Glycine Betaine Accumulation, Significance and Interests for Heavy Metal Tolerance in Plants

 ,
,  , , , ,
, , , ,  and
and
Abstract
:1. Introduction
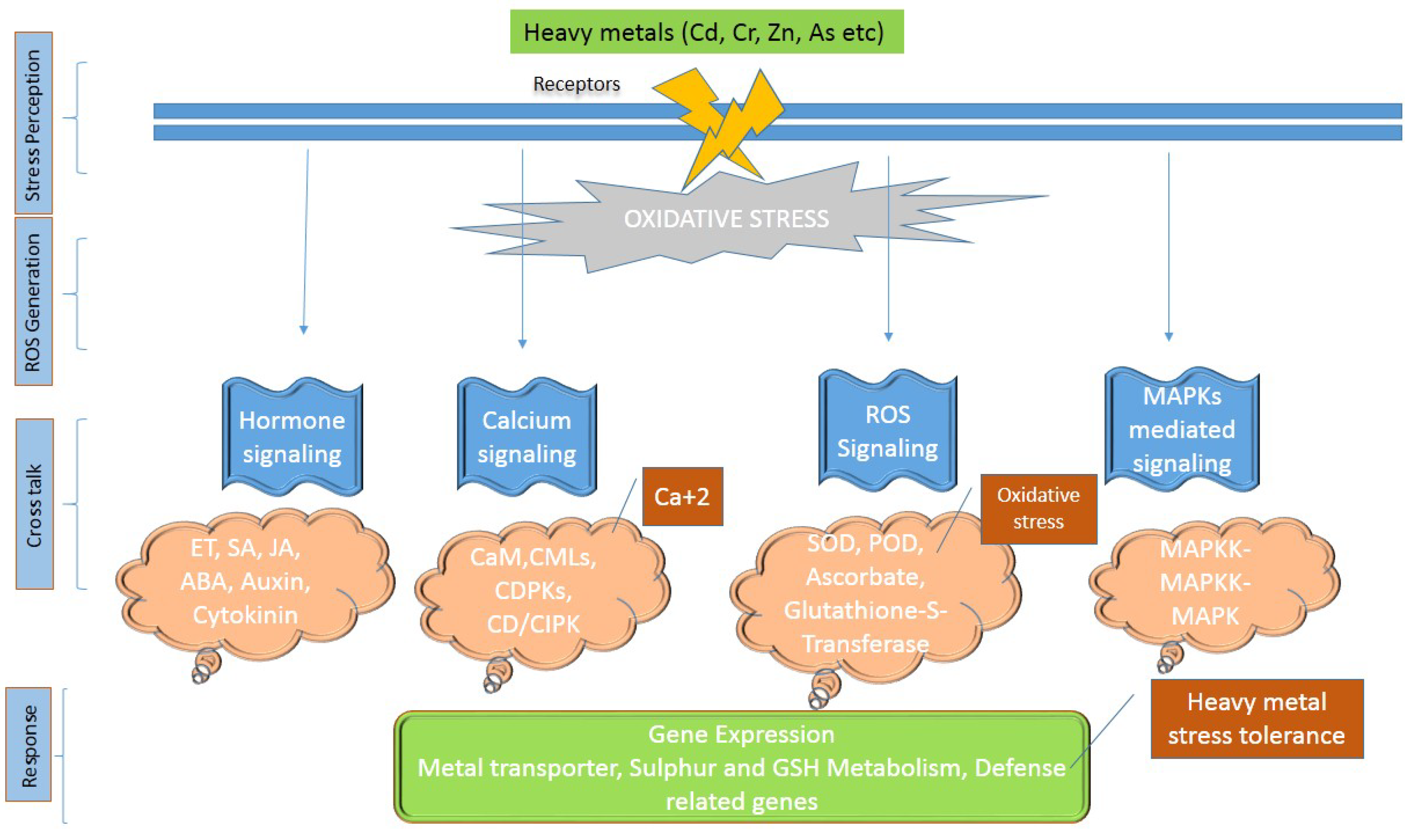
2. Signaling Response in Plants against Heavy Metal Stress
3. Accumulation of Glycine Betaine (GB) as an Emerging Signal Molecule in Plants
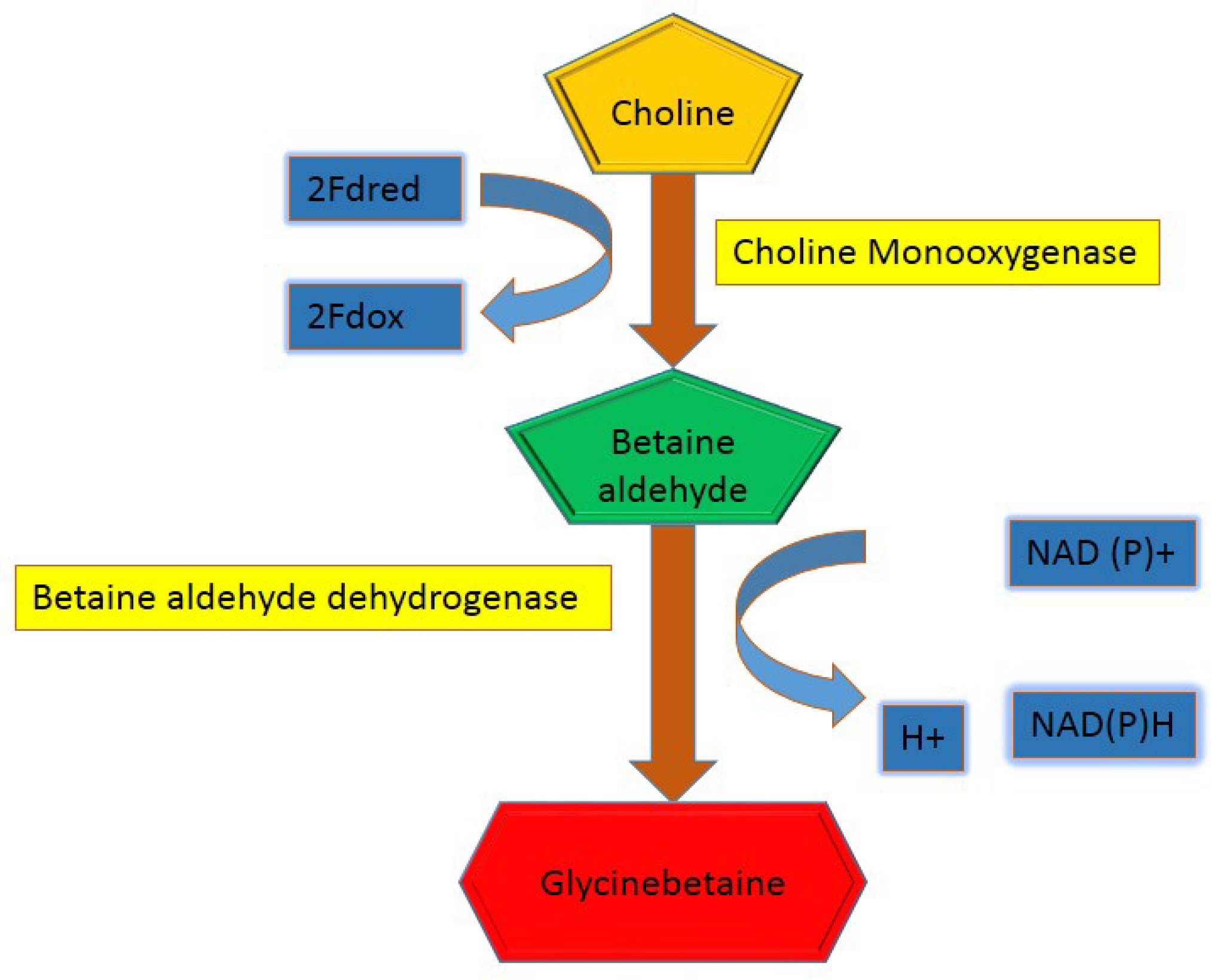
4. Biosynthesis of Glycine Betaine in Plants
5. Role of Glycine Betaine in Plant Growth
6. Promotive Role of Glycine Betaine in Mitigating Heavy Metal Stress in Plants
6.1. Improvement in Plant Growth and Biomass
6.2. Enhancement in Rate of Photosynthesis
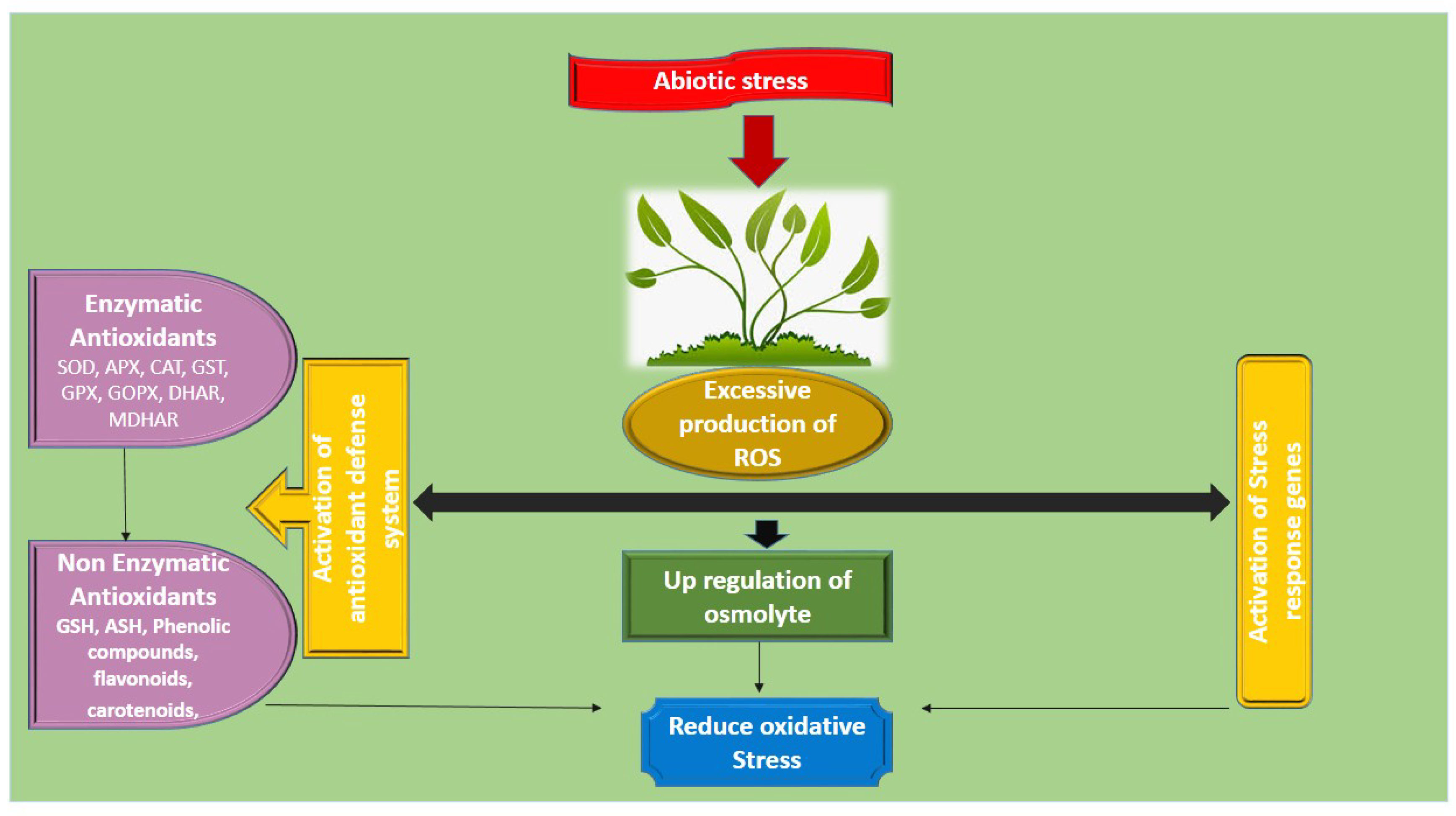
6.3. Up-Regulation of Antioxidant Defense System
6.4. Upgrading of Metal and Mineral Uptake
6.5. Alleviation of Electrolyte Leakage (EL), Malondialdehyde (MDA) and Hydrogen Peroxide (H2O2)
7. Involvement of Glycine Betaine against Combinations of Abiotic Stresses in Plants
8. Application Methods of Glycine Betaine
8.1. Foliar Application of GB
8.2. Pre-Sowing Seed Treatments
9. Role of Glycine Betaine in Crop Improvement
10. Genetic Engineering
11. Conclusions
Funding
Acknowledgments
Conflicts of Interest
References
- Dixit, R.; Wasiullah, E.Y.; Malaviya, D.; Pandiyan, K.; Singh, U.; Sahu, A.; Shukla, R.; Singh, B.; Rai, J.; Sharma, P.; et al. Bioremediation of heavy metals from soil and aquatic environment: An overview of principles and criteria of fundamental processes. Sustainability 2015, 7, 2189–2212. [Google Scholar] [CrossRef] [Green Version]
- Peng, W.; Li, X.; Xiao, S.; Fan, W. Review of remediation technologies for sediments contaminated by heavy metals. J. Soils Sediments 2018, 18, 1701–1719. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Santanen, A.; Jaakkola, S.; Ekholm, P.; Hartikainen, H.; Stoddard, F.L.; Mäkelä, P.S.A. Biomass yield and quality of bioenergy crops grown with synthetic and organic fertilizers. Biomass Bioenerg. 2013, 59, 477–485. [Google Scholar] [CrossRef]
- Seleiman, M.F.; Santanen, A.; Kleemola, J.; Stoddard, F.L.; Mäkelä, P.S.A. Improved sustainability of feedstock production with sludge and interacting mychorriza. Chemosphere 2013, 91, 1236–1242. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Santanen, A.; Stoddard, F.L.; Mäkelä, P.S.A. Feedstock quality and growth of bioenergy crops fertilized with sewage sludge. Chemosphere 2012, 89, 1211–1217. [Google Scholar] [CrossRef] [PubMed]
- Shahid, M.J.; Ali, S.; Shabir, G.; Siddique, M.; Rizwan, M.; Seleiman, M.F.; Afzal, M. Comparing the performance of four macrophytes in bacterial assisted floating treatment wetlands for the removal of trace metals (Fe, Mn, Ni, Pb, and Cr) from polluted river water. Chemosphere 2020, 243, 125353. [Google Scholar] [CrossRef] [PubMed]
- Seleiman, M.F.; Kheir, A.S. Maize productivity, heavy metals uptake and their availability in contaminated clay and sandy alkaline soils as affected by inorganic and organic amendments. Chemosphere 2018, 204, 514–522. [Google Scholar] [CrossRef]
- Salem, H.S.; Abdel-Salam, A.; Abdel-Salam, M.A.; Seleiman, M.F. Phytoremediation of metal and metalloids from contaminated soil. In Plants under Metal and Metalloid Stress- Responses, Tolerance and Remediation; Hasanuzzaman, M., Nahar, K., Fujita, M., Eds.; Series Soil Biology; Springer Nature Pte Ltd.: Singapore, 2018; Chapter 9; pp. 249–262. [Google Scholar]
- Seleiman, M.F.; Selim, S.; Jaakkola, S.; Mäkelä, P. Chemical composition and in vitro digestibility of whole-crop maize fertilized with synthetic fertilizer or digestate and harvested at two maturity stages in boreal growing conditions. Agric. Food Sci. 2017, 26, 47–55. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Salam, A.; Salem, H.M.; Abdel-Salam, M.A.; Seleiman, M.F. Phyto and chemical removal of heavy metal-contaminated soils. In Heavy Metal Contamination of Soils: Monitoring and Remediation; Sherameti, I., Varma, A., Eds.; Series Soil Biology; Springer: Cham, Switzerland, 2015; Volume 44, pp. 299–308. [Google Scholar]
- Seleimana, M.F.; Santanen, A.; Mäkelä, P.S.A. Recycling sludge on cropland as fertilizer—Advantages and risks. Resour. Conserv. Recycl. 2020, 155, 104647. [Google Scholar] [CrossRef]
- Qayyum, M.F.; Rehman, M.Z.; Ali, S.; Rizwan, M.; Naeem, A.; Maqsood, M.A.; Khalid, H.; Rinklebe, J.; Ok, Y.S. Residual effcts of monoammonium phosphate, gypsum and elemental sulfur on cadmium phytoavailability and translocation from soil to wheat in an effluent irrigated field. Chemosphere 2017, 174, 515–523. [Google Scholar] [CrossRef]
- Ali, S.; Rizwan, M.; Zaid, A.; Arif, M.S.; Yasmeen, T.; Hussain, A.; Shahid, M.R.; Bukhari, S.A.H.; Hussain, S.; Abbasi, G.H. 5-Aminolevulinic Acid-Induced Heavy Metal Stress Tolerance and Underlying Mechanisms in Plants. J. Plant Growth Regul. 2018, 37, 1423–1433. [Google Scholar] [CrossRef]
- Tchounwou, P.B.; Yedjou, C.G.; Patlolla, A.K.; Sutton, D.J. Heavy metals toxicity and the environment. Mol. Clin. Environ. Toxicol. 2012, 101, 133–164. [Google Scholar]
- Taiwo, A.M.; Gbadebo, A.M.; Oyedepo, J.A.; Ojekunle, Z.O.; Alo, O.M.; Oyeniran, A.A.; Onalaja, O.J.; Ogunjimi, D.; Taiwo, O.T. Bioremediation of industrially contaminated soil using compost and plant technology. J. Hazard Mater. 2016, 304, 166–172. [Google Scholar] [CrossRef] [PubMed]
- Rizwan, M.; Ali, S.; Adrees, M.; Ibrahim, M.; Tsang, D.C.; Rehman, M.Z.; Zahir, Z.A.; Rinklebe, J.; Tack, F.M.; Ok, Y.S. A critical review on effcts, tolerance mechanisms and management of cadmium in vegetables. Chemosphere 2017, 182, 90–105. [Google Scholar] [CrossRef]
- Jacob, J.M.; Karthik, C.; Saratale, R.G.; Smita, S.; Kumar, D.; Prabakar, D.; Kadirvelu, K.; Pugazhendhi, A. Biological approaches to tackle heavy metal pollution: A survey of literature. J. Environ. Manag. 2018, 217, 56–70. [Google Scholar] [CrossRef]
- Huang, T.L.; Huang, L.Y.; Fu, S.F.; Trinh, N.N.; Huang, H.J. Genomic profiling of rice roots with short-and long-term chromium stress. Plant Mol. Biol. 2014, 86, 157–170. [Google Scholar] [CrossRef]
- Kohzadi, S.; Shahmoradi, B.; Ghaderi, E.; Loqmani, H.; Maleki, A. Concentration, Source, and Potential Human Health Risk of Heavy Metals in the Commonly Consumed Medicinal Plants. Biol. Trace Elem. Res. 2019, 187, 41–50. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Ilahi, I. Environmental chemistry and ecotoxicology of hazardous heavy metals: Environmental persistence, toxicity, and bioaccumulation. J. Chem. 2019, 2019, 6730305. [Google Scholar] [CrossRef] [Green Version]
- Sofy, M.; Seleiman, M.F.; Alhammad, B.A.; Alharbi, B.A.; Mohamed, H.I. Minimizing adverse effects of Pb stress on maize yield, macro elements, and physiological and biochemical traits by combined treatment with jasmonic acid, salicylic acid, and proline. Agronomy 2020, 10, 699. [Google Scholar] [CrossRef]
- Saleem, M.H.; Ali, S.; Seleiman, M.F.; Rizwan, M.; Rehman, M.; Akram, N.A.; Liu, L.; Alotaibi, M.; Al-Ashkar, I.; Mubushar, M. Assessing the Correlations between Different Traits in Copper-Sensitive and Copper-Resistant Varieties of Jute (Corchorus capsularis L.). Plants 2019, 8, 545. [Google Scholar] [CrossRef] [Green Version]
- Seleiman, M.F.; Alotaibi, M.; Alhammad, B.; Alharbi, B.; Refay, Y.; Badawy, S. Effects of ZnO nanoparticles and biochar of rice straw and cow manure on characteristics of contaminated soil and sunflower productivity, oil quality, and heavy metals uptake. Agronomy 2020, 10, 790. [Google Scholar] [CrossRef]
- Chandra, R.; Yadav, S.; Yadav, S. Phytoextraction potential of heavy metals by native wetland plants growing on chlorolignin containing sludge of pulp and paper industry. Ecol. Eng. 2017, 98, 134–145. [Google Scholar] [CrossRef]
- Willscher, S.; Jablonski, L.; Fona, Z.; Rahmi, R.; Wittig, J. Phytoremediation experiments with Helianthus tuberosus under different pH and heavy metal soil concentrations. Hydrometallurgy 2017, 168, 153–158. [Google Scholar] [CrossRef]
- Nayak, A.K.; Panda, S.S.; Basu, A.; Dhal, N.K. Enhancement of toxic Cr (VI), Fe, and other heavy metals phytoremediation by the synergistic combination of native Bacillus cereus strain and Vetiveria zizanioides L. Int. J. Phytoremediat. 2018, 20, 682–691. [Google Scholar] [CrossRef] [PubMed]
- Malar, S.; Vikram, S.S.; JC Favas, P.; Peruma, V. Lead heavy metal toxicity induced changes on growth and antioxidative enzymes level in water hyacinths [Eichhornia crassipes (Mart)]. Bot. Stud. 2014, 55, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Tauqeer, H.M.; Ali, S.; Rizwan, M.; Ali, Q.; Saeed, R.; Iftikhar, U.; Ahmad, R.; Farid, M.; Abbasi, G.H. Phytoremediation of heavy metals by Alternanthera bettzickiana: Growth and physiological response. Ecotoxicol. Environ. Saf. 2016, 126, 138–146. [Google Scholar] [CrossRef]
- Abbas, T.; Rizwan, M.; Ali, S.; Rehman, M.Z.; Qayyum, M.F.; Abbas, F.; Hannan, F.; Rinklebe, J.; Ok, Y.S. Effct of biochar on cadmium bioavailability and uptake in wheat (Triticum aestivum L,) grown in a soil with aged contamination. Ecotoxicol. Environ. Saf. 2017, 140, 37–47. [Google Scholar] [CrossRef]
- Khan, A.; Fatima, H.; Ghania, A.; Nadeem, M.; Aziz, A.; Hussain, M.; Ikram, M. Improving Salinity Tolerance in Brassica (Brassica napus var, Bsa and Brassica campestris var, Toria) by Exogenous Application of Proline and Glycine Betaine. Pak. J. Sci. Ind. Res. Ser. B Boil. Sci. 2018, 1, 1–8. [Google Scholar]
- Peters, D.E.; Eebu, C.; Nkpaa, K.N. Potential Human Health Risk Assessment of Heavy Metals via Consumption of Root Tubers from Ogoniland, Rivers State, Nigeria. Biol. Trace Elem. Res. 2018, 186, 568–578. [Google Scholar] [CrossRef]
- Intawongse, M.; Kongchouy, N.; Dean, J.R. Bioaccessibility of heavy metals in the seaweed Caulerparacemosa var, corynephora: Human health risk from consumption. Instrum. Sci. Technol. 2018, 46, 628–644. [Google Scholar] [CrossRef] [Green Version]
- Chibuike, G.U.; Obiora, S.C. Heavy metal polluted soils: Effect on plants and bioremediation methods. Appl. Environ. Soil Sci. 2014, 2014, 12. [Google Scholar] [CrossRef] [Green Version]
- Ashfaque, F.; Inam, A.; Sahay, S.; Iqbal, S. Influence of Heavy Metal Toxicity on Plant Growth, Metabolism and Its Alleviation by Phytoremediation—A Promising Technology. J. Agric. Ecol. Res. Int. 2016, 6, 1–19. [Google Scholar] [CrossRef]
- Khan, M.; Iqbal, R.; Khan, N.A. Reactive Oxygen Species and Antioxidant Systems in Plants: Role and Regulation under Abiotic Stress; Springer: Cham, Switzerland, 2017. [Google Scholar]
- Seneviratne, M.; Rajakaruna, N.; Rizwan, M.; Madawala, H.M.S.P.; Ok, Y.S.; Vithanage, M. Heavy metal-induced oxidative stress on seed germination and seedling development: A critical review. Environ. Geochem. Health 2017, 41, 1813–1831. [Google Scholar] [CrossRef] [PubMed]
- Farid, M.; Ali, S.; Rizwan, M.; Ali, Q.; Saeed, R.; Nasir, T.; Abbasi, G.H.; Rehmani, M.I.; Ata-Ul-Karim, S.T.; Bukhari, S.A.; et al. Phytomanagement of chromium contaminated soils through sunflower under exogenously applied 5-aminolevulinic acid. Ecotoxicol. Environ. Saf. 2018, 151, 255–265. [Google Scholar] [CrossRef] [PubMed]
- Genthe, B.; Kapwata, T.; Le Roux, W.; Chamier, J.; Wright, C.Y. The reach of human health risks associated with metals/metalloids in water and vegetables along a contaminated river catchment: South Africa and Mozambique. Chemosphere 2018, 199, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Rizwan, M.; Ali, S.; Rehman, M.Z.; Rinklebe, J.; Tsang, D.C.W.; Bashir, A.; Maqbool, A.; Tack, F.M.G.; Ok, Y.S. Cadmium phytoremediation potential of Brassica crop species: A review. Sci. Total Environ. 2018, 631–632, 1175–1191. [Google Scholar] [CrossRef] [PubMed]
- Babst-Kosteckaa, A.A.; Waldmannc, P.; Frérotd, H.; Vollenweider, P. Plant adaptation to metal polluted environments—Physiological, morphological, and evolutionary insights from Biscutella laevigata. Environ. Exp. Bot. 2016, 127, 1–13. [Google Scholar] [CrossRef]
- Shanying, H.E.; Xiaoe, Y.; Zhenli, H.E.; Baligar, V.C. Morphological and Physiological Responses of Plants to Cadmium Toxicity: A Review. Pedosphere 2017, 27, 421–438. [Google Scholar]
- Khan, A.; Khan, S.; Khan, M.A.; Qamar, Z.; Waqas, M. The uptake and bioaccumulation of heavy metals by food plants, their effects on plants nutrients, and associated health risk: A review. Environ. Sci. Pollut. Res. 2015, 22, 13772–13799. [Google Scholar] [CrossRef]
- Anjum, S.A.; Ashraf, U.; Khan, I.; Tanveer, M.; Ali, M.; Hussain, I.; Wang, L.C. Chromium and aluminum phyto-toxicity in maize; morphophysiological responses and metal uptake. Clean-Soil Air Water 2016, 44, 915–1084. [Google Scholar] [CrossRef]
- Anjum, S.A.; Tanveer, M.; Hussain, S.; Ashraf, U.; Khan, I.; Wang, L.C. Alteration in growth, leaf gas exchange, and photosynthetic pigments of maize plants under combined cadmium and arsenic stress. Water Air Soil Pollut. 2017, 228, 13. [Google Scholar] [CrossRef]
- Pongrac, P.; Zhao, F.J.; Razinger, J.; Zrimec, A.; Regvar, M. Physiological responses to Cd and Zn in two Cd/Zn hyperaccumulating Thlaspi species. Environ. Exp. Bot. 2009, 66, 479–486. [Google Scholar] [CrossRef]
- Lämke, J.; Bäurle, I. Epigenetic and chromatin-based mechanisms in environmental stress adaptation and stress memory in plants. Genome Biol. 2017, 18, 124. [Google Scholar] [CrossRef] [PubMed]
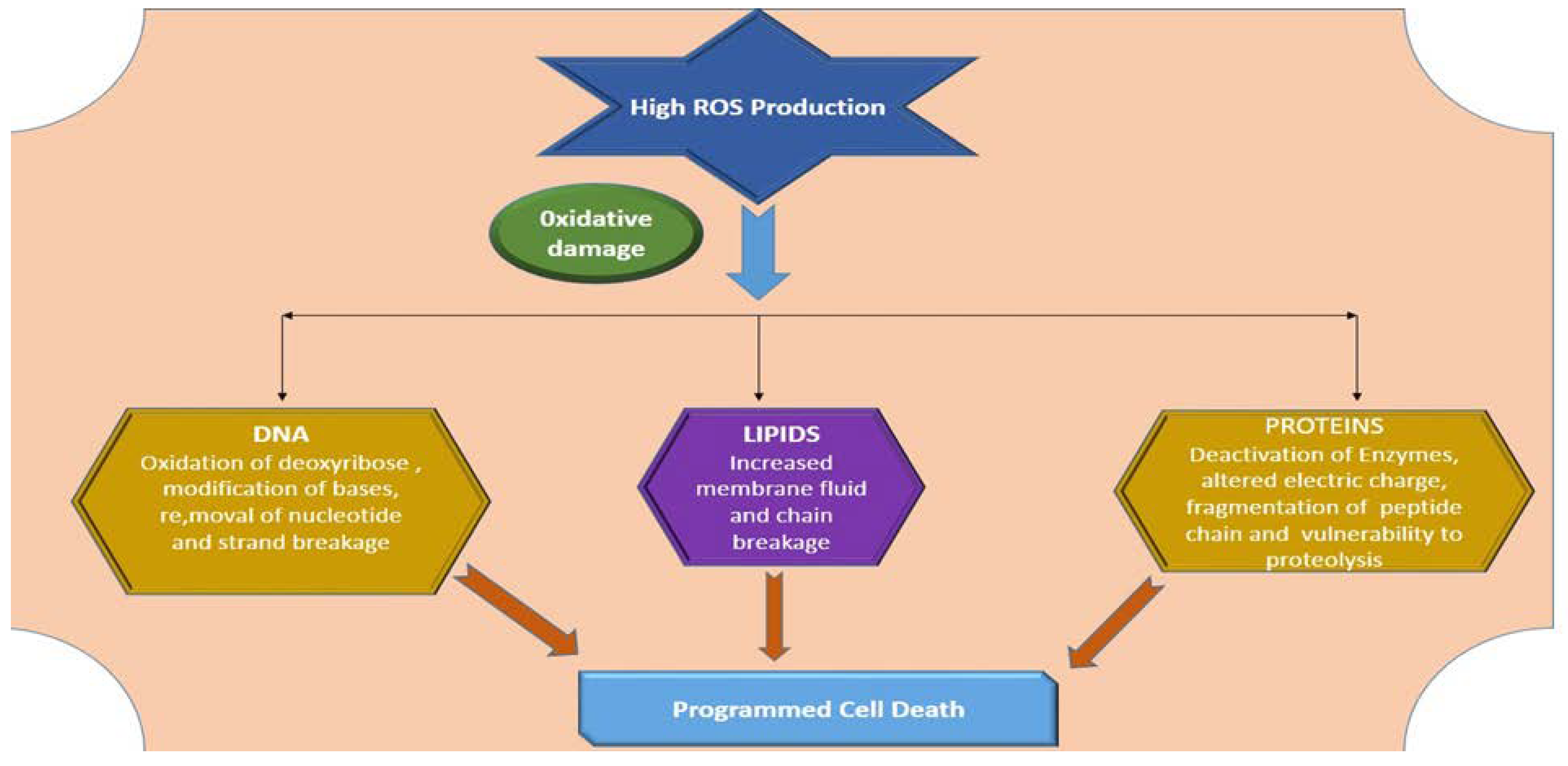
- Demidchik, V. Mechanisms of oxidative stress in plants: From classical chemistry to cell biology. Environ. Exp. Bot. 2015, 109, 212–228. [Google Scholar] [CrossRef]
- Zaid, A.; Wani, S.H. Reactive Oxygen Species Generation, Scavenging and Signaling in Plant Defense Responses. In Bioactive Molecules in Plant Defense; Springer Science and Business Media LLC: Berlin, Germany, 2019; pp. 111–132. [Google Scholar]
- Noctor, G.; Foyer, C.H. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279. [Google Scholar] [CrossRef]
- Steffens, B. The role of ethylene and ROS in salinity, heavy metal, and flooding responses in rice. Front. Plant Sci. 2014, 5, 1–5. [Google Scholar] [CrossRef]
- Xu, J.; Zhao, M.; Pei, L.; Zhang, R.; Liu, X.; Wei, L.; Yang, M.; Xu, Q. Oxidative stress and DNA damage in a long-term hexavalent chromiumexposed population in North China: A cross-sectional study. BMJ Open 2018, 8, 021470. [Google Scholar] [CrossRef] [Green Version]
- Hall, J.L. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot. 2002, 53, 1–11. [Google Scholar] [CrossRef]
- Hossain, M.A.; Piyatida, P.; da Silva, J.A.T.; Fujita, M. Molecular mechanism of heavy metal toxicity and tolerance in plants: Central role of glutathione in detoxifiation of reactive oxygen species and methylglyoxal and in heavy metal chelation. J. Bot. 2012, 2012, 37. [Google Scholar] [CrossRef]
- Kumar, S.; Trivedi, P.K. Heavy Metal Stress Signalling in Plants. Plant Metal Interaction (Emerging Remediation Techniques); Elsevier: Amsterdam, The Netherlands, 2016; pp. 585–603. [Google Scholar]
- Tiwari, S.; Lata, C. Heavy Metal Stress, Signaling, and Tolerance Due to Plant-Associated Microbes: An Overview. Front. Plant Sci. 2018, 9, 452. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.H.; Cao, L.; Dou, S.Z. Bioaccumulation of heavy metals and health risk assessment in three benthic bivalves along the coast of Laizhou Bay, China. Mar. Pollut. Bull. 2017, 117, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Dubey, S.; Shri, M.; Misra, P.; Lakhwani, D.; Bag, S.K.; Asif, M.H. Heavy metals induce oxidative stress and genome-wide modulation in transcriptome of rice root. Funct. Integr. Genom. 2014, 14, 401–417. [Google Scholar] [CrossRef] [PubMed]
- Singh, I.; Shah, K. Exogenous application of methyl jasmonate lowers the effect of cadmium-induced oxidative injury in rice seedlings. Phytochemistry 2014, 108, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Dutta, S.; Mitra, M.; Agarwal, P.; Mahapatra, K.; De, S.; Sett, U.; Roy, S. Oxidative and genotoxic damages in plants in response to heavy metal stress and maintenance of genome stability. Plant Signal. Behav. 2018, 13, e1460048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiwari, S.; Lata, C.; Chauhan, S.; Prasad, P.; Prasad, M.A. Functional genomic perspective on drought signalling and its crosstalk with phytohormone-mediated signalling pathways in plants. Curr. Genom. 2017, 18, 469–482. [Google Scholar] [CrossRef]
- Zhu, J.K. Salt and drought stress signal transduction in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2002, 53, 247–273. [Google Scholar] [CrossRef] [Green Version]
- Rhodes, D.; Hanson, A.D. Quaternary ammonium and tertiary sulfonium compounds in higher-plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1993, 44, 357–384. [Google Scholar] [CrossRef]
- Bohnert, H.J.; Jensen, R.G. Strategies for engineering water-stress tolerance in plants. Trends Biotechnol. 1996, 14, 89–97. [Google Scholar] [CrossRef]
- Ashraf, M.; Foolad, M.R. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot. 2007, 59, 206–216. [Google Scholar] [CrossRef]
- Umar, J.; Aliyu, A.; Shehu, K.; Abubakar, L. Influence of Salt Stress on Proline and Glycine Betaine Accumulation in Tomato (Solanum lycopersicum L.). J. Hort. Plant 2018, 1, 19–25. [Google Scholar] [CrossRef]
- Yildirima, E.; Ekincia, M.; Turanb, M.; Dursuna, A.; Kula, R.; Parlakova, F. Roles of glycine betaine in mitigating deleterious effect of salt stress on lettuce (Lactuca sativa L.). Arch. Agron. Soil Sci. 2015, 61, 1673–1689. [Google Scholar] [CrossRef]
- Park, E.J.; Jeknic, Z.; Pino, M.T.; Murata, N.; Chen, T.H. Glycinebetaine accumulation is more effective in chloroplasts than in the cytosol for protecting transgenic tomato plants against abiotic stress. Plant Cell Environ. 2007, 30, 994–1005. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Zhang, W.; Gao, Q.; Yang, A.; Hu, X.; Zhang, J. Enhancement of drought resistance and biomass by increasing the amount of glycine betaine in wheat seedlings. Euphytica 2011, 177, 151–167. [Google Scholar] [CrossRef]
- Sakamoto, A.; Murata, N. The role of glycine betaine in the protection of plants from stress: Clues from transgenic plants. Plant Cell Environ. 2002, 25, 163–171. [Google Scholar] [CrossRef]
- Rasheed, R.; Iqbal, M.; Ashraf, M.A.; Hussain, I.; Shafiq, F.; Yousaf, A.; Zaheer, A. Glycine betaine counteracts the inhibitory effects of waterlogging on growth, photosynthetic pigments, oxidative defence system, nutrient composition, and fruit quality in tomato. J. Hortic. Sci. Biotechnol. 2017, 93, 385–391. [Google Scholar] [CrossRef]
- Hisyam, B.; Alam, M.A.; Naimah, N.; Jahan, M.S. Roles of Glycinebetaine on Antioxidants and Gene Function in Rice Plants Under Water Stress. Asian J. Plant Sci. 2017, 16, 132–140. [Google Scholar]
- Yadu, S.; Dewangan, T.L.; Chandrakar, V.; Keshavkant, S. Imperative roles of salicylic acid and nitric oxide in improving salinity tolerance in Pisum sativum L. Physiol. Mol. Biol. Plants 2017, 23, 43–58. [Google Scholar] [CrossRef] [Green Version]
- Yao, W.Q.; Lei, Y.K.; Yang, P.; Li, Q.S.; Wang, L.L.; He, B.Y.; Xu, Z.M.; Zhou, C.; Ye, H.J. Exogenous Glycinebetaine Promotes Soil Cadmium Uptake by Edible Amaranth Grown during Subtropical Hot Season. Int. J. Environ. Res. Public Health 2018, 15, 1794. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.H.; Murata, N. Glycinebetaine: An effective protectant against abiotic stress in plants. Trends Plant Sci. 2008, 13, 499–505. [Google Scholar] [CrossRef]
- Hanson, A.D.; Scott, N.A. Betaine synthesis from radioactive precursors in attached, water-stressed barley leaves. Plant Physiol. 1980, 66, 342–348. [Google Scholar] [CrossRef] [Green Version]
- Tian, F.; Wang, W.; Liang, C.; Wang, X.; Wang, G.; Wang, W. Over accumulation of glycine betaine makes the function of the thylakoid membrane better in wheat under salt stress. Crop J. 2017, 5, 73–82. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Khan, S.; Khan, M.A.; Aamir, M.; Ullah, H.J.; Nawab, J.; Rehman, I.U.; Shah, J. Heavy metals effects on plant growth and dietary intake of trace metals in vegetables cultivated in contaminated soil. Int. J. Environ. Sci. Technol. 2018, 16, 2295–2304. [Google Scholar] [CrossRef]
- Gupta, N.; Thind, S.K.; Bains, N.S. Glycine betaine application modifies biochemical attributes of osmotic adjustment in drought-stressed wheat. Plant Growth Regul. 2013, 72, 221–228. [Google Scholar] [CrossRef]
- Yang, N.; Wang, C.L.; He, W.P.; Qu, Y.Z.; Li, Y.S. Photosynthetic characteristics and effects of exogenous glycine of Chorispora bungeana under drought stress. Photosynthetica 2016, 54, 459–467. [Google Scholar] [CrossRef]
- Liu, N.; Lin, S.; Huang, B. Differential Effects of Glycine Betaine and Spermidine on Osmotic Adjustment and Antioxidant Defense Contributing to Improved Drought Tolerance in Creeping Bentgrass. J. Am. Soc. Hortic. Sci. 2017, 142, 20–26. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.; Chaudhary, A.; Rizwan, M.; Anwar, H.T.; Adrees, M.; Farid, M.; Irshad, M.K.; Hayat, T.; Anjum, S.A. Alleviation of chromium toxicity by glycinebetaine is related to elevated antioxidant enzymes and suppressed chromium uptake and oxidative stress in wheat (Triticum aestivum L.). Environ. Sci. Pollut. Res. 2015, 22, 10669–10678. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.A.; Ali, S.; Hameed, A.; Bharwana, S.A.; Rizwan, M.; Ishaque, W.; Farid, M.; Mahmood, K.; Iqbal, Z. Cadmium stress in cotton seedlings: Physiological, photosynthesis andoxidative damages alleviated by glycinebetaine. S. Afr. J. Bot. 2016, 104, 61–68. [Google Scholar] [CrossRef]
- Jabeen, N.; Abbas, Z.; Iqbal, M.; Rizwan, M.; Jabbar, A.; Farid, M.; Ali, S.; Ibrahim, M.; Abbas, F. Glycinebetaine mediates chromium tolerance in mung bean through lowering of Cr uptake and improved antioxidant system. Arch. Agron. Soil Sci. 2016, 62, 648–662. [Google Scholar] [CrossRef]
- Xalxo, R.; Yadu, B.; Chakraborty, P.; Chandrakar, V.; Keshavkant, S. Modulation of nickel toxicity by glycinebetaine and aspirin in Pennisetum typhoideum. Acta Biol. Szeged. 2017, 61, 163–171. [Google Scholar]
- Yao, W.; Xu, T.; Farooq, S.U.; Jin, P.; Zheng, Y. Glycine betaine treatment alleviates chilling injury in zucchini fruit (Cucurbita pepo L.) by modulating antioxidant enzymes and membrane fatty acid metabolism. Postharvest Biol. Technol. 2018, 144, 20–28. [Google Scholar] [CrossRef]
- Nusrat, N.; Shahbaz, M.; Perveen, S. Modulation in growth, photosynthetic efficiency, activity of antioxidants and mineral ions by foliar application of glycine betaine on pea (Pisum sativum L.) under salt stress. Acta Physiol. Plant 2014, 36, 2985–2998. [Google Scholar] [CrossRef]
- Ahanger, M.A.; Tyagi, S.R.; Wani, M.R.; Ahmad, P. Drought tolerance: Roles of organic osmolytes, growth regulators and mineral nutrients. In Physiological Mechanisms and Adaptation Strategies in Plants Under Changing Environment; Ahmad, P., Wani, M.R., Eds.; Springer: New York, NY, USA, 2014; Volume 1, pp. 25–56. [Google Scholar]
- Ahanger, M.A.; Gul, F.; Ahmad, P.; Akram, N.A. Environmental Stresses and Metabolomics—Deciphering the Role of Stress Responsive Metabolites. In Plant Metabolites and Regulation Under Environmental Stress; Elsevier Inc.: Amsterdam, The Netherlands, 2018; pp. 53–67. [Google Scholar]
- Fan, W.; Zhang, M.; Zhang, H.; Zhang, P. Improved Tolerance to Various Abiotic Stresses in Transgenic Sweet Potato (Ipomoea batatas) Expressing Spinach Betaine Aldehyde Dehydrogenase. PLoS ONE 2012, 7, e37344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.; Yan, K.; Shao, H.; Zhao, S. Physiological Mechanisms for High Salt Tolerance in Wild Soybean (Glycine soja) from Yellow River Delta, China: Photosynthesis, Osmotic Regulation, Ion Flux and antioxidant Capacity. PLoS ONE 2013, 8, e83227. [Google Scholar] [CrossRef]
- Malekzadeh, P. Inflence of exogenous application of glycine betaine on antioxidative system and growth of salt-stressed soybean seedlings (Glycine max L.). Physiol. Mol. Biol. Plants 2015, 21, 225–232. [Google Scholar] [CrossRef] [Green Version]
- Akram, N.A.; Ashraf, M.; Al-Qurainy, F. Aminolevulinic acidinduced changes in some key physiological attributes and activities of antioxidant enzymes in sunflwer (Helianthus annuus L.) plants under saline regimes. Sci. Hortic. 2012, 142, 143–148. [Google Scholar] [CrossRef]
- Noctor, G.; Mhamdi, A.; Foyer, C.H. The roles of reactive oxygen metabolism in drought: Not so cut and dried. Plant Physiol. 2014, 164, 1636–1648. [Google Scholar] [CrossRef] [Green Version]
- You, J.; Chan, Z.L. ROS regulation during abiotic stress responses in crop plants. Front. Plant Sci. 2015, 6, 1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, M.I.R.; Iqbal, N.; Masood, A.; Mobin, M.; Anjum, N.A.; Khan, N.A. Modulation and significance of nitrogen and sulfur metabolism in cadmium challenged plants. Plant Growth Regul. 2016, 78, 1–11. [Google Scholar] [CrossRef]
- Lou, Y.; Yang, Y.; Hu, L.; Liu, H.; Xu, Q. Exogenous glycinebetaine alleviates the detrimental effect of Cd stress on perennial ryegrass. Ecotoxicology 2015, 24, 1330–1340. [Google Scholar] [CrossRef]
- Ali, Q.; Anwar, F.; Ashraf, M.; Saari, N.; Perveen, R. Ameliorating effects of exogenously applied proline on seed composition, seed oil quality and oil antioxidant activity of maize (Zea mays L.) under drought stress. Int. J. Mol. Sci. 2013, 14, 818–835. [Google Scholar] [CrossRef]
- Hossain, M.A.; Hasanuzzaman, M.; Fujita, M. Up-regulation of antioxidant and glyoxalase systems by exogenous glycine betaine and proline in mung bean confer tolerance to cadmium stress. Physiol. Mol. Biol. Plants 2010, 16, 259–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bharwana, S.A.; Ali, S.; Farooq, M.A.; Iqbal, N.; Hameed, A.; Abbas, F.; Ahmad, M.S.A. Glycine betaine-induced lead toxicity tolerance related to elevated photosynthesis, antioxidant enzymes suppressed lead uptake and oxidative stress in cotton. Turk. J. Bot. 2014, 38, 281–292. [Google Scholar] [CrossRef]
- Islam, M.M.; Anamul-Hoque, M.; Okuma, E.; Banu, N.A.; Shimoishi, Y.; Nakamura, Y.; Murata, Y. Exogenous proline and glycinebetaine increase antioxidant enzyme activities and confer tolerance to cadmium stress in cultured tobacco cells. J. Plant Physiol. 2009, 166, 1587–1597. [Google Scholar] [CrossRef]
- Molla, M.R.; Ali, M.R.; Hasanuzzaman, M.; Al-Mamun, M.H.; Ahmed, A.; Nazim-ud-Dowla, M.A.N.; Rohman, M.M. Exogenous proline and betaine-induced upregulation of glutathione transferase and glyoxalase I in lentil (Lens culinaris) under drought stress. Not. Bot. Horti. Agrobot. Cluj Napoca 2014, 42, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Raza, M.A.S.; Saleem, M.F.; Moazzam, J.M.; Khan, I. Impact of foliar applied glycinebetaine on growth and Physiology of wheat (Triticum Aestivum L.) Under drought. Pak. J. Agric. Sci. 2014, 51, 327–334. [Google Scholar]
- Cha-um, S.; Samphumphuang, T.; Kirdmanee, C. Glycinebetaine alleviates water deficit stress in indica rice using proline accumulation, photosynthetic efficiencies, growth performances and yield attributes. Aust. J. Crop Sci. 2013, 7, 213–218. [Google Scholar]
- Masindi, V.; Muedi, K. Environmental contamination by heavy Metals. In Heavy Metals; Hosam El-Din, M., Saleh Refaat Aglan, F., Eds.; IntechOpen: Rijeka, Croatia, 2018. [Google Scholar] [CrossRef] [Green Version]
- Ruciniska-Sobkowiak, R. Oxidative stress in plants exposed to heavy metals. Postepy Biochemii 2010, 56, 191–200. [Google Scholar]
- Shahid, M.; Pourrut, B.; Dumat, C.; Nadeem, M.; Aslam, M.; Pinelli, E. Heavy-metal-induced reactive oxygen species: Phytotoxicity and physicochemical changes in plants. Rev. Environ. Contamin. Toxicol. 2014, 232, 1–44. [Google Scholar]
- Fariduddin, Q.; Varshney, P.; Yousuf, M.; Ali, A.; Ahmad, A. Dissecting the role of Glycine Betaine in Plants under Abiotic Stress. Plant Stress. 2013, 7, 8–18. [Google Scholar]
- Hasan, M.K.; Cheng, Y.; Kanwar, M.K.; Chu, X.Y.; Ahammed, G.J.; Qi, Z.Y. Responses of plant proteins to heavy metal stress—A review. Front. Plant Sci. 2017, 8, 1492. [Google Scholar] [CrossRef] [Green Version]
- Zouari, M.; Elloumi, N.; Labrousse, P.; Ben Rouina, B.; Ben Abdallah, F.; Ben Ahmed, C. Olive trees response to lead stress: Exogenous proline provided better tolerance than glycine betaine. S. Afr. J. Bot. 2018, 118, 158–165. [Google Scholar] [CrossRef]
- Duman, F.; Aksoy, A.; Aydin, Z.; Temizgul, R. Effects of Exogenous Glycinebetaine and Trehalose on Cadmium Accumulation and Biological Responses of an Aquatic Plant (Lemna gibba L.). Water Air Soil Pollut. 2011, 217, 545–556. [Google Scholar] [CrossRef]
- Rasheed, R.; Ashraf, M.A.; Hussain, I.; Haider, M.Z.; Kanwal, U.; Iqbal, M. Exogenous proline and glycinebetaine mitigate cadmium stress in two genetically different spring wheat (Triticum aestivum L.) cultivars. Braz. J. Bot. 2014, 37, 399–406. [Google Scholar] [CrossRef]
- Stepien, P.; Gediga, K.; Piszcz, U.; Karmowska, K. Effects of the exogenous glycine betaine on photosynthetic apparatus in cucumber leaves challenging Al stress. In Proceedings of the 18th International Conference on Heavy Metals in the Environment, Ghent, Belgium, 12–15 October 2016. [Google Scholar]
- Dubeya, A.K.; Kumara, N.; Ranjana, R.; Gautam, A.; Pande, V.; Sanyal, I.; Mallick, S. Application of glycine reduces arsenic accumulation and toxicity in Oryza sativa L. by reducing the expression of silicon transporter genes. Ecotoxicol. Environ. Saf. 2018, 148, 410–417. [Google Scholar] [CrossRef]
- Kumar, P.; Tokas, J.; Singal, H.R. Amelioration of Chromium VI Toxicity in Sorghum (Sorghum bicolor L.) using Glycine Betaine. Sci. Rep. 2019, 9, 16020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.; Richmond, M.E.; Williams, D.V.; Zheng, W.; Wu, F. Exogenous Glycine betaine reduces cadmium uptake and mitigates cadmium toxicity in two tobacco genotypes differing in cadmium tolerance. Int. J. Mol. Sci. 2019, 20, 1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad, R.; Ali, S.; Abid, M.; Rizwan, M.; Ali, B.; Tanveer, A.; Ghani, M.A. Glycinebetaine alleviates the chromium toxicity in Brassica oleracea L. by suppressing oxidative stress and modulating the plant morphology and photosynthetic attributes. Environ. Sci. Pollut. Res. 2020, 27, 1101–1111. [Google Scholar] [CrossRef]
- Kurepin, L.V.; Ivanov, A.G.; Zaman, M.; Pharis, R.P.; Hurry, V.; Hüner, N.P. Interaction of glycine betaine and plant hormones: Protection of the photosynthetic apparatus during abiotic stress. In Photosynthesis: Structures, Mechanisms, and Applications; Springer: Cham, Switzerland, 2017; pp. 185–202. [Google Scholar]
- Anjum, S.A.; Farooq, M.; Wang, L.C.; Xue, L.L.; Wang, S.G.; Wang, L.; Chen, M. Gas exchange and chlorophyll synthesis of maize cultivars are enhanced by exogenously-applied glycinebetaine under drought conditions. Plant Soil Environ. 2011, 57, 326–331. [Google Scholar] [CrossRef] [Green Version]
- Bhatti, K.H.; Anwar, S.; Nawaz, K.; Hussain, K.; Siddiqi, E.H.; Sharif, R.U.; Talat, A.; Khalid, A. Effect of Exogenous Application of Glycinebetaine on Wheat (Triticum aestivum L.) Under Heavy Metal Stress. Middle-East J. Sci. Res. 2013, 14, 130–137. [Google Scholar]
- Bai, X.Y.; Dong, Y.J.; Wang, Q.H.; Xu, L.L.; Kong, J.; Liu, S. Effects of lead and nitric oxide on photosynthesis, antioxidative ability, and mineral element content of perennial ryegrass. Biol. Plantarum 2015, 59, 163–170. [Google Scholar] [CrossRef]
- Mahdavian, K.; Ghaderian, S.M.; Schat, H. Pb accumulation, Pb tolerance, antioxidants, thiols, and organic acids in metallicolous and non-metallicolous Peganum harmala L. under Pb exposure. Environ. Exp. Bot. 2016, 126, 21–31. [Google Scholar] [CrossRef]
- Cuypers, A.; Smeets, K.; Vangronsveld, J. Heavy Metal Stress in Plants. In Plant Stress Biology: From Genomics to Systems Biology; Hirt, H., Ed.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2009. [Google Scholar]
- Pourrut, B.; Pohu, A.L.; Pruvot, C.; Garçon, G.; Verdin, A.; Waterlot, C.; Bidar, G.; Shirali, P.; Douay, F. Assessment of flash-aided phytostabilisation of highly contaminated soils after an 8-year field trial Part 2. Influence on plants. Sci. Total Environ. 2011, 409, 4504–4510. [Google Scholar] [CrossRef] [PubMed]
- Sheetal, K.R.; Singh, S.D.; Anand, A.; Prasad, S. Heavy metal accumulation and effects on growth, biomass and physiological processes in mustard. Ind. J. Plant Physiol. 2016, 21, 219–223. [Google Scholar] [CrossRef]
- Chen, F.; Gao, J.; Zhou, Q. Toxicity assessment of simulated urban runoff containing polycyclic musks and cadmium in Carassius auratus using oxidative stress biomarkers. Environ. Pollut. 2012, 162, 91–99. [Google Scholar] [CrossRef] [PubMed]
- Farmer, E.E.; Mueller, M.J. ROS-Mediated lipid peroxidation and RES-activated signaling. Annu. Rev. Plant Biol. 2013, 64, 429–450. [Google Scholar] [CrossRef]
- Genard, H.; LeSaos, J.; Hillard, J.; Tremolieres, A.; Boucaud, J. Effect of salinity on lipid composition, glycine betaine content and photosynthetic activity in chloroplasts of Suaeda maritime. Plant Physiol. Biochem. 1991, 29, 421–427. [Google Scholar]
- Iqbal, N.; Ashraf, M.; Ashraf, M.Y. Glycinebetaine, an osmolyte of interest to improve water stress tolerance in sunflower (Helianthus annuus L.): Water relations and yield. S. Afr. J. Bot. 2008, 74, 274–281. [Google Scholar] [CrossRef] [Green Version]
- Ibrahim, A.H.; Aldesuquy, H.S. Glycine Betaine and Shikimic Acid—Induced Modification in Growth Criteria, Water Relation and Productivity of Droughted Sorghum bicolor Plants. Phyton Horn 2003, 43, 351–361. [Google Scholar]
- Nomura, M.; Hibino, T.; Takabe, T.; Sugiyama, T.; Yokota, A.; Miyake, H.; Takabe, T. Transgenically produced glycinebetaine protects ribulose-1, 5-biophosphate carboxylase/oxygenase from inactivation of Synechococcus sp. PCC7942 under salt stress. Plant Cell Physiol. 1998, 32, 425–432. [Google Scholar] [CrossRef] [Green Version]
- Feigl, G.; Lehotai, N.; Molnár, Á.; Ördög, A.; Rodríguez-Ruiz, M.; Palma, J.M.; Corpas, F.J.; Erdei, L.; Kolbert, Z. Zinc induces distinct changes in the metabolism of reactive oxygen and nitrogen species (ROS and RNS) in the roots of two Brassica species with different sensitivity to zinc stress. Ann. Bot. 2015, 116, 613–625. [Google Scholar] [CrossRef] [Green Version]
- Das, K.; Roychoudhury, A. Reactive oxygen species (ROS) and response of antioxidants as ROS-scavengers during environmental stress in plants. Front. Environ. Sci. 2014, 2, 53. [Google Scholar] [CrossRef] [Green Version]
- Maleki, M.; Ghorbanpour, M.; Kariman, K. Physiological and antioxidative responses of medicinal plants exposed to heavy metals stress. Plant Gene 2017, 11, 247–254. [Google Scholar] [CrossRef]
- Miller, G.; Suzuki, N.; Ciftci-Yilmaz, S.; Ron, M. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010, 33, 453–467. [Google Scholar] [CrossRef]
- Srivastava, S.; Dubey, R.S. Manganese-excess induces oxidative stress, lowers the pool of antioxidants and elevates activities of key antioxidative enzymes in rice seedlings. Plant Growth Regul. 2011, 64, 1–16. [Google Scholar] [CrossRef]
- Luis, A. ROS and RNS in plant physiology: An overview. J. Exp. Bot. 2015, 66, 2827–2837. [Google Scholar]
- Rehman, M.Z.; Rizwan, M.; Ghafoor, A.; Naeem, A.; Ali, S.; Sabir, M.; Qayyum, M.F. Effect of inorganic amendments for in situ stabilization of cadmium in contaminated soil and its phyto-availability to wheat and rice under rotation. Environ. Sci. Pollut. Res. 2015, 22, 16897–16906. [Google Scholar] [CrossRef] [PubMed]
- Giri, J. Glycinebetaine and abiotic stress tolerance in plants. Plant Signal. Behav. 2011, 6, 1746–1751. [Google Scholar] [CrossRef] [PubMed]
- Shahbaz, M.; Zia, B. Does exogenous application of glycinebetaine through rooting medium alter rice (Oryza sativa L.) mineral nutrient status under saline conditions. J. Appl. Bot. Food Qual. 2012, 84, 54–60. [Google Scholar]
- Cao, F.; Liu, L.; Ibrahim, W.; Cai, Y.; Wu, F. Alleviating effects of exogenous glutathione, glycinebetaine, brassinosteroids and salicylic acid on cadmium toxicity in rice seedlings (Oryza Sativa). Agrotechnology 2013, 2, 107–112. [Google Scholar] [CrossRef] [Green Version]
- Mahmood, T.; Ashraf, M.; Shahbaz, M. Does exogenous application of Glycine betaine as a Pre sowing seed treatment improves growth and regulate some key physiological attributes in wheat plants grown under water deficit conditions. Pak. J. Bot. 2009, 41, 1291–1302. [Google Scholar]
- Tripathi, D.K.; Singh, S.; Singh, S.; Mishra, S.; Chauhan, D.K.; Dubey, N.K. Micronutrients and their diverse role in agricultural crops: Advances and future prospective. Acta Physiol. Plant. 2015, 37, 1–14. [Google Scholar] [CrossRef]
- Hu, R.; Sunc, K.; Suc, X.; Pana, Y.; Zhang, Y. Physiological responses and tolerance mechanisms to Pb in two xerophils: Salsola passerina Bunge and Chenopodium album L. J. Hazard. Mater. 2012, 205–206, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Shi, J.B.; Wang, X.; Meng, M.; Huang, L.; Qi, XL.; He, B.; Ye, Z.H. Variations and constancy of mercury and methylmercury accumulation in rice grown at contaminated paddy field sites in three Provinces of China. Environ. Pollut. 2013, 181, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Yonny, M.E.; Rodríguez Torressi, A.; Nazareno, M.A.; Cerutti, S. Development of a novel, sensitive, selective, and fast methodology to determine malondialdehyde in leaves of melon plants by ultra-high-performance liquid chromatography-tandem mass spectrometry. J. Anal. Methods Chem. 2017, 9. [Google Scholar] [CrossRef]
- Patel, A.; Pandey, V.; Patra, D.D. Metal absorption properties of Mentha spicata grown under tannery sludge amended soil—Its effect on antioxidant system and quality. Chemosphere 2016, 147, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.A.; Ali, S.; Hameed, A.; Ishaque, W.; Mahmood, K.; Iqbal, Z. Alleviation of cadmium toxicity by silicon is related to elevated photosynthesis, antioxidant enzymes; suppressed cadmium uptake and oxidative stress in cotton. Ecotoxicol. Environ. Saf. 2013, 96, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Ali, B.; Qian, P.; Jin, R.; Ali, S.; Khan, M.; Aziz, R.; Tian, T.; Zhou, W. Physiological and ultra-structural changes in Brassica napus seedlings induced by cadmium stress. Biol. Plant 2014, 58, 131–138. [Google Scholar] [CrossRef]
- Zaheer, I.E.; Ali, S.; Rizwan, M.; Bareen, F.E.; Abbas, Z.; Bukhari, S.A.H.; Wijaya, L.; Alyemeni, M.N.; Ahmad, P. Zinc-lysine prevents chromium-induced morphological, photosynthetic, and oxidative alterations in spinach irrigated with tannery wastewater. Environ. Sci. Pollut. Res. 2019, 26, 28951–28961. [Google Scholar] [CrossRef]
- Najeeb, U.; Jilani, G.; Ali, S.; Sarwar, M.; Xu, L.; Zhou, W. Insights into cadmium induced physiological and ultra-structural disorders in Juncus effusus L, and its remediation through exogenous citric acid. J. Hazard. Mater. 2011, 186, 565–574. [Google Scholar] [CrossRef]
- Saidi, I.; Ayouni, M.; Dhieb, A.; Chtourou, Y.; Chaïbi, W.; Djebali, W. Oxidative damages induced by short-term exposure to cadmium in bean plants: Protective role of salicylic acid. S. Afr. J. Bot. 2013, 85, 32–38. [Google Scholar] [CrossRef] [Green Version]
- Ehsan, S.; Ali, S.; Noureen, S.; Mahmood, K.; Farid, M.; Ishaque, W.; Shakoor, M.B.; Rizwan, M. Citric acid assisted phytoremediation of cadmium by Brassica napus L. Ecotoxicol. Environ. Saf. 2014, 106, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Zeng, G.; Huang, D.; Dong, H.; Lai, C.; Chen, M.; Tang, W.; Li, F.; Leng, Y.; Cheng, M.; et al. Cadmium induced hydrogen peroxide accumulation and responses of enzymatic antioxidants in Phanerochaete chrysosporium. Ecol. Eng. 2015, 75, 110–115. [Google Scholar] [CrossRef]
- Al Hassan, M.; Fuertes, M.M.; Ramos Sánchez, F.J.; Vicente, O.; Boscaiu, M. Effects of Salt and Water Stress on Plant Growth and on Accumulation of Osmolytes and Antioxidant Compounds in Cherry Tomato. Not BotHortiAgrobo 2015, 43, 1–11. [Google Scholar]
- Kito, K.; Yamamori, T.; Theerawitaya, C.; Cha-um, S.; Fukaya, M.; Rai, V.; Takabe, T. Differential Effects of Excess Potassium and Sodium on Plant Growth and Betaine Accumulation in Sugar Beet. J. Agron. Agric. Asp. 2017, 3, 1–7. [Google Scholar]
- Sozharajan, R.; Natarajan, S. NaCl stress causes changes in photosynthetic pigments and accumulation of compatible solutes in Zea mays L. J Appl Advan Res. 2016, 1, 3–9. [Google Scholar] [CrossRef] [Green Version]
- De la Torre-González, A.; Montesinos-pereira, D.; Blasco, B.; Ruiz, J.M. Influence of the proline metabolism and glycine betaine on tolerance to salt stress in tomato (Solanum lycopersicum L.) commercial genotypes. J. Plant Physiol. 2018, 231, 329–336. [Google Scholar] [CrossRef]
- Rasheed, R.; Wahid, A.; Ashraf, M.; Basra, S.M.A. Role of proline and glycinebetaine in improving chilling stress tolerance in sugarcane buds at sprouting. Int. J. Agric. Biol. 2010, 12, 1–8. [Google Scholar]
- Jin, P.; Zhang, Y.; Shan, T.; Huang, Y.; Xu, J.; Zheng, Y. Low-Temperature Conditioning Alleviates Chilling Injury in Loquat Fruit and Regulates Glycine Betaine Content and Energy Status. J. Agric. Food Chem. 2015, 63, 3654–3659. [Google Scholar] [CrossRef]
- Wang, Q.; Ding, T.; Zuo, J.; Gao, L.; Fan, L. Amelioration of postharvest chilling injury in sweet pepper by glycine betaine. Postharvest Biol. Technol. 2016, 112, 114–120. [Google Scholar] [CrossRef]
- Ahmad, R.; Hussain, J.; Jamil, M.; Kim, M.D.; Kwak, S.S.; Shah, M.M.; El-Hendawy, S.E.; Al-Suhaibani, N.A.; Rehman, S. Glycinebetaine Synthesizing Transgenic Potato Plants Exhibit Enhanced Tolerance To Salt And Cold Stresses. Pak. J. Bot. 2014, 46, 1987–1993. [Google Scholar]
- Burgess, P.; Huang, B. Effects of sequential application of plant growth regulators and osmoregulants on drought tolerance of creeping bentgrass. Crop Sci. 2013, 54, 837–844. [Google Scholar] [CrossRef]
- Liu, X.M.; Kim, K.E.; Kim, K.C.; Nguyen, X.C.; Han, H.J.; Jung, M.S.; Kim, H.S.; Kim, S.H.; Park, H.C.; Yun, D.J.; et al. Cadmium activates Arabidopsis MPK3 and MPK6 via accumulation of reactive oxygen species. Phytochemistry 2010, 71, 614–618. [Google Scholar] [CrossRef] [PubMed]
- Wei, D.; Zhang, W.; Wang, C.; Meng, Q.; Li, G.; Chen, T.H.H.; Yang, X. Genetic engineering of the biosynthesis of glycinebetaine leads to alleviate salt-induced potassium efflux and enhances salt tolerance in tomato plants. Plant Sci. 2017, 257, 74–83. [Google Scholar] [CrossRef] [PubMed]
- Rady, M.O.A.; Semida, W.M.; Abd El-Mageed, T.A.; Hemida, K.A.; Rady, M.M. Up-regulation of antioxidative defense systems by glycine betaine foliar application in onion plants confer tolerance to salinity stress. Sci. Hortic. 2018, 240, 614–622. [Google Scholar] [CrossRef]
- Kumar, A.; Dharamvir Kumar, M.; Jangra, M.; Kumar, N. Growth and Development of Rapeseed Mustard and Other Field Crops under Different Sowing Dates. Int. J. Pure Appl. Biosci. 2018, 6, 144–156. [Google Scholar] [CrossRef]
- Sabagh, A.E.L.; Sorour, S.; Ragab, A.; Saneoka, H.; Islam, M.S. The Effect of Exogenous Application of Proline and Glycine Betaineon the Nodule Activity of Soybean under Saline Condition. J. Agric. Biotechnol. 2017, 2, 1–5. [Google Scholar]
- Yao, T.C.; Shaharuddin, N.A.; Ling, H.C.; Mahmood, M. Exogenous Application of Glycine Betaine Alleviates Salt Induced Damages More Efficiently Than Ascorbic Acid In In Vitro Rice Shoots. Aust. J. Basic Appl. Sci. 2016, 10, 58–65. [Google Scholar]
- Alasvandyari, F.; Mahdavi, B. Effect of Glycinebetaine On Growth and Antioxidant Enzymes of Safflower Under Salinity Stress Condition. Agric. For. 2017, 63, 85–95. [Google Scholar] [CrossRef]
- Shams, M.; Yildirim, E.; Ekinci, M.; Turan, M.; Dursun, A.; Parlakova, F.; Kul, R. Exogenously applied glycine betaine regulates some chemical characteristics and antioxidative defence systems in lettuce under salt stress. Hortic. Environ. Biotechnol. 2016, 57, 225–231. [Google Scholar] [CrossRef]
- Xing, W.; Rajashekar, C.B. Glycinebetaine involvement in freezing tolerance and water stress is Arabidopsis thaliana. Environ. Exp. Bot. 2001, 46, 21–28. [Google Scholar] [CrossRef]
- Hussain, H.A.; Men, S.; Hussain, S.; Chen, Y.; Ali, S.; Zhang, S.; Zhang, K.; Li, Y.; Xu, Q.; Liao, C.; et al. Interactive effects of drought and heat stresses on morpho-physiological attributes, yield, nutrient uptake and oxidative status in maize hybrids. Sci. Rep. 2019, 9, 3890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashraf, M.; Harris, J.C. Potential biochemical indicators of salinity tolerance in plants. Plant Sci. 2004, 166, 3–16. [Google Scholar] [CrossRef]
- Somersalo, S.; Kyei-Boahen, S.; Pehu, E. Exogenous glycine betaine application as a possibility to increase low temperature tolerance of crop plants. Nordisk Jordbruksforsk 1996, 78, 10. [Google Scholar]
- Chen, W.P.; Li, P.H.; Chen, T.H.H. Glycine betaine increases chilling tolerance and reduces chilling-induced lipid peroxidation in Zea mays L. Plant Cell Environ. 2000, 23, 609–618. [Google Scholar] [CrossRef]
- Nayyar, H.; Chander, K.; Kumar, S.; Bains, T. Glycine betaine mitigates cold stress damage in Chickpea. Agron. Sustain. Dev. 2005, 25, 381–388. [Google Scholar] [CrossRef]
- Cheng, C.; Pei, L.M.; Yin, T.T.; Zhang, K.W. Seed treatment with glycine betaine enhances tolerance of cotton to chilling stress. J. Agric. Sci. 2018, 156, 323–332. [Google Scholar] [CrossRef]
- Wang, L.; Shan, T.; Xie, B.; Ling, C.; Shao, S.; Jin, P.; Zheng, Y. Glycine betaine reduces chilling injury in peach fruit by enhancing phenolic and sugar metabolisms. Food Chem. 2019, 272, 530–538. [Google Scholar] [CrossRef]
- Jagendorf, T.A.; Takabe, T. Inducers of glycine betaine synthesis in barley. Plant Physiol. 2001, 127, 1827–1835. [Google Scholar] [CrossRef]
- Di Martino, C.; Delfine, S.; Pizzuto, R.; Loreto, F.; Fuggi, A. Free amino acids and glycine betaine in leaf osmoregulation of spinach responding to increasing salt stress. New Phytol. 2003, 158, 455–463. [Google Scholar] [CrossRef]
- Sulpice, R.; Tsukaya, H.; Nonaka, H.; Mustardy, L.; Chen, T.H.; Murata, N. Enhanced formation of flowers in salt-stressed Arabidopsis after genetic engineering of the synthesis of glycine betaine. Plant J. 2003, 36, 165–176. [Google Scholar] [CrossRef]
- Nuccio, M.L.; McNeil, S.D.; Ziemak, M.J.; Hanson, A.D.; Jain, R.K.; Selvaraj, G. Choline Import into Chloroplasts Limits Glycine Betaine Synthesis in Tobacco: Analysis of Plants Engineered with a Chloroplastic or a Cytosolic Pathway. Metab. Eng. 2000, 2, 300–311. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Li, S.; Jiang, T.; Liu, Z.; Zhang, W.; Jian, G.; Qi, F. Chilling stress—The key predisposing factor for causing Alternaria alternata infection and leading to cotton (Gossypium hirsutum L.) leaf senescence. PLoS ONE 2012, 7, e36126. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Rue, K. The effect of glycine betaine priming on seed germination of six turfgrass species under drought, salinity, or temperature stress. HortScience 2014, 49, 1454–1460. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, N.; Zhang, Y.; Yu, H.; Gabar, A.; Zhou, Y.; Li, Z.; Zhang, M. Seed priming with Glycine betaine improve seed germination characteristics and antioxidant capacity of wheat (Triticum aestivum L.) seedlings under water-stress conditions. Appl. Ecol. Environ. Res. 2019, 17, 8333–8350. [Google Scholar] [CrossRef]
- Grumet, R.; Isleib, G.T.; Hanson, A.D. Genetic Control of Glycinebetaine Level in Barley. Crop Sci. 1985, 25, 618–622. [Google Scholar] [CrossRef]
- Iqbal, N.; Ashraf, M.Y. Does Seed Treatment with Glycinebetaine Improve Germination Rate and Seedling Growth of Sunflower (Helianthus Annuus L.) Under Osmotic Stress. Pak. J. Bot. 2006, 38, 1641–1648. [Google Scholar]
- Naidu, B.P. Production of betaine from Australian Melaleuca spp. for use in agriculture to reduce plant stress. Aust. J. Exp. Agric. 2003, 43, 1163–1170. [Google Scholar] [CrossRef]
- Demir, I.; Oztokat, C. Effect of salt priming on germination and seedling growth at low temperatures in watermelon seeds during development. Seed Sci. Technol. 2003, 31, 765–770. [Google Scholar] [CrossRef]
- Farooq, M.; Basra, S.M.A.; Ahmad, N. Improving the performance of transplanted rice by seed priming. Plant Growth Regul. 2007, 51, 129–137. [Google Scholar] [CrossRef]
- Allakhverdiev, S.I.; Los, D.A.; Mohanty, P.; Nishiyama, Y.; Murata, N. Glycinebetaine alleviates the inhibitory effectof moderate heat stress on the repair of photosystem II during photoinhibition. Biochim. Biophys. Acta 2007, 1767, 1363–1371. [Google Scholar] [CrossRef] [Green Version]
- Hassanein, R.A.; Hassanein, A.A.; Haider, A.S.; Hashem, H.A. Improving salt tolerance of Zea mays L. plants by presoaking their grains in glycine betaine. Aust. J. Basic Appl. Sci. 2009, 3, 928–942. [Google Scholar]
- Korkmaz, A.; Sirikci, R.; Kocacınar, F.; Özlem Deger, O.; Demirkırıan, A.R. Alleviation of salt-induced adverse effects in pepper seedlings by seed application of glycinebetaine. Sci. Hortic. 2012, 148, 197–205. [Google Scholar] [CrossRef]
- Arafa, A.A.; Khafagy, M.A.; El-Banna, M.F. The effect of glycinebetaine or ascorbic acid on grain germination and leaf structure of sorghum plants grown under salinity stress. Aust. J. Crop Sci. 2009, 3, 294–304. [Google Scholar]
- Heuer, B. Influence of exogenous application of proline and glycinebetaine on growth of salt-stressed tomato plants. Plant Sci. 2003, 165, 693–699. [Google Scholar] [CrossRef]
- Corol, D.; Ravel, C.; Rakszegi, M.; Bedo, Z.; Charmet, G.; Beale, M.E. Effects of genotype and environment on the contents of betaine, choline, and trigonelline in cereal grains. J. Agric. Food Chem. 2012, 60, 5471–5481. [Google Scholar] [CrossRef] [PubMed]
- Moharramnejad, S.; Sofalian, O.; Valizadeh, M.; Asgari, A.; Shiri, M. Proline, glycine betaine, total phenolics and pigment con-tents in response to osmotic stress in maize seedlings. J. Biosci. Biotechnol. 2015, 4, 313–319. [Google Scholar]
- Lerma, C.; Roch, P.J.; Ju, G.C.; Yang, W.J.; Hanson, A.D.; Rhodes, D. Betaine deficiency in maize: Complementation tests and metabolic basis. Plant Physiol. 1991, 95, 1113–1119. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.J.; Nadolska-Orczyk, A.; Wood, K.V.; Hahn, D.T.; Rich, P.J.; Wood, A.J.; Saneoka, H.; Premachandra, G.S.; Bonham, C.C.; Rhodes, J.C. Near-Isogenic Lines of Maize Differing for Glycinebetaine. Plant Physiol. 1995, 107, 621–630. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Liang, J.; Wang, M.; Li, D.; Liu, Y.; Chen, T.H.; Yang, X. Genetic engineering of the biosynthesis of glycine betaine enhances the fruit development and size of tomato. Plant Sci. 2019, 280, 355–366. [Google Scholar] [CrossRef]
- Munns, R.; Tester, M. Mechanisms of salinity tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681. [Google Scholar] [CrossRef] [Green Version]
- Ashraf, M.; Akram, N.A.; Arteca, R.N.; Foolad, M.R. The physiological, biochemical and molecular roles of brassinosteroids and salicylic acid in plant processes and salt tolerance. Crit. Rev. Plant Sci. 2010, 29, 162–190. [Google Scholar] [CrossRef]
- Zhang, H.; Dong, H.; Li, W.; Sun, Y.; Chen, S.; Kong, X. Increased glycine betaine synthesis and salinity tolerance in ahcmo transgenic cotton lines. Mol. Breed. 2009, 23, 289–298. [Google Scholar] [CrossRef]
- Kishitani, S.; Takanami, T.; Suzuki, M.; Oikawa, M.; Yokoi, S.; Ishitani, M. Compatibility of glycinebetaine in rice plants: Evaluation using transgenic rice plants with a gene for peroxisomal betaine aldehyde dehydrogenase from barley. Plant Cell Environ. 2000, 23, 107–114. [Google Scholar] [CrossRef]
- Khan, M.S.; Yu, X.; Kikuchi, A.; Asahina, M.; Watanabe, K.N. Genetic engineering of glycine betaine biosynthesis to enhance abiotic stress tolerance in plants. Plant Biotechnol. 2009, 26, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Qin, D.; Zhao, C.L.; Liu, X.Y.; Wang, P.W. Transgenic soybeans expressing betaine aldehyde dehydrogenase from Atriplex canescens show increased drought tolerance. Plant Breed. 2017, 136, 699–709. [Google Scholar] [CrossRef]
- Hashemi, F.S.G.; Ismail, M.R.; Rafii, M.Y.; Aslani, F.; Miah, G.; Muharam, F.M. Critical multifunctional role of the betaine aldehyde dehydrogenase gene in plants. Biotechnol. Equip. 2018, 32, 818–829. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plant Species | Heavy Metal Stress | Effect of Exogenous Glycine Betaine (GB) | References |
|---|---|---|---|
| Wheat | Cr Stress | Glycine Betaine (GB) improved chlorophyll contents, biomass, growth characteristics and protein content. | [81] |
| Cotton | Cd Stress | Glycine Betaine (GB) boosted the plant growth, improved activities of antioxidant enzyme and rate of photosynthesis | [82] |
| Mung Bean | Cr Stress | Glycine Betaine (GB) improved plant growth. | [83] |
| Amaranth | Cd Stress | Glycine Betaine (GB) significantly encouraged the rate of photosynthesis in edible amaranth and considerably improved the chlorophyll content of leaves. | [89] |
| Perennial Ryegrass | Cd Stress | Glycine Betaine (GB) improved stability of cell membrane via decreasing lipid membrane oxidation. | [97] |
| Cotton | Pb Stress | Effectively improved the gas attributes and plant growth under Pb stress. | [99] |
| Wheat | Cd Stress | Glycine Betaine (GB) improved fresh biomass of roots and shoots. | [111] |
| Cucumber | Al Stress | Glycine Betaine (GB) showed significant protective effect on chlorophyll content. | [112] |
| Asian Rice | As Stress | Glycine Betaine (GB) increased the GST and GRX gene expression alongside As stress. | [113] |
| Sorghum | Cr Stress | Glycine Betaine (GB) improved the quality and total yield of sorghum. | [114] |
| Tobacco | Cd Stress | Glycine Betaine (GB) reduces the stomatal closure, accumulation of malondialdehyde (MDA) and damage to leaf. | [115] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, S.; Abbas, Z.; Seleiman, M.F.; Rizwan, M.; YAVAŞ, İ.; Alhammad, B.A.; Shami, A.; Hasanuzzaman, M.; Kalderis, D. Glycine Betaine Accumulation, Significance and Interests for Heavy Metal Tolerance in Plants. Plants 2020, 9, 896. https://doi.org/10.3390/plants9070896
Ali S, Abbas Z, Seleiman MF, Rizwan M, YAVAŞ İ, Alhammad BA, Shami A, Hasanuzzaman M, Kalderis D. Glycine Betaine Accumulation, Significance and Interests for Heavy Metal Tolerance in Plants. Plants. 2020; 9(7):896. https://doi.org/10.3390/plants9070896
Chicago/Turabian StyleAli, Shafaqat, Zohaib Abbas, Mahmoud F. Seleiman, Muhammad Rizwan, İlkay YAVAŞ, Bushra Ahmed Alhammad, Ashwag Shami, Mirza Hasanuzzaman, and Dimitris Kalderis. 2020. "Glycine Betaine Accumulation, Significance and Interests for Heavy Metal Tolerance in Plants" Plants 9, no. 7: 896. https://doi.org/10.3390/plants9070896