate bract | May–Jun. | Following year | |
| Q. longispica (Hand.-Mazz.) A. Camus | 6 | Evergreen | Ovate bract | May–Jun. | Present year | |
| Q. monimotricha Hand.-Mazz. | 4 | Evergreen | Ovate bract | May–Jun. | Following year | |
| Q. oxyphylla (E. H. Wilson) Hand.-Mazz. | 6 | Evergreen | Lanceolate bract | May–Jun. | Following year | |
| Q. pannosa Hand.-Mazz. | 4 | Evergreen | Ovate bract | May–Jun. | Following year | |
| Q. phillyraeoides A. Gray | 6 | Evergreen | Ovate bract | Mar.–Apr. | Present year | |
| Q. pseudosemecarpifolia A. Camus | 6 | Evergreen | Ovate bract | May–Jun. | Present year | |
| Q. rehderiana Hand.-Mazz. | 3 | Evergreen | Ovate bract | May–Jun. | Present year | |
| Q. semecarpifolia Smith | 3 | Evergreen | Ovate bract | May–Jun. | Present year | |
| Q. senescens Hand.-Mazz. | 10 | Evergreen | Ovate bract | Mar.–May | Present year | |
| Q. spinosa David ex Franchet | 15 | Evergreen | Ovate bract | May–Jun. | Following year | |
| Q. tarokoensis Hayata | 4 | Evergreen | Ovate bract | Apr.–May | Following year | |
| Q. utilis Hu and W. C. Cheng | 3 | Evergreen | Ovate bract | Apr.–May | Present year | |
| Cerris | Q. acutissima Carruth. | 9 | Deciduous | Subulate bract | Mar.–Apr. | Following year |
| Q. chenii Nakai | 3 | Deciduous | Lanceolate bract | Mar.–Apr. | Following year | |
| Q. variabilis Blume | 6 | Deciduous | Subulate bract | Mar.–Apr. | Following year | |
| Cyclobalanopsis | Q. arbutifolia Hickel and A. Camus | 2 | Evergreen | Ring shaped bract | Dec.–Jan. | Following year |
| Q. championii (Bentham) Oersted | 3 | Evergreen | Ring shaped bract | Dec.–Mar. | Following year | |
| Q. gilva (Blume) Oersted | 5 | Evergreen | Ring shaped bract | May | Present year | |
| Q. glauca (Thunberg) Oersted | 7 | Evergreen | Ring shaped bract | Apr.–May | Present year | |
| Q. glaucoides Schottky | 2 | Evergreen | Ring shaped bract | May | Present year | |
| Q. macrocalyx Hickel and A. Camus | 2 | Evergreen | Ring shaped bract | Mar.–Apr. | Present year | |
| Q. multinervis W. C. Cheng and T. Hong | 4 | Evergreen | Ring shaped bract | May–Jun. | Following year | |
| Q. myrsinifolia (Blume) Oersted | 7 | Evergreen | Ring shaped bract | Jun. | Present year | |
| Q. neglecta Schottky | 2 | Evergreen | Ring shaped bract | Feb.–Mar. | Following year | |
| Q. oxyodon (Miquel) Oersted | 3 | Evergreen | Ring shaped bract | May–Jun. | Present year | |
| Q. patelliformis (Chun) Y. C. Hsu and H. W. Jen | 2 | Evergreen | Ring shaped bract | May–Jun. | Present year | |
| Q. sessilifolia (Blume) Schottky | 5 | Evergreen | Ring shaped bract | Apr.–May | Present year | |
| Q. sichourensis Hu | 2 | Evergreen | Ring shaped bract | Apr.–May | Present year |
| Infrageneric Event | Description (Node) | Time Estimation (Ma) | Mean Speciation/Extinction/Turnover Rate (Species/Myr) | Related Climatic/Geological Event † |
|---|---|---|---|---|
| Clade divergence | New World clade–Old World clade (A) | 43.54 (95% HPD: 25.02–64.92) | -- | Early uplift of the Qinghai-Tibetan Plateau |
| Cyclobalanopsis-(Ilex + Cerris) (B) | 38.69 | -- | Eocene–Oligocene Glaciation | |
| Ilex-Cerris (C) | 36.04 (95% HPD: 28.37–52.00) | -- | Eocene–Oligocene Glaciation | |
| Section diversification | Cyclobalanopsis (D) | 29.29 (95% HPD: 15.56–46.97) | 0.1899/0.0934/0.2833 | Oligocene Glaciation |
| Ilex (E) | 30.97 | 0.1759/0.0865/0.2624 | Oligocene Glaciation | |
| Cerris (F) | 24.56 (95% HPD: 18.47–30.44) | 0.1730/0.0874/0.2604 | Late Oligocene Warming/Establishment of Asian Monsoon | |
| Quercus (G) | 6.66 (95% HPD: 2.61–15.91) | 0.2496/0.1508/0.4004 | Miocene cooling/Intensification of Asian Monsoon |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, J.; Guo, Y.-F.; Chen, X.-D.; Zhang, X.; Ju, M.-M.; Bai, G.-Q.; Liu, Z.-L.; Zhao, G.-F. Framework Phylogeny, Evolution and Complex Diversification of Chinese Oaks. Plants 2020, 9, 1024. https://doi.org/10.3390/plants9081024
Yang J, Guo Y-F, Chen X-D, Zhang X, Ju M-M, Bai G-Q, Liu Z-L, Zhao G-F. Framework Phylogeny, Evolution and Complex Diversification of Chinese Oaks. Plants. 2020; 9(8):1024. https://doi.org/10.3390/plants9081024
Chicago/Turabian StyleYang, Jia, Yu-Fan Guo, Xiao-Dan Chen, Xiao Zhang, Miao-Miao Ju, Guo-Qing Bai, Zhan-Lin Liu, and Gui-Fang Zhao. 2020. "Framework Phylogeny, Evolution and Complex Diversification of Chinese Oaks" Plants 9, no. 8: 1024. https://doi.org/10.3390/plants9081024