Figure 1.
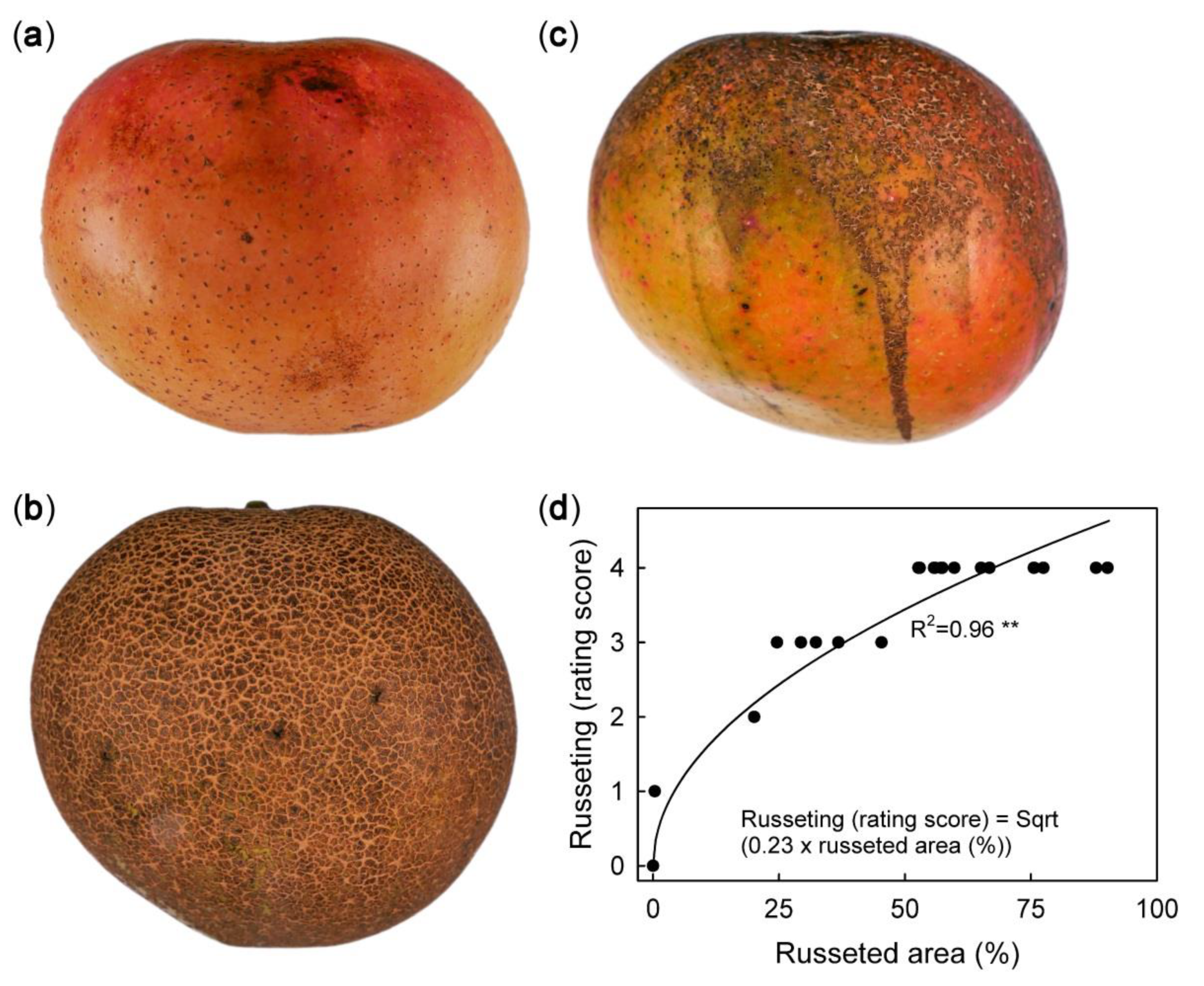
Macroscope view of mature ‘Apple’ mango without (a, score 0), moderate (b, score 2), and extreme (c, score 4) russet symptoms. (d): Plot of russeting (rating score) against percentage area affected by russet (image analysis). Each fruit was rated visually prior to image analysis. The number of observations was 18.
Figure 1.
Macroscope view of mature ‘Apple’ mango without (a, score 0), moderate (b, score 2), and extreme (c, score 4) russet symptoms. (d): Plot of russeting (rating score) against percentage area affected by russet (image analysis). Each fruit was rated visually prior to image analysis. The number of observations was 18.
Figure 2.
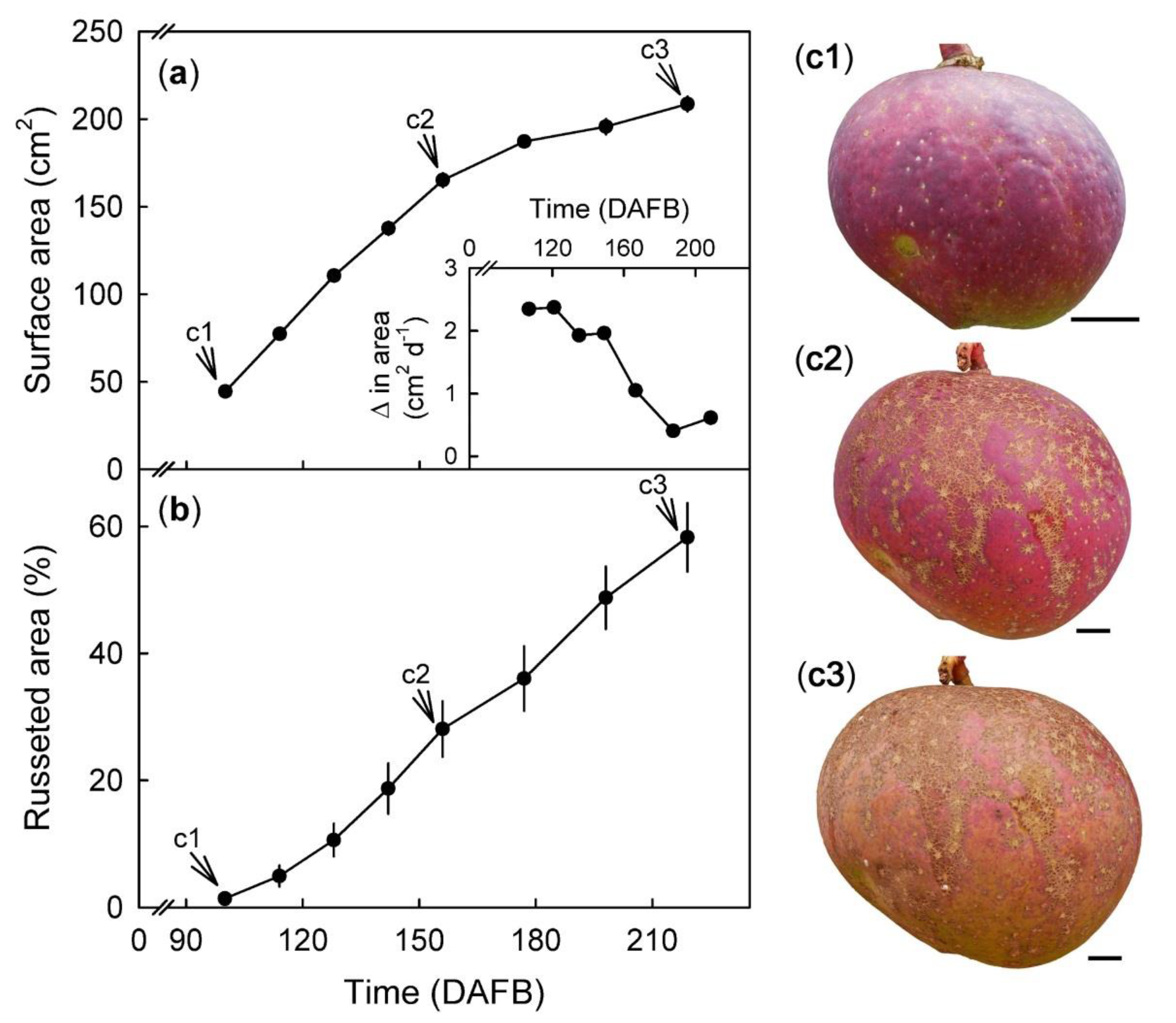
Change in fruit surface area (cm2) and rate of surface expansion (a and a inset) with time (days after full bloom, DAFB). Percent of skin with russet in developing fruit (b) calculated from a defined area of the fruit cheek. The same fruit was photographed at 100, 156, and 216 DAFB (see arrows). Pictorial representation of russet progression in a developing ‘Apple’ mango fruit (c1–c3). Scale bar is 10 mm. Data represent means ± SE of 19 replicates.
Figure 2.
Change in fruit surface area (cm2) and rate of surface expansion (a and a inset) with time (days after full bloom, DAFB). Percent of skin with russet in developing fruit (b) calculated from a defined area of the fruit cheek. The same fruit was photographed at 100, 156, and 216 DAFB (see arrows). Pictorial representation of russet progression in a developing ‘Apple’ mango fruit (c1–c3). Scale bar is 10 mm. Data represent means ± SE of 19 replicates.
Figure 3.
Microscopic view of ‘Apple’ mango skin infiltrated with acridine orange dye when viewed with a binocular microscope under natural (a,c,e,g,i,k,m) or fluorescent light (b,d,f,h,j,l,n). The scale bars refer to the corresponding pairs of images.
Figure 3.
Microscopic view of ‘Apple’ mango skin infiltrated with acridine orange dye when viewed with a binocular microscope under natural (a,c,e,g,i,k,m) or fluorescent light (b,d,f,h,j,l,n). The scale bars refer to the corresponding pairs of images.
Figure 4.
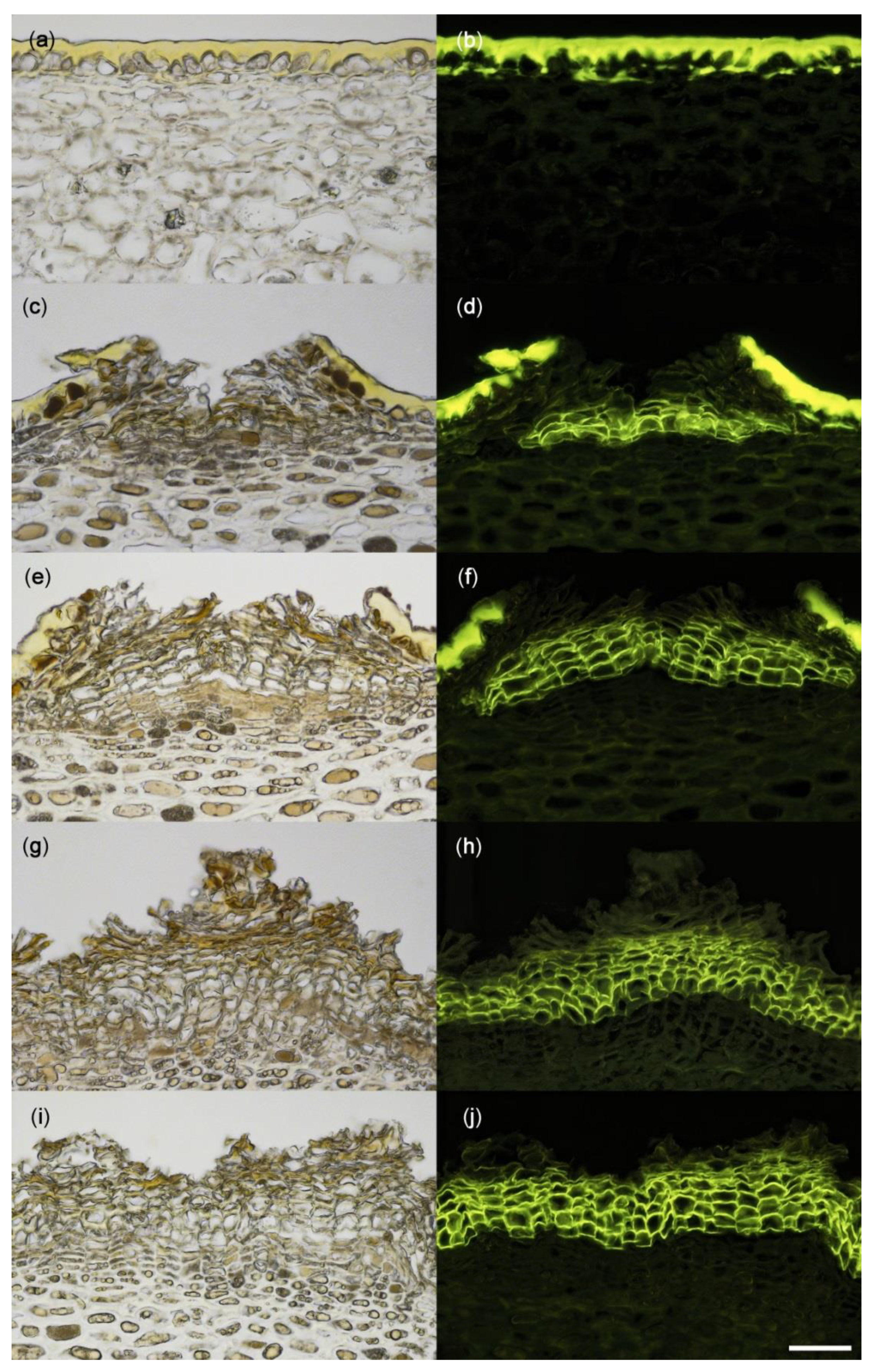
Cross-sectional microscope images of a non-russeted (a,b) and russeted (c–j) skin of ‘Apple’ mango viewed under incident white (a,c,e,g,i) or fluorescent light (b,d,f,h,j) following staining with fluorol yellow dye. Scale bar is 50 µm.
Figure 4.
Cross-sectional microscope images of a non-russeted (a,b) and russeted (c–j) skin of ‘Apple’ mango viewed under incident white (a,c,e,g,i) or fluorescent light (b,d,f,h,j) following staining with fluorol yellow dye. Scale bar is 50 µm.
Figure 5.
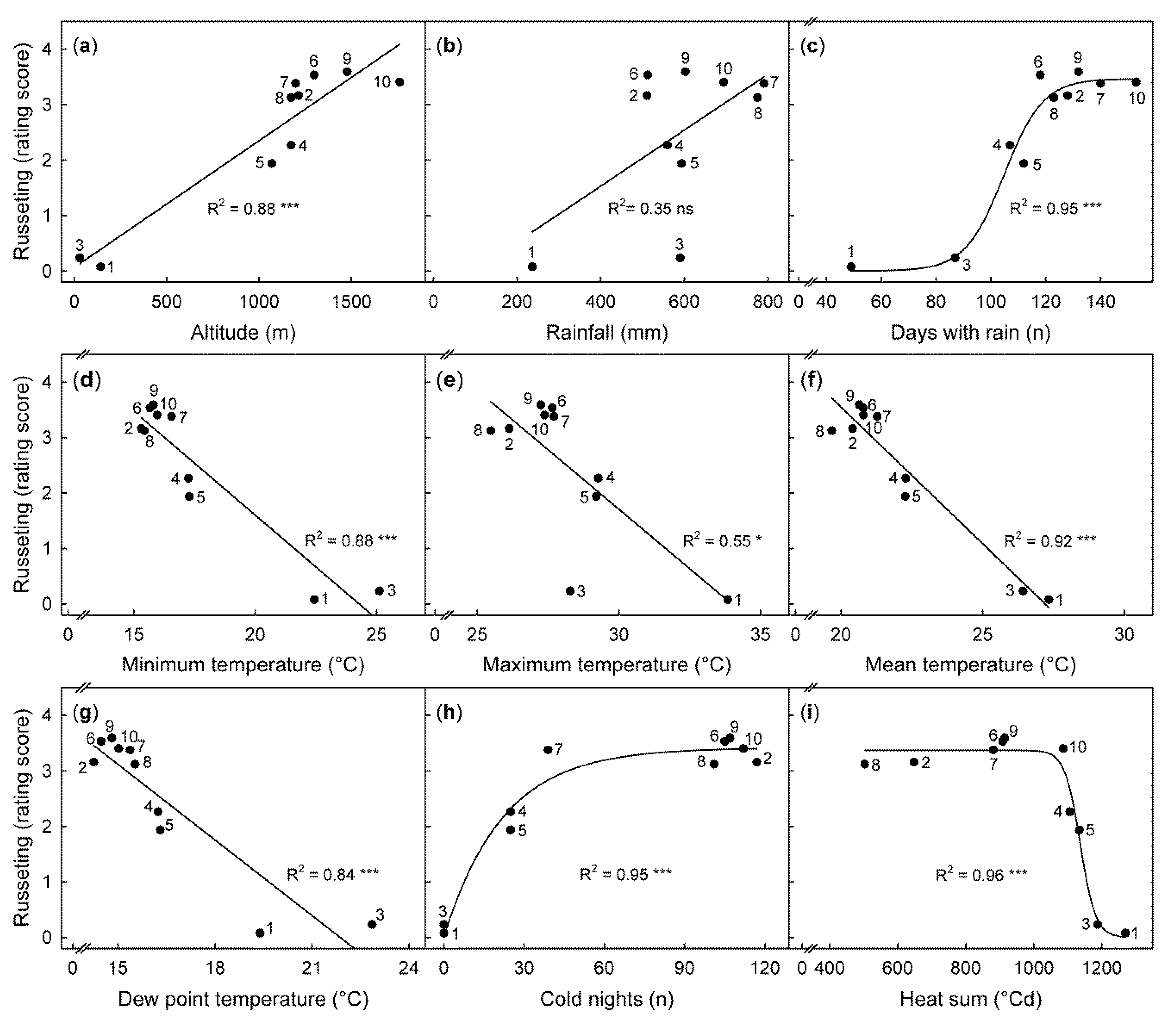
Relationship between climatic variables and average russeting (rating score) determined for the cumulative period of fruit maturity at ten locations in Kenya. The ten locations were: Garissa (1), Chepsigot (2), Malindi (3), Mumbuni (4), Yeemulwa (5), Kaiti (6), Kasafari (7), Kambirwa (8), Thika (9), and Machakos (10) situated at different altitudes. (a) The climatic variables include: rainfall amount (b), days with rainfall (c), relative humidity (d), minimum, maximum, and mean daily temperatures (e, f, and g, respectively). Cold nights (h) correspond to the number of days when the minimum temperature fell below the base temperature of 16 °C. Heat sum (i) was calculated based on a base temperature of 16 °C. Russeting was quantified using a five-score rating scheme: score 0: 0% of the fruit surface area russeted; score 1: 1–10% russeted area; score 2: 11–25% russeted area; score 3: 26–50% russeted area; and score 4: 51–100% russeted area. Data points represent means of 210 fruit per site.
Figure 5.
Relationship between climatic variables and average russeting (rating score) determined for the cumulative period of fruit maturity at ten locations in Kenya. The ten locations were: Garissa (1), Chepsigot (2), Malindi (3), Mumbuni (4), Yeemulwa (5), Kaiti (6), Kasafari (7), Kambirwa (8), Thika (9), and Machakos (10) situated at different altitudes. (a) The climatic variables include: rainfall amount (b), days with rainfall (c), relative humidity (d), minimum, maximum, and mean daily temperatures (e, f, and g, respectively). Cold nights (h) correspond to the number of days when the minimum temperature fell below the base temperature of 16 °C. Heat sum (i) was calculated based on a base temperature of 16 °C. Russeting was quantified using a five-score rating scheme: score 0: 0% of the fruit surface area russeted; score 1: 1–10% russeted area; score 2: 11–25% russeted area; score 3: 26–50% russeted area; and score 4: 51–100% russeted area. Data points represent means of 210 fruit per site.
![Plants 09 00898 g005]()
Figure 6.
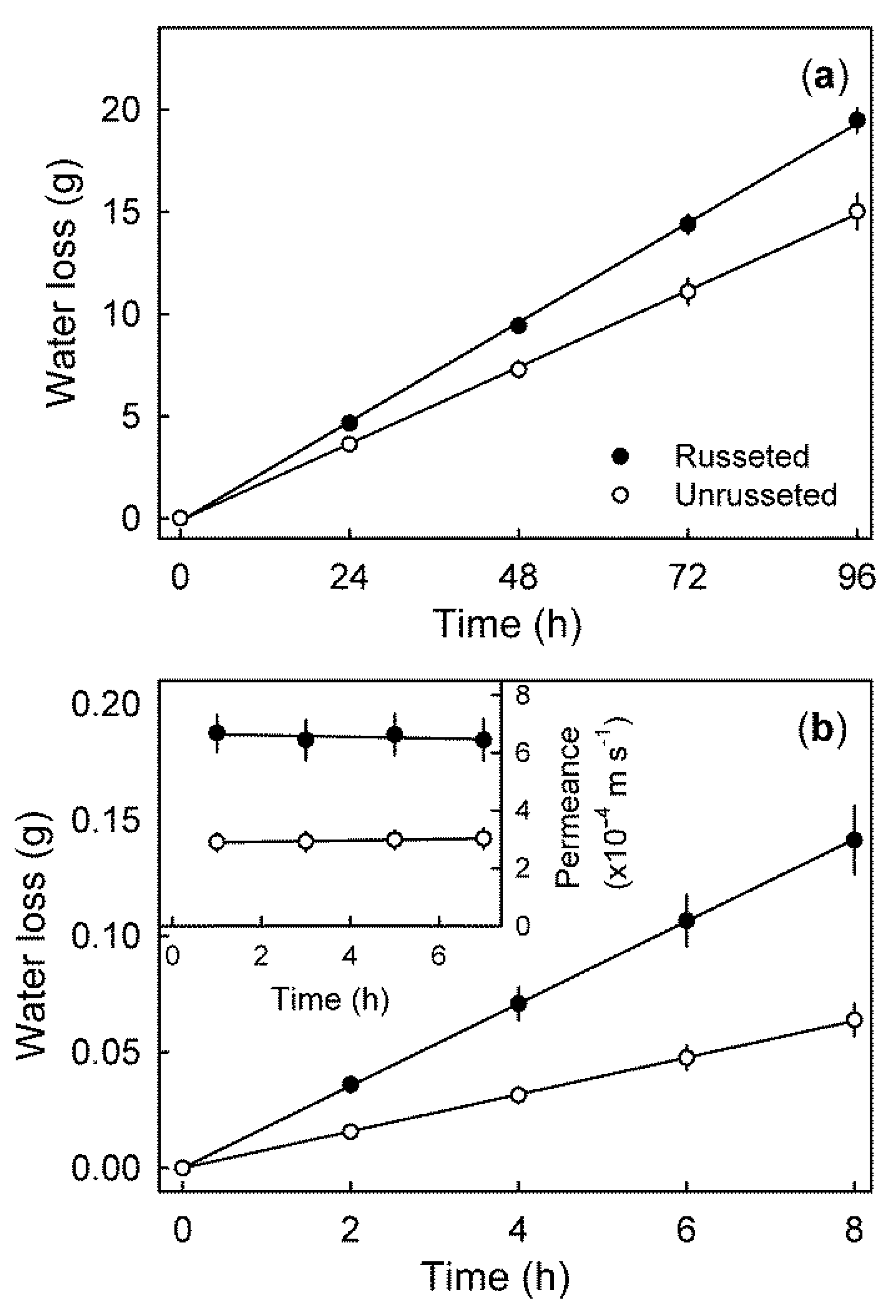
Time course of transpiration by whole fruits with extreme (>50%) (russeted) and with minimal (<25%) russet (not russeted) (a) and through epidermal sections (ES) excised from the cheek of mature ‘Apple’ mango fruit with and without russeting (b). Permeance of water vapour diffusion through the ES (b inset) was calculated under conditions of steady state water loss. Data represent means ± SE of a minimum of 10 replicates.
Figure 6.
Time course of transpiration by whole fruits with extreme (>50%) (russeted) and with minimal (<25%) russet (not russeted) (a) and through epidermal sections (ES) excised from the cheek of mature ‘Apple’ mango fruit with and without russeting (b). Permeance of water vapour diffusion through the ES (b inset) was calculated under conditions of steady state water loss. Data represent means ± SE of a minimum of 10 replicates.
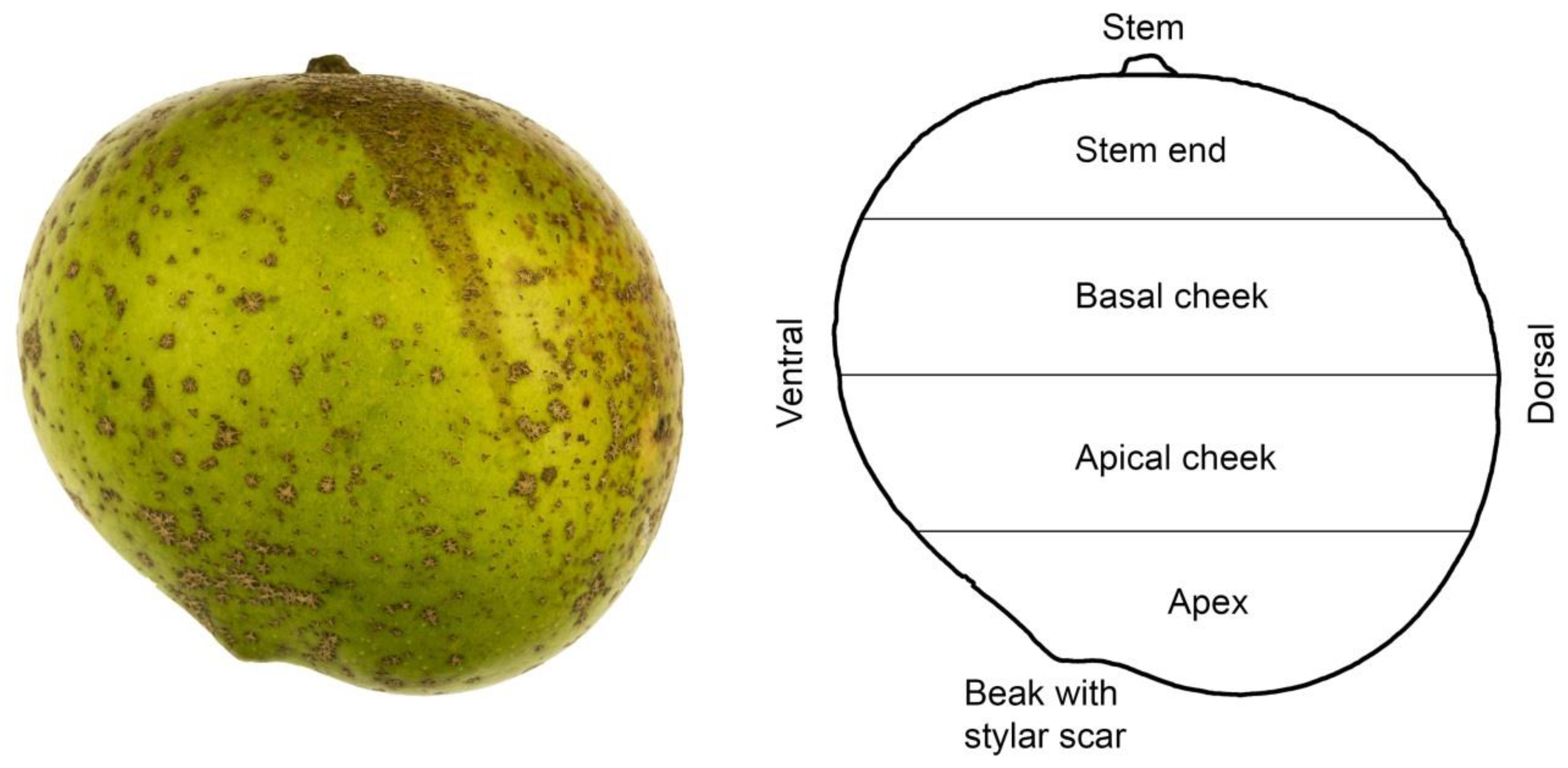
Figure 7.
Photograph of ‘Apple’ mango and sketch illustrating the nomenclature used to describe regions of the fruit surface [
41].
Figure 7.
Photograph of ‘Apple’ mango and sketch illustrating the nomenclature used to describe regions of the fruit surface [
41].
Table 1.
Percent distribution of russet on ‘Apple’ mango fruit along the stem/apex axis on either the blushed or the non-blushed sides of the fruit. Percentage of russet was quantified using image analysis following photography. For the different regions of the fruit, see Figure 7.
Table 1.
Percent distribution of russet on ‘Apple’ mango fruit along the stem/apex axis on either the blushed or the non-blushed sides of the fruit. Percentage of russet was quantified using image analysis following photography. For the different regions of the fruit, see Figure 7.
| Fruit Region | Russeted Area (%) |
|---|
| Blushed Side | Non-Blushed Side | Mean Side |
|---|
| Stem end | 85.8 ± 6.9 | 87.2 ± 5.7 | 86.5 ± 4.4 a z |
| Basal cheek | 47.7 ± 5.7 | 43.8 ± 5.5 | 45.8 ± 4.0 b |
| Apical cheek | 19.2 ± 5.8 | 18.4 ± 5.3 | 18.8 ± 3.9 c |
| Apex | 15.4 ± 6.1 | 17.2 ± 6.3 | 16.3 ± 4.3 c |
| Mean Fruit region | 42.0 ± 4.5 a | 41.7 ± 4.4 a | |
Table 2.
Russeting in ventral and dorsal regions of ‘Apple’ mango from three different sites. The sites were selected because they differed in elevation. Russeting was quantified using a five-score rating scheme. Score 0: 0% of the fruit surface area russeted; score 1: 1–10% russeted area; score 2: 11–25% russeted area; score 3: 26–50% russeted area; and score 4: 51–100% russeted area. For ventral and dorsal regions of the fruit, see Figure 7.
Table 2.
Russeting in ventral and dorsal regions of ‘Apple’ mango from three different sites. The sites were selected because they differed in elevation. Russeting was quantified using a five-score rating scheme. Score 0: 0% of the fruit surface area russeted; score 1: 1–10% russeted area; score 2: 11–25% russeted area; score 3: 26–50% russeted area; and score 4: 51–100% russeted area. For ventral and dorsal regions of the fruit, see Figure 7.
| Site | Extent of Russet (Rating Score) |
|---|
| Ventral | Dorsal | Mean Region |
|---|
| Kaiti | 2.9 ± 0.1 | 3.5 ± 0.1 | 3.2 ± 0.1 a z |
| Mumbuni | 2.0 ± 0.1 | 2.8 ± 0.1 | 2.4 ± 0.1 b |
| Yeemulwa | 1.6 ± 0.1 | 2.3 ± 0.1 | 1.9 ± 0.1 c |
| Mean Site | 2.2 ± 0.1 b | 2.8 ± 0.0 a | |
Table 3.
Russeting of peripheral (exposed) or central (shaded) ‘Apple’ mango fruit in the canopy at three different sites. The sites were selected because they differ in elevation. Russeting was quantified using a five-score rating scheme. Score 0: 0% of the fruit surface area russeted; score 1: 1–10% russeted area; score 2: 11–25% russeted area; score 3: 26–50% russeted area; and score 4: 51–100% russeted area.
Table 3.
Russeting of peripheral (exposed) or central (shaded) ‘Apple’ mango fruit in the canopy at three different sites. The sites were selected because they differ in elevation. Russeting was quantified using a five-score rating scheme. Score 0: 0% of the fruit surface area russeted; score 1: 1–10% russeted area; score 2: 11–25% russeted area; score 3: 26–50% russeted area; and score 4: 51–100% russeted area.
| Site | Russeting (Rating Score) |
|---|
| Exposed Fruits | Shaded Fruits | Mean Fruit Position |
|---|
| Kaiti | 3.7 ± 0.0 | 3.7 ± 0.0 | 3.7 ± 0.0 a z |
| Mumbuni | 2.3 ± 0.1 | 2.5 ± 0.1 | 2.4 ± 0.1 b |
| Yeemulwa | 2.0 ± 0.1 | 1.8 ± 0.1 | 1.9 ± 0.1 c |
| Mean Site | 2.7 ± 0.1 a | 2.7 ± 0.1 a | |
Table 4.
Effect of fruit position within the tree canopy on russeting of ‘Apple’ mango in different sites. Fruit positions were: top (>2 m above the ground), middle (1–2 m), and bottom (0.5–1 m). Russeting was quantified using a five-score rating scheme. Score 0: 0% of the fruit surface area russeted; score 1: 1–10% russeted area; score 2: 11–25% russeted area; score 3: 26–50% russeted area; and score 4: 51–100% russeted area.
Table 4.
Effect of fruit position within the tree canopy on russeting of ‘Apple’ mango in different sites. Fruit positions were: top (>2 m above the ground), middle (1–2 m), and bottom (0.5–1 m). Russeting was quantified using a five-score rating scheme. Score 0: 0% of the fruit surface area russeted; score 1: 1–10% russeted area; score 2: 11–25% russeted area; score 3: 26–50% russeted area; and score 4: 51–100% russeted area.
| Site | Russeting (Rating Score) |
|---|
| Top | Middle | Bottom | Mean Fruit position |
|---|
| Kaiti | 3.9 ± 0.0 a z | 3.7 ± 0.0 b | 3.5 ± 0.1 c | 3.7 ± 0.0 |
| Mumbuni | 3.1 ± 0.1 a | 2.3 ± 0.1 b | 1.9 ± 0.1 c | 2.4 ± 0.0 |
| Yeemulwa | 2.5 ± 0.1 a | 1.7 ± 0.1 b | 1.0 ± 0.1 c | 1.7 ± 0.0 |
| Mean Site | 3.0 ± 0.0 | 2.4 ± 0.0 | 1.9 ± 0.0 | |
Table 5.
Effect of geographical orientation (aspect) of ‘Apple’ mango on russeting. Fruits were sampled from north-, south-, east-, and west-facing sides of the canopy. The tree rows were aligned perpendicularly to the slope and N, S, E, and W positions. Russeting was quantified using a five-score rating scheme. Score 0: 0% of the fruit surface area russeted; score 1: 1–10% russeted area; score 2: 11–25% russeted area; score 3: 26–50% russeted area; and score 4: 51–100% russeted area.
Table 5.
Effect of geographical orientation (aspect) of ‘Apple’ mango on russeting. Fruits were sampled from north-, south-, east-, and west-facing sides of the canopy. The tree rows were aligned perpendicularly to the slope and N, S, E, and W positions. Russeting was quantified using a five-score rating scheme. Score 0: 0% of the fruit surface area russeted; score 1: 1–10% russeted area; score 2: 11–25% russeted area; score 3: 26–50% russeted area; and score 4: 51–100% russeted area.
| Site | Russeting (Rating Score) |
|---|
| North | South | East | West | Mean Aspect |
|---|
| Kaiti | 3.6 ± 0.0 | 3.6 ± 0.0 | 3.8 ± 0.0 | 3.5 ± 0.1 | 3.6 ± 0.0 a z |
| Mumbuni | 2.4 ± 0.1 | 2.5 ± 0.1 | 2.2 ± 0.1 | 2.6 ± 0.1 | 2.4 ± 0.0 b |
| Yeemulwa | 1.8 ± 0.1 | 1.7 ± 0.1 | 1.6 ± 0.1 | 1.8 ± 0.1 | 1.7 ± 0.0 c |
| Mean Site | 2.5 ± 0.1 a | 2.6 ± 0.1 a | 2.5 ± 0.1 a | 2.6 ± 0.1 a | |
Table 6.
Russeting of ‘Apple’ Mango at ten different sites across Kenya. Russeting was quantified using a five-score rating scheme: score 0: 0% of the fruit surface area russeted; score 1: 1–10% russeted area; score 2: 11–25% russeted area; score 3: 26–50% russeted area; and score 4: 51–100% russeted area.
Table 6.
Russeting of ‘Apple’ Mango at ten different sites across Kenya. Russeting was quantified using a five-score rating scheme: score 0: 0% of the fruit surface area russeted; score 1: 1–10% russeted area; score 2: 11–25% russeted area; score 3: 26–50% russeted area; and score 4: 51–100% russeted area.
| Site | Maturity (Days after Full Bloom) | Rating (Score) |
|---|
| Thika | 196 | 3.6 ± 0.1 a z |
| Kaiti | 189 | 3.5 ± 0.1 a |
| Machakos | 226 | 3.4 ± 0.1 ab |
| Kasafari | 166 | 3.4 ± 0.1 ab |
| Chepsigot | 146 | 3.2 ± 0.1 b |
| Kambirwa | 136 | 3.1 ± 0.1 b |
| Mumbuni | 175 | 2.3 ± 0.1 c |
| Yeemulwa | 180 | 1.9 ± 0.1 d |
| Malindi | 113 | 0.2 ± 0.0 e |
| Garissa | 111 | 0.1 ± 0.0 e |
Table 7.
Pearson correlation between climatic variables and russeting for ‘Apple’ mango fruits grown at ten sites throughout Kenya. Climatic variables include rainfall amount, number (n) of rainy days, relative humidity, maximum, minimum, and mean daily temperatures, and heat sum. Heat sum is the sum of mean temperatures above the base temperature of 16 °C. Number of cold nights is the sum of the number of days when the minimum temperature was less than 16 °C. The growth season was divided into three periods of equal duration at each site (early, middle, and late phases). “Cumulative” refers to the entire growth period. Climatic data were obtained from the NASA Langley Research Centre (LaRC) POWER Project.
Table 7.
Pearson correlation between climatic variables and russeting for ‘Apple’ mango fruits grown at ten sites throughout Kenya. Climatic variables include rainfall amount, number (n) of rainy days, relative humidity, maximum, minimum, and mean daily temperatures, and heat sum. Heat sum is the sum of mean temperatures above the base temperature of 16 °C. Number of cold nights is the sum of the number of days when the minimum temperature was less than 16 °C. The growth season was divided into three periods of equal duration at each site (early, middle, and late phases). “Cumulative” refers to the entire growth period. Climatic data were obtained from the NASA Langley Research Centre (LaRC) POWER Project.
| Weather Parameter | Pearson Coefficients of Correlation (r) |
|---|
| Time Period |
|---|
| Early | Middle | Late | Cumulative |
|---|
| Rainfall (mm) | 0.39 ns z | 0.44 ns | 0.26 ns | 0.60 ns |
| Rainy days (n) | 0.66 * | 0.67 * | 0.79 ** | 0.90 *** |
| Relative humidity (%) | −0.27 ns | 0.11 ns | 0.08 ns | −0.05 ns |
| Maximum temperature (°C) | −0.56 ns | −0.71 * | −0.85 ** | −0.74 * |
| Minimum temperature (°C) | −0.94 *** | −0.93 *** | −0.94 *** | −0.94 *** |
| Mean temperature (°C) | −0.96 *** | −0.95 *** | −0.96 *** | −0.96 *** |
| Heat sum (°Cd) | −0.57 ns | −0.67 * | −0.72 * | −0.67 * |
| Dew point temperature (°C) | −0.88 *** | −0.85 ** | −0.91 *** | −0.92 *** |
| Cold nights (n) | 0.88 *** | 0.73 * | 0.63 ns | 0.84 ** |











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}