Antioxidant and Biological Activities of Acacia saligna and Lawsonia inermis Natural Populations
 ,
,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Preparation
2.2. Analyses of Phenolic Compounds
2.3. Antioxidant Activity
2.4. Detection Intercellular ROS Accumulation
2.5. Anticancer Activities
2.6. Antibacterial Effect
2.7. Antifungal Effect
3. Results
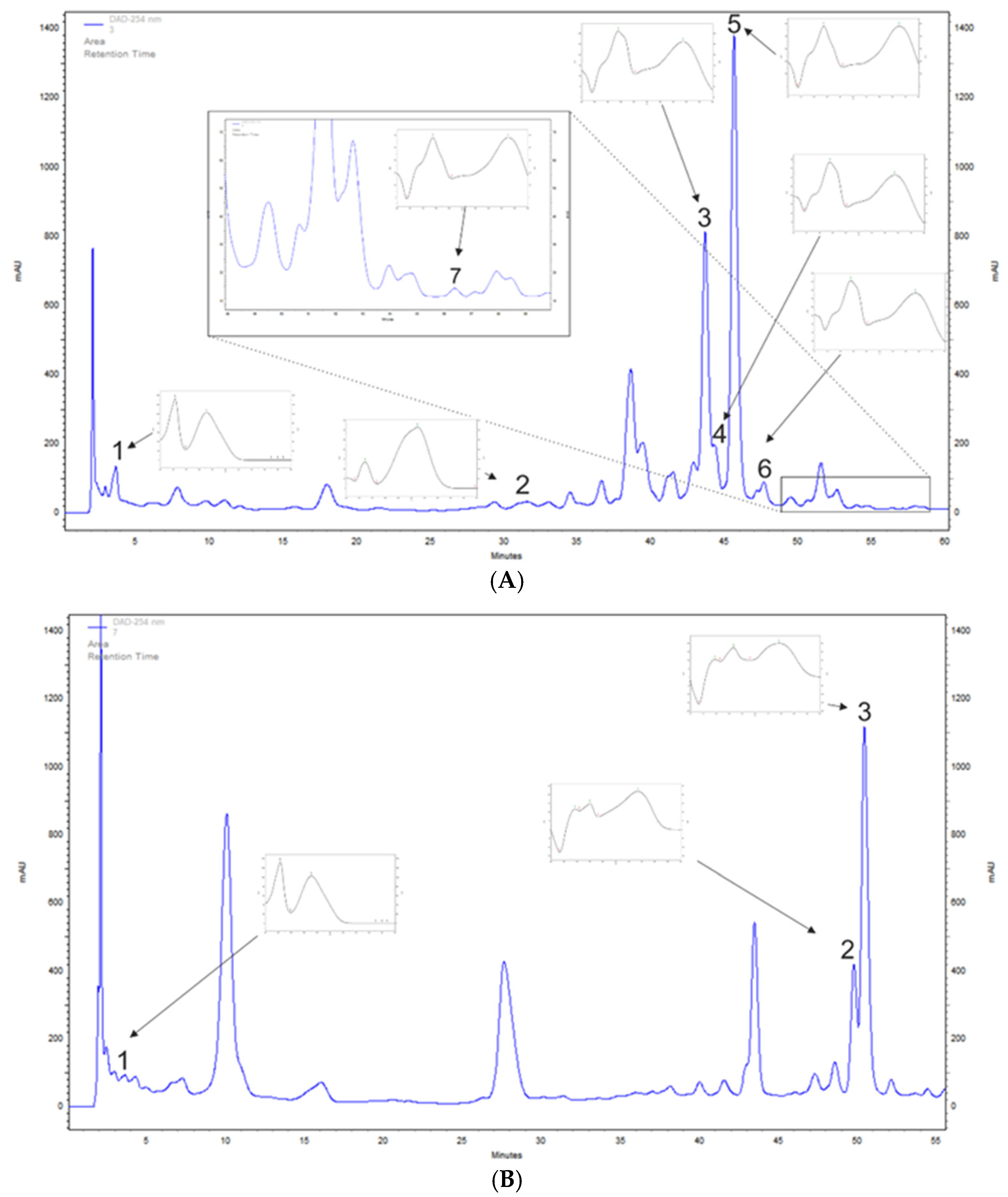
3.1. A. saligna and L. inermis Polyphenol Profiling of Leaf Extracts
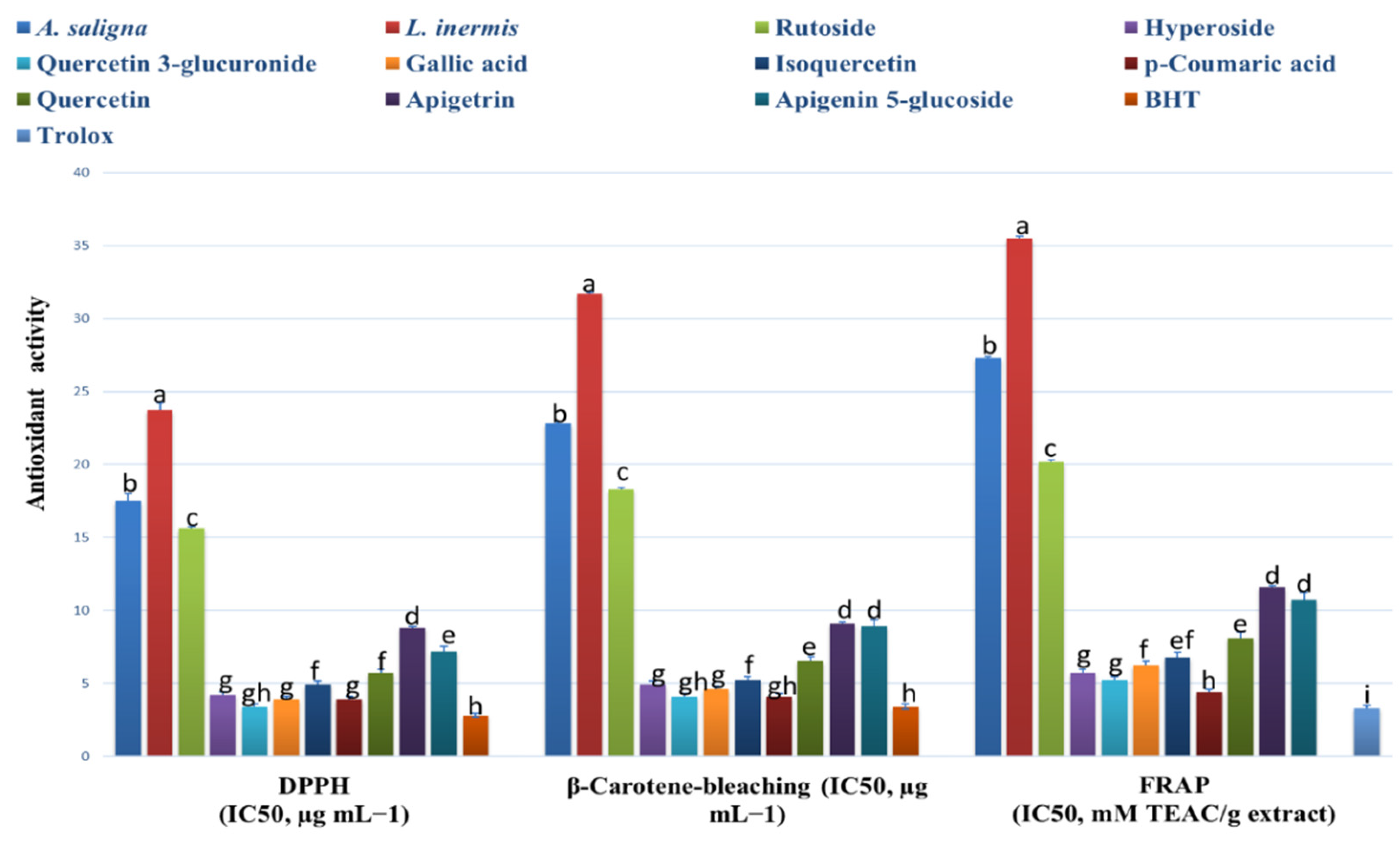
3.2. Antioxidant Effects
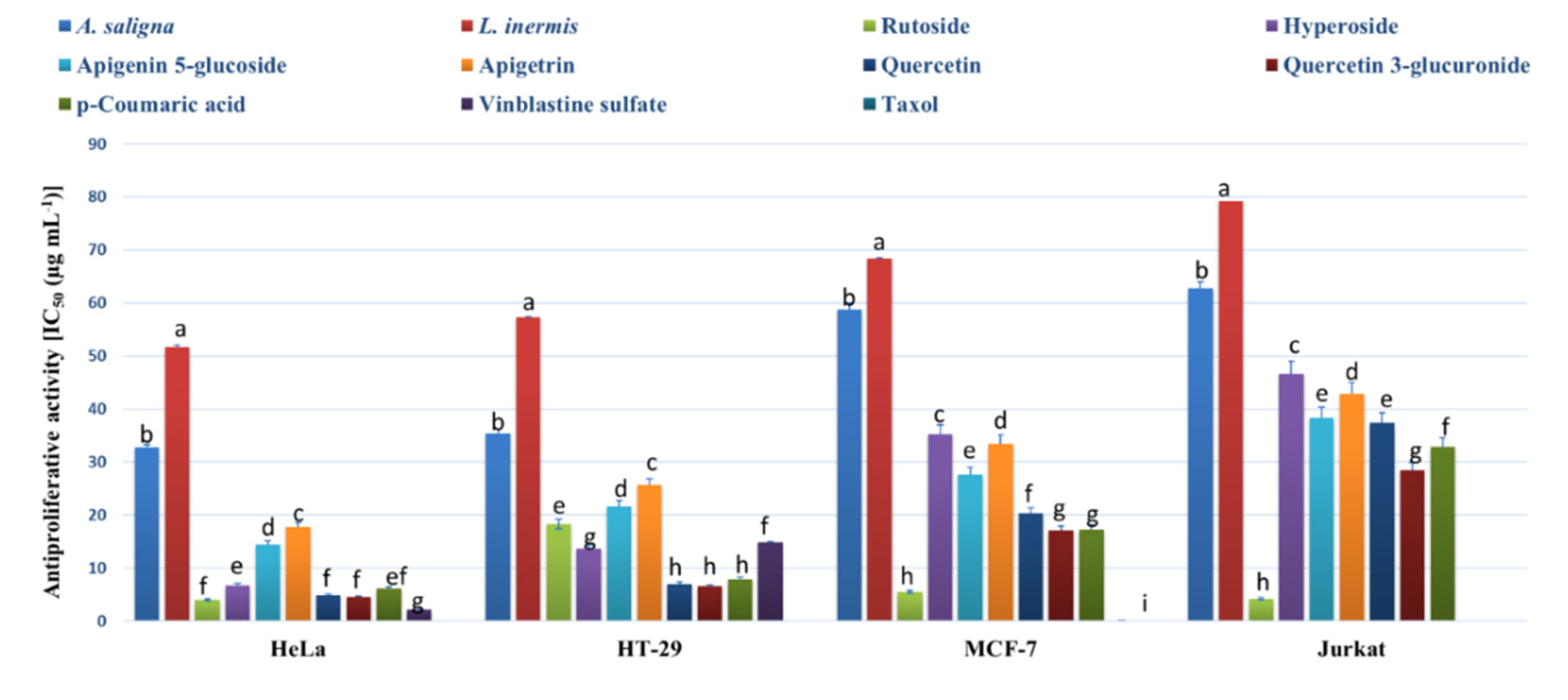
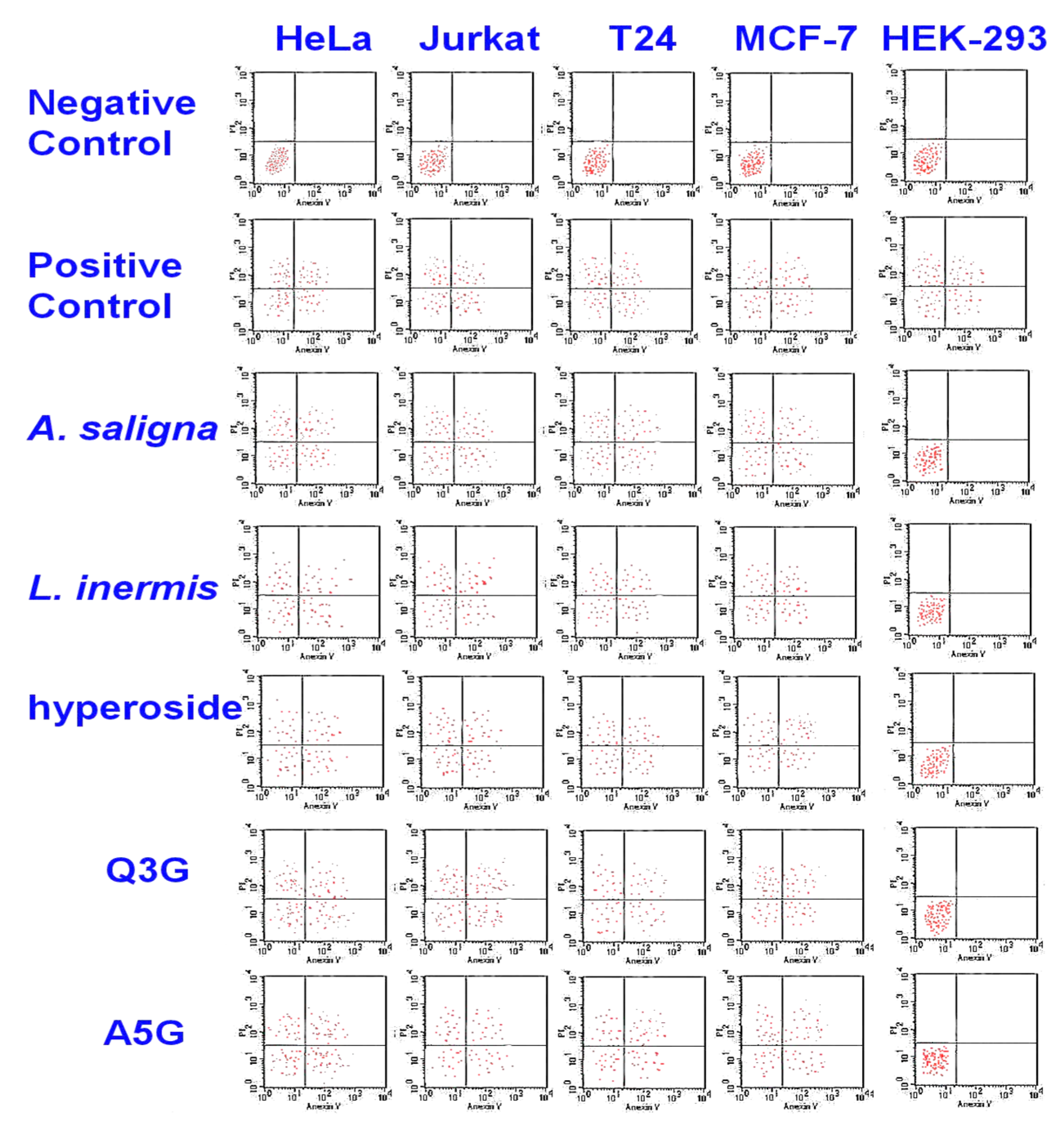
3.3. MTT Assay and Flow Cytometry
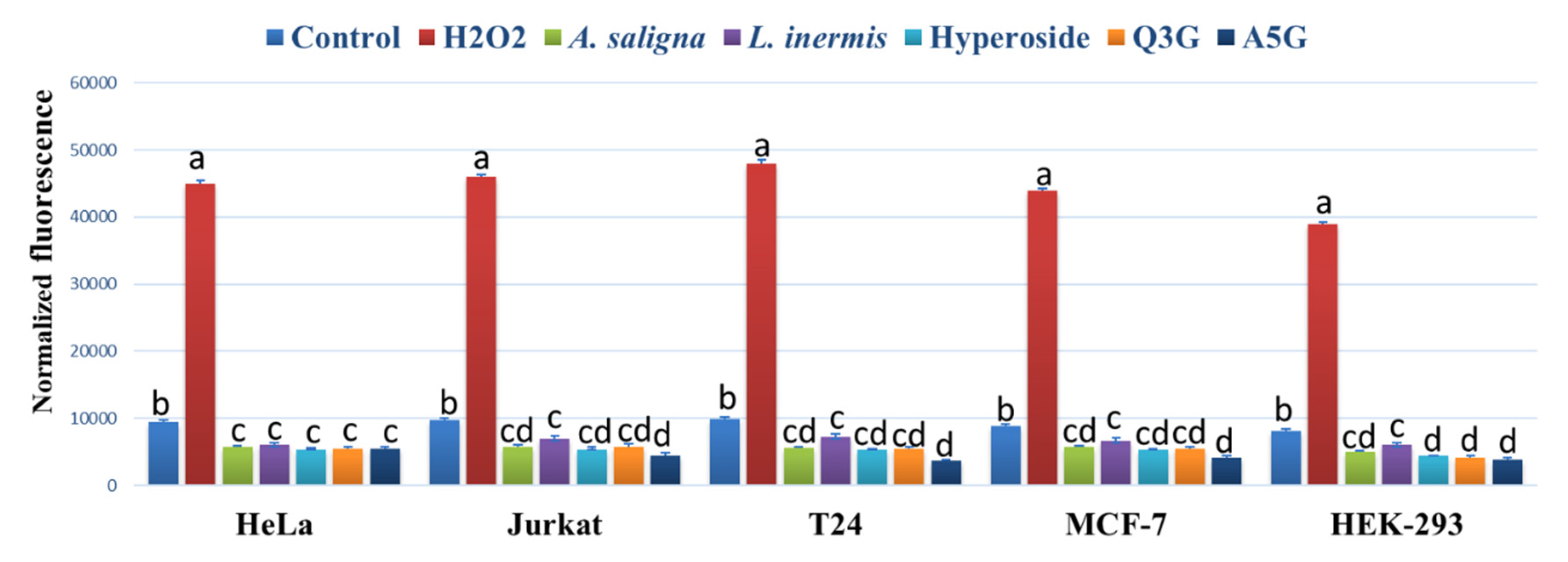
3.4. ROS Accumulation
3.5. Antibacterial Activities of A. saligna and L. inermis Leaf Extracts
3.6. Antifungal Effects of Leaf Extracts
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Organization, W.H. WHO Traditional Medicine Strategy: 2014–2023; WHO Press: Geneva, Switzerland, 2013; Available online: https://apps.who.int/iris/bitstream/handle/10665/92455/9786167697581-tha.pdf (accessed on 16 July 2020).
- Queen, B.; Tollefsbol, T. Polyphenols and Aging. Curr. Aging Sci. 2010, 3, 34–42. [Google Scholar] [CrossRef] [PubMed]
- Su, B.; Wang, X.; Nunomura, A.; Moreira, P.I.; Lee, H.G.; Perry, G.; Smith, M.A.; Zhu, X. Oxidative stress signaling in Alzheimer’s disease. Curr. Alzheimer. Res. 2008, 5, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Luo, Q.; Nie, R.; Yang, X.; Tang, Z.; Chen, H. Potential implications of polyphenols on aging considering oxidative stress, inflammation, autophagy, and gut microbiota. Crit. Rev. Food Sci. Nutr. 2020, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Elansary, H.O.; Szopa, A.; Kubica, P.; Al-Mana, F.A.; Mahmoud, E.A.; El-Abedin, T.K.A.Z.; Mattar, M.A.; Ekiert, H. Phenolic Compounds of Catalpa speciosa, Taxus cuspidata, and Magnolia acuminata have Antioxidant and Anticancer Activity. Molecules 2019, 24, 412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elansary, O.H.; Szopa, A.; Kubica, P.; Ekiert, H.; Mattar, M.A.; Al-Yafrasi, A.M.; El-Ansary, O.D.; Zin El-Abedin, K.T.; Yessoufou, K. Polyphenol Profile and Pharmaceutical Potential of Quercus spp. Bark Extracts. Plants 2019, 8, 486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleh, A.; ElFayoumi, H.M.; Youns, M.; Barakat, W. Rutin and orlistat produce antitumor effects via antioxidant and apoptotic actions. Naunyn. Schmiedebergs Arch. Pharmacol. 2019, 392, 165–175. [Google Scholar] [CrossRef] [PubMed]
- Yoo, T.-K.; Kim, J.-S.; Hyun, T.K. Polyphenolic Composition and Anti-Melanoma Activity of White Forsythia (Abeliophyllum distichum Nakai) Organ Extracts. Plants 2020, 9, 757. [Google Scholar] [CrossRef]
- Gutiérrez-Grijalva, E.P.; Picos-Salas, M.A.; Leyva-López, N.; Criollo-Mendoza, M.S.; Vazquez-Olivo, G.; Heredia, J.B. Flavonoids and Phenolic Acids from Oregano: Occurrence, Biological Activity and Health Benefits. Plants 2018, 7, 2. [Google Scholar] [CrossRef] [Green Version]
- Okla, K.M.; Alamri, A.S.; Salem, Z.M.M.; Ali, M.H.; Behiry, I.S.; Nasser, A.R.; Alaraidh, A.I.; Al-Ghtani, M.S.; Soufan, W. Yield, Phytochemical Constituents, and Antibacterial Activity of Essential Oils from the Leaves/Twigs, Branches, Branch Wood, and Branch Bark of Sour Orange (Citrus aurantium L.). Processes 2019, 7, 363. [Google Scholar] [CrossRef] [Green Version]
- Salem, M.Z.M.; Elansary, H.O.; Ali, H.M.; El-Settawy, A.A.; Elshikh, M.S.; Abdel-Salam, E.M.; Skalicka-Wozniak, K. Bioactivity of essential oils extracted from Cupressus macrocarpa branchlets and Corymbia citriodora leaves grown in Egypt. BMC Complement Altern. Med. 2018, 18, 23. [Google Scholar] [CrossRef]
- Khan, M.; Siddiqui, S.A. Concurrent chemoradiotherapy with or without induction chemotherapy for the management of cervical lymph node metastasis from unknown primary tumor. J. Cancer Res. Ther. 2018, 14, 1117–1120. [Google Scholar] [CrossRef] [PubMed]
- Elansary, H.; Szopa, A.; Kubica, P.; Ekiert, H.; El-Ansary, D.; Al-Mana, F.; Mahmoud, E. Polyphenol Content and Biological Activities of Ruta graveolens L. and Artemisia abrotanum L. in Northern Saudi Arabia. Processes 2020, 8, 531. [Google Scholar] [CrossRef]
- Elansary, H.O.; Szopa, A.; Klimek-Szczykutowicz, M.; Jafernik, K.; Ekiert, H.; Mahmoud, E.A.; Barakat, A.A.; El-Ansary, D.O. Mammillaria Species—Polyphenols Studies and Anti-Cancer, Anti-Oxidant, and Anti-Bacterial Activities. Molecules 2019, 25, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adiamo, O.Q.; Netzel, M.E.; Hoffman, L.C.; Sultanbawa, Y. Acacia seed proteins: Low or high quality? A comprehensive review. Compr. Rev. Food Sci. Food Saf. 2020, 19, 21–43. [Google Scholar] [CrossRef]
- El Sissi, H.I.; El Sherbeiny, A.E.A. The flavanoid components of the leaves of Acacia saligna. Qual. Plant. Mater. Veg. 1967, 14, 257–266. [Google Scholar] [CrossRef]
- El-Toumy, S.; Salib, J.; Mohamed, W.; Morsy, F. Phytochemical and antimicrobial studies on Acacia Saligna Leaves. Egypt. J. Chem. 2010, 53, 705. [Google Scholar]
- Al-Huqail, A.A.; Behiry, S.I.; Salem, M.Z.M.; Ali, H.M.; Siddiqui, M.H.; Salem, A.Z.M. Antifungal, Antibacterial, and Antioxidant Activities of Acacia Saligna (Labill.) H. L. Wendl. Flower Extract: HPLC Analysis of Phenolic and Flavonoid Compounds. Molecules 2019, 24, 700. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Farid, I.B.; Sheded, M.G.; Mohamed, E.A. Metabolomic profiling and antioxidant activity of some Acacia species. Saudi. J. Biol. Sci. 2014, 21, 400–408. [Google Scholar] [CrossRef] [Green Version]
- Alajmi, M.F.; Alam, P.; Alqasoumi, S.I.; Ali Siddiqui, N.; Basudan, O.A.; Hussain, A.; Mabood Husain, F.; Ali Khan, A. Comparative anticancer and antimicrobial activity of aerial parts of Acacia salicina, Acacia laeta, Acacia hamulosa and Acacia tortilis grown in Saudi Arabia. Saudi Pharm. J. 2017, 25, 1248–1252. [Google Scholar] [CrossRef]
- Abd-ElGawad, A.; El-Amier, Y. Allelopathy and Potential Impact of Invasive Acacia saligna (Labill.) Wendl. on Plant Diversity in the Nile Delta Coast of Egypt. Int. J. Environ. Res. 2015, 9, 923–932. [Google Scholar]
- Badoni Semwal, R.; Semwal, D.K.; Combrinck, S.; Cartwright-Jones, C.; Viljoen, A. Lawsonia inermis L. (henna): Ethnobotanical, phytochemical and pharmacological aspects. J. Ethnopharmacol. 2014, 155, 80–103. [Google Scholar] [CrossRef] [PubMed]
- Hsouna, A.; Trigui, M.; Culioli, G.; Blache, Y.; Jaoua, S. Antioxidant constituents from Lawsonia inermis leaves: Isolation, structure elucidation and antioxidative capacity. Food Chem. 2011, 125, 193–200. [Google Scholar] [CrossRef]
- Wiem, A.; Smail, A.; Mnif, W.; Faleiro, M.L.; Miguel, M. Antioxidant, anti-inflammatory and anti-acetylcholinesterase activities of leaf, flower and seed aqueous extracts of Lawsonia inermis from Tunisia. Int. J. Pharm. Pharm. Sci. 2014, 6, 445–452. [Google Scholar]
- Pasandi Pour, A.; Farahbakhsh, H. Lawsonia inermis L. leaves aqueous extract as a natural antioxidant and antibacterial product. Nat. Prod. Res. 2019, 1–5. [Google Scholar] [CrossRef]
- Guha, G.; Rajkumar, V.; Kumar, R.A.; Mathew, L. Antioxidant Activity of Lawsonia inermis Extracts Inhibits Chromium(VI)-Induced Cellular and DNA Toxicity. Evid.-Based Complement. Altern. Med. eCAM 2011, 2011, 576456. [Google Scholar] [CrossRef] [Green Version]
- Elansary, H.O.; Szopa, A.; Kubica, P.; El-Ansary, D.O.; Ekiert, H.; Al-Mana, F.A. Malus baccata var. gracilis and Malus toringoides Bark Polyphenol Studies and Antioxidant, Antimicrobial and Anticancer Activities. Processes 2020, 8, 283. [Google Scholar] [CrossRef] [Green Version]
- Ellnain-Wojtaszek, M.; Zgórka, G. High-performance liquid chromatography and thin-layer chromatography of phenolic acids from Ginkgo biloba L. leaves collected within vegetative period. J. Liq. Chromatogr. R. T 1999, 22, 1457–1471. [Google Scholar] [CrossRef]
- Sułkowska-Ziaja, K.; Maślanka, A.; Szewczyk, A.; Muszyńska, B. Physiologically Active Compounds in Four Species of Phellinus. Nat. Prod. Commun. 2017, 12, 363–366. [Google Scholar] [CrossRef] [Green Version]
- Elansary, H.O.; Yessoufou, K.; Abdel-Hamid, A.M.E.; El-Esawi, M.A.; Ali, H.M.; Elshikh, M.S. Seaweed Extracts Enhance Salam Turfgrass Performance during Prolonged Irrigation Intervals and Saline Shock. Front. Plant Sci. 2017, 8, 10–3389. [Google Scholar] [CrossRef] [Green Version]
- Ferreira, J.P.A.; Miranda, I.; Sousa, V.B.; Pereira, H. Chemical composition of barks from Quercus faginea trees and characterization of their lipophilic and polar extracts. PLoS ONE 2018, 13, e0197135. [Google Scholar] [CrossRef]
- Elansary, H.O.; Mahmoud, E.A. Egyptian herbal tea infusions’ antioxidants and their antiproliferative and cytotoxic activities against cancer cells. Nat. Prod. Res. 2015, 29, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Elansary, H.O.; Szopa, A.; Klimek-Szczykutowicz, M.; Ekiert, H.; Barakat, A.A.; Al-Mana, F.A. Antiproliferative, Antimicrobial, and Antifungal Activities of Polyphenol Extracts from Ferocactus Species. Processes 2020, 8, 138. [Google Scholar] [CrossRef] [Green Version]
- Elansary, H.O.; Mahmoud, E.A. Basil cultivar identification using chemotyping still favored over genotyping using core barcodes and possible resources of antioxidants. J. Essent. Oil Res. 2015, 27, 82–87. [Google Scholar] [CrossRef]
- Nizioł-Łukaszewska, Z.; Furman-Toczek, D.; Zagórska-Dziok, M. Antioxidant activity and cytotoxicity of Jerusalem artichoke tubers and leaves extract on HaCaT and BJ fibroblast cells. Lipids Health Dis. 2018, 17, 280. [Google Scholar] [CrossRef] [Green Version]
- Yessoufou, K.; Elansary, H.O.; Mahmoud, E.A.; Skalicka-Wozniak, K. Antifungal, antibacterial and anticancer activities of Ficus drupacea L. stem bark extract and biologically active isolated compounds. Ind. Crop Prod. 2015, 74, 752–758. [Google Scholar] [CrossRef]
- Elansary, H.O.; Abdelgaleil, S.A.M.; Mahmoud, E.A.; Yessoufou, K.; Elhindi, K.; El-Hendawy, S. Effective antioxidant, antimicrobial and anticancer activities of essential oils of horticultural aromatic crops in northern Egypt. BMC Complement. Altern. Med. 2018, 18, 214. [Google Scholar] [CrossRef]
- Figueroa, D.; Asaduzzaman, M.; Young, F. Real time monitoring and quantification of reactive oxygen species in breast cancer cell line MCF-7 by 2′,7′–dichlorofluorescin diacetate (DCFDA) assay. J. Pharmacol. Toxicol. Methods 2018, 94, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Elansary, H.O.; Szopa, A.; Kubica, P.; Ekiert, H.; Ali, H.M.; Elshikh, M.S.; Abdel-Salam, E.M.; El-Esawi, M.; El-Ansary, D.O. Bioactivities of Traditional Medicinal Plants in Alexandria. Evid. Based Complement. Altern. Med. 2018, 2018, 1463579. [Google Scholar] [CrossRef] [Green Version]
- Elansary, H.O.; Yessoufou, K.; Shokralla, S.; Mahmoud, E.A.; Skaicka-Wozniak, K. Enhancing mint and basil oil composition and antibacterial activity using seaweed extracts. Ind. Crop Prod. 2016, 92, 50–56. [Google Scholar] [CrossRef]
- Abd El-Kareem, M.S.M.; Mohamed, A.R.; Elansary, H.O.; Al-Mana, F.A. Mass Spectral Fragmentation of Pelargonium graveolens Essential Oil Using GC–MS Semi-Empirical Calculations and Biological Potential. Processes 2020, 8, 128. [Google Scholar] [CrossRef] [Green Version]
- Gedara, S.; Galala, A. New cytotoxic spirostane saponin and biflavonoid glycoside from the leaves of Acacia saligna (Labill.) H.L. Wendl. Nat. Prod. Res. 2013, 28, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Dube, J.S.; Reed, J.D.; Ndlovu, L.R. Proanthocyanidins and other phenolics in Acacia leaves of Southern Africa. Anim. Feed Sci. Technol. 2001, 91, 59–67. [Google Scholar] [CrossRef]
- Bishai, B.; Badria, F.; Maatooq, G.; Amer, M. Antioxidant and Immunomodulatory Constituents of Henna Leaves. Z. Nat. C J. Biosci. 2004, 59, 468–476. [Google Scholar] [CrossRef]
- Mahmoud, Z.F.; Salam, N.A.; Khafagy, S.M. Constituents of henna leaves (Lawsonia inermis L.) growing in Egypt. Fitoterapia 1980, 51, 153–155. [Google Scholar]
- Ostovari, A.; Hoseinieh, S.M.; Peikari, M.; Shadizadeh, S.R.; Hashemi, S.J. Corrosion inhibition of mild steel in 1M HCl solution by henna extract: A comparative study of the inhibition by henna and its constituents (Lawsone, Gallic acid, α-d-Glucose and Tannic acid). Corros. Sci. 2009, 51, 1935–1949. [Google Scholar] [CrossRef]
- Ran, J.; Sun, H.; Xu, Y.; Wang, T.; Zhao, R. Comparison of Antioxidant Activities and HPLC Analysis of Polyphenol from Different Apple Varieties. Int. J. Food Prop. 2016, 19. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Swarts, S.G.; Yin, L.; Liu, C.; Tian, Y.; Cao, Y.; Swarts, M.; Yang, S.; Zhang, S.B.; Zhang, K.; et al. Antioxidant properties of quercetin. Adv. Exp. Med. Biol. 2011, 701, 283–289. [Google Scholar] [CrossRef]
- Murakami, Y.; Kawata, A.; Ito, S.; Katayama, T.; Fujisawa, S. Radical-Scavenging and Anti-inflammatory Activity of Quercetin and Related Compounds and Their Combinations Against RAW264.7 Cells Stimulated with Porphyromonas gingivalis Fimbriae. Relationships between Anti-inflammatory Activity and Quantum Chemical Parameters. In Vivo 2015, 29, 701–710. [Google Scholar]
- Velderrain-Rodríguez, G.R.; Torres-Moreno, H.; Villegas-Ochoa, M.A.; Ayala-Zavala, J.F.; Robles-Zepeda, R.E.; Wall-Medrano, A.; González-Aguilar, G.A. Gallic Acid Content and an Antioxidant Mechanism Are Responsible for the Antiproliferative Activity of ‘Ataulfo’ Mango Peel on LS180 Cells. Molecules 2018, 23, 695. [Google Scholar] [CrossRef] [Green Version]
- Kilic, I.; Yesiloglu, Y. Spectroscopic studies on the antioxidant activity of p-coumaric acid. Spectrochim. Acta A Mol. Biomol. Spectrosc. 2013, 115, 719–724. [Google Scholar] [CrossRef]
- Buravlev, E.V.; Dvornikova, I.A.; Schevchenko, O.G.; Kutchin, A.V. Synthesis and Antioxidant Ability of Novel Derivatives Based on para-Coumaric Acid Containing Isobornyl Groups. Chem. Biodivers. 2019, 16, e1900362. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.; Gupta, S. Apigenin: A promising molecule for cancer prevention. Pharm. Res. 2010, 27, 962–978. [Google Scholar] [CrossRef] [PubMed]
- Ganeshpurkar, A.; Saluja, A.K. The Pharmacological Potential of Rutin. Saudi Pharm. J. 2017, 25, 149–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Z.; Zhao, P.; Xu, H. Hyperoside exhibits anticancer activity in non-small cell lung cancer cells with T790M mutations by upregulating FoxO1 via CCAT1. Oncol. Rep. 2020, 43, 617–624. [Google Scholar] [CrossRef]
- Zhang, Y.; Dong, H.; Zhang, J.; Zhang, L. Inhibitory effect of hyperoside isolated from Zanthoxylum bungeanum leaves on SW620 human colorectal cancer cells via induction of the p53 signaling pathway and apoptosis. Mol. Med. Rep. 2017, 16, 1125–1132. [Google Scholar] [CrossRef] [Green Version]
- Ghițu, A.; Schwiebs, A.; Radeke, H.H.; Avram, S.; Zupko, I.; Bor, A.; Pavel, I.Z.; Dehelean, C.A.; Oprean, C.; Bojin, F.; et al. A Comprehensive Assessment of Apigenin as an Antiproliferative, Proapoptotic, Antiangiogenic and Immunomodulatory Phytocompound. Nutrients 2019, 11, 858. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Heber, D.; Henning, S.M. Quercetin increased the antiproliferative activity of green tea polyphenol (-)-epigallocatechin gallate in prostate cancer cells. Nutr. Cancer 2012, 64, 580–587. [Google Scholar] [CrossRef]
- Zhao, M.H.; Yuan, L.; Meng, L.Y.; Qiu, J.L.; Wang, C.B. Quercetin-loaded mixed micelles exhibit enhanced cytotoxic efficacy in non-small cell lung cancer in vitro. Exp. Ther. Med. 2017, 14, 5503–5508. [Google Scholar] [CrossRef] [Green Version]
- Karpiński, T.; Adamczak, A.; Ożarowski, M. Antibacterial activity of apigenin, luteolin, and their C-glucosides. In Proceedings of the 5th International Electronic Conference on Medicinal Chemistry, Poland, 1–30 November 2019. [Google Scholar] [CrossRef] [Green Version]
- Bag, A.; Chattopadhyay, R.R. Synergistic antibacterial and antibiofilm efficacy of nisin in combination with p-coumaric acid against food-borne bacteria Bacillus cereus and Salmonella typhimurium. Lett. Appl. Microbiol. 2017, 65, 366–372. [Google Scholar] [CrossRef]
- Hirai, I.; Okuno, M.; Katsuma, R.; Arita, N.; Tachibana, M.; Yamamoto, Y. Characterisation of anti-Staphylococcus aureus activity of quercetin. Int. J. Food Sci. Technol. 2010, 45, 1250–1254. [Google Scholar] [CrossRef]
- Jaisinghani, R. Antibacterial properties of quercetin. Microbiol. Res. 2017, 8. [Google Scholar] [CrossRef] [Green Version]
- Sze Kwan, L.; Tzi Bun, N. Acafusin, a Dimeric Antifungal Protein from Acacia confusa Seeds. Protein Pept. Lett. 2010, 17, 817–822. [Google Scholar] [CrossRef] [PubMed]
- Smiljkovic, M.; Stanisavljevic, D.; Stojkovic, D.; Petrovic, I.; Marjanovic Vicentic, J.; Popovic, J.; Golic Grdadolnik, S.; Markovic, D.; Sankovic-Babice, S.; Glamoclija, J.; et al. Apigenin-7-O-glucoside versus apigenin: Insight into the modes of anticandidal and cytotoxic actions. EXCLI J. 2017, 16, 795–807. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Zhang, Z.; Cain, A.; Wang, B.; Long, M.; Taylor, J. Antifungal Activity of Camptothecin, Trifolin, and Hyperoside Isolated from Camptotheca acuminata. J. Agric. Food Chem. 2005, 53, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Morales, J.; Mendoza, L.; Cotoras, M. Alteration of oxidative phosphorylation as a possible mechanism of the antifungal action of p-coumaric acid against Botrytis cinerea. J. Appl. Microbiol. 2017, 123, 969–976. [Google Scholar] [CrossRef]
- Rocha, M.F.G.; Sales, J.A.; da Rocha, M.G.; Galdino, L.M.; de Aguiar, L.; Pereira-Neto, W.d.A.; de Aguiar Cordeiro, R.; Castelo-Branco, D.d.S.C.M.; Sidrim, J.J.C.; Brilhante, R.S.N. Antifungal effects of the flavonoids kaempferol and quercetin: A possible alternative for the control of fungal biofilms. Biofouling 2019, 35, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Yessoufou, K.; Daru, B.H.; Tafirei, R.; Elansary, H.O.; Rampedi, I. Integrating biogeography, threat and evolutionary data to explore extinction crisis in the taxonomic group of cycads. Ecol. Evol. 2017, 7, 2735–2746. [Google Scholar] [CrossRef] [Green Version]
- Salem, M.Z.M.; El-Hefny, M.; Nasser, R.A.; Ali, H.M.; El-Shanhorey, N.A.; Elansary, H.O. Medicinal and biological values of Callistemon viminalis extracts: History, current situation and prospects. Asian Pac. J. Trop. Med. 2017, 10, 229–237. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Acacia saligna | Lawsonia inermis | |
|---|---|---|---|
| Phenolic acids | Gallic acid | 136.2 ± 11.1 | 81.0 ± 13.2 |
| p-Coumaric acid | 34.8 ± 2.9 | nd * | |
| Flavonoids | Hyperoside | 632.5 ± 45.5 | nd |
| Isoquercetin | 73.2 ± 9.2 | nd | |
| Quercetin | 6.1 ± 0.5 | nd | |
| Quercetin 3-glucuronide | 125.5 ± 2.5 | nd | |
| Rutoside | 1533.0 ± 55.7 | nd | |
| Apigetrin | nd | 1180.9 ± 153.3 | |
| Apigenin 5-glucoside | nd | 596.3 ± 49.1 | |
| B. cereus MIC MBC | P. aeruginosa MIC MBC | L. monocytogenes MIC MBC | E. coli MIC MBC | M. flavus MIC MBC | S. aureus MIC MBC | |
|---|---|---|---|---|---|---|
| A. saligna | 0.35 ± 0.01 | 0.37 ± 0.02 | 0.47 ± 0.03 | 0.31 ± 0.03 | 0.41 ± 0.02 | 0.30 ± 0.05 |
| 0.73 ± 0.03 | 0.79 ± 0.03 | 0.99 ± 0.05 | 0.72 ± 0.01 | 0.85 ± 0.03 | 0.73 ± 0.03 | |
| L. inermis | 0.43 ± 0.03 | 0.41 ± 0.03 | 0.42 ± 0.03 | 0.34 ± 0.02 | 0.52 ± 0.05 | 0.41 ± 0.02 |
| 0.86 ± 0.04 | 0.96 ± 0.05 | 0.93 ± 0.02 | 0.75 ± 0.03 | 1.53 ± 0.12 | 0.93 ± 0.03 | |
| Rutoside | 0.11 ± 0.01 | 0.07 ± 0.01 | 0.11 ± 0.01 | 0.12 ± 0.01 | 0.12 ± 0.01 | 0.11 ± 0.01 |
| 0.22 ± 0.01 | 0.12 ± 0.01 | 0.21 ± 0.02 | 0.23 ± 0.01 | 0.23 ± 0.01 | 0.25 ± 0.03 | |
| Hyperoside | 23.3 ± 2.4 | 27.2 ± 3.53 | 34.3 ± 4.21 | 31.2 ± 1.43 | 22.42 ± 1.55 | 19.54 ± 2.41 |
| >500 | >500 | >500 | >500 | >500 | >500 | |
| Apigenin 5-glucoside | 0.13 ± 0.01 | 0.12 ± 0.01 | 0.10 ± 0.01 | 0.09 ± 0.01 | 0.10 ± 0.01 | 0.13 ± 0.02 |
| 0.33 ± 0.03 | 0.31 ± 0.03 | 0.28 ± 0.02 | 0.27 ± 0.03 | 0.30 ± 0.03 | 0.33 ± 0.03 | |
| p-Coumaric acid | 0.12 ± 0.01 | 0.06 ± 0.01 | 0.26 ± 0.02 | 0.12 ± 0.01 | 0.16 ± 0.02 | 0.23± 0.02 |
| 0.31 ± 0.03 | 0.22 ± 0.02 | 0.58 ± 0.03 | 0.25 ± 0.02 | 0.38 ± 0.03 | 0.47 ± 0.03 | |
| Quercetin | 30.6 ± 3.1 | 31.8 ± 2.11 | 43.7 ± 3.51 | 38.6 ± 3.14 | 28.65 ± 2.11 | 21.53 ± 1.53 |
| >500 | >500 | >500 | >500 | >500 | >500 | |
| Quercetin 3-glucuronide | 35.2 ± 3.1 | 25.3 ± 1.54 | 32.1 ± 3.78 | 31.8 ± 3.32 | 21.65 ± 2.11 | 17.1 ± 1.53 |
| >500 | 315.14 ± 18.42 | >500 | >500 | 411.17 ± 22.67 | 427 ± 27.23 | |
| Streptomycin | 0.07 ± 0.01 | 0.11 ± 0.01 | 0.12 ± 0.01 | 0.11 ± 0.01 | 0.10 ± 0.01 | 0.15 ± 0.01 |
| 0.16 ± 0.02 | 0.21 ± 0.02 | 0.21 ± 0.02 | 0.20 ± 0.01 | 0.21 ± 0.02 | 0.31 ± 0.02 |
| Aspergillus flavus MIC MFC | Aspergillus ochraceus MIC MFC | Aspergillus niger MIC MFC | Candida albicans MIC MFC | Penicillium funiculosum MIC MFC | Penicillium ochrochloron MIC MFC | |
|---|---|---|---|---|---|---|
| A. saligna | 0.30 ± 0.02 | 0.38 ± 0.02 | 0.48 ± 0.03 | 0.58 ± 0.03 | 0.43 ± 0.03 | 0.44 ± 0.05 |
| 0.91 ± 0.05 | 0.95 ± 0.3 | 1.02 ± 0.05 | 1.42 ± 0.18 | 1.01 ± 0.06 | 1.31 ± 0.12 | |
| L. inermis | 0.45 ± 0.05 | 0.47 ± 0.03 | 0.64 ± 0.05 | 0.88 ± 0.04 | 0.75 ± 0.02 | 0.79 ± 0.03 |
| 0.98 ± 0.06 | 1.07 ± 0.05 | 1.45 ± 0.17 | 1.86 ± 0.13 | 1.53 ± 0.05 | 1.64 ± 0.12 | |
| Rutoside | 0.21 ± 0.02 | 0.18 ± 0.01 | 0.28 ± 0.03 | 0.25 ± 0.03 | 0.30 ± 0.02 | 0.23 ± 0.03 |
| 0.45 ± 0.03 | 0.55 ± 0.03 | 0.62 ± 0.03 | 0.51 ± 0.03 | 0.71 ± 0.03 | 0.43 ± 0.03 | |
| Hyperoside | 0.10 ± 0.02 | 0.13 ± 0.03 | 0.15 ± 0.03 | 0.21 ± 0.03 | 0.25 ± 0.03 | 0.31 ± 0.03 |
| 0.46 ± 0.03 | 0.50 ± 0.05 | 0.52 ± 0.04 | 0.93 ± 0.07 | 1.03 ± 0.11 | 1.19 ± 0.10 | |
| Apigenin 5-glucoside | 0.11 ± 0.03 | 0.17 ± 0.02 | 0.23 ± 0.03 | 0.03 ± 0.01 | 0.75 ± 0.05 | 0.59 ± 0.05 |
| 0.84 ± 0.04 | 1.03 ± 0.08 | 1.11 ± 0.15 | 0.08 ± 0.01 | 1.33 ± 0.14 | 1.26 ± 0.13 | |
| Quercetin | 0.31 ± 0.03 | 0.20 ± 0.02 | 0.21 ± 0.01 | 0.06 ± 0.01 | 0.24 ± 0.03 | 0.29 ± 0.03 |
| 0.63 ± 0.05 | 0.75 ± 0.03 | 0.75 ± 0.02 | 0.33 ± 0.03 | 0.70 ± 0.04 | 0.63 ± 0.03 | |
| Quercetin 3-glucuronide | 0.26 ± 0.03 | 0.17 ± 0.03 | 0.18 ± 0.02 | 0.06 ± 0.01 | 0.21 ± 0.01 | 0.26 ± 0.02 |
| 0.52 ± 0.03 | 0.61 ± 0.05 | 0.62 ± 0.05 | 0.27 ± 0.03 | 0.60 ± 0.05 | 0.54 ± 0.05 | |
| p-Coumaric acid | 0.22 ± 0.03 | 0.23 ± 0.02 | 0.21 ± 0.01 | 0.32 ± 0.03 | 0.22 ± 0.01 | 0.20 ± 0.02 |
| 0.43 ± 0.01 | 0.45 ± 0.05 | 0.41 ± 0.03 | 0.60 ± 0.01 | 0.59 ± 0.05 | 0.40 ± 0.03 | |
| KTZ (Ketoconazole) | 0.20 ± 0.01 | 0.23 ± 0.02 | 0.10 ± 0.01 | 0.22 ± 0.01 | 2.05 ± 0.13 | 0.21 ± 0.01 |
| 0.41 ± 0.03 | 0.46 ± 0.03 | 0.21 ± 0.03 | 0.43 ± 0.02 | 3.51 ± 0.11 | 0.43 ± 0.05 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elansary, H.O.; Szopa, A.; Kubica, P.; Ekiert, H.; A. Al-Mana, F.; Al-Yafrsi, M.A. Antioxidant and Biological Activities of Acacia saligna and Lawsonia inermis Natural Populations. Plants 2020, 9, 908. https://doi.org/10.3390/plants9070908
Elansary HO, Szopa A, Kubica P, Ekiert H, A. Al-Mana F, Al-Yafrsi MA. Antioxidant and Biological Activities of Acacia saligna and Lawsonia inermis Natural Populations. Plants. 2020; 9(7):908. https://doi.org/10.3390/plants9070908
Chicago/Turabian StyleElansary, Hosam O., Agnieszka Szopa, Paweł Kubica, Halina Ekiert, Fahed A. Al-Mana, and Mohammed A. Al-Yafrsi. 2020. "Antioxidant and Biological Activities of Acacia saligna and Lawsonia inermis Natural Populations" Plants 9, no. 7: 908. https://doi.org/10.3390/plants9070908