Exploring Mechanisms of Quantitative Resistance to Leptosphaeria maculans (Blackleg) in the Cotyledons of Canola (Brassica napus) Based on Transcriptomic and Microscopic Analyses

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
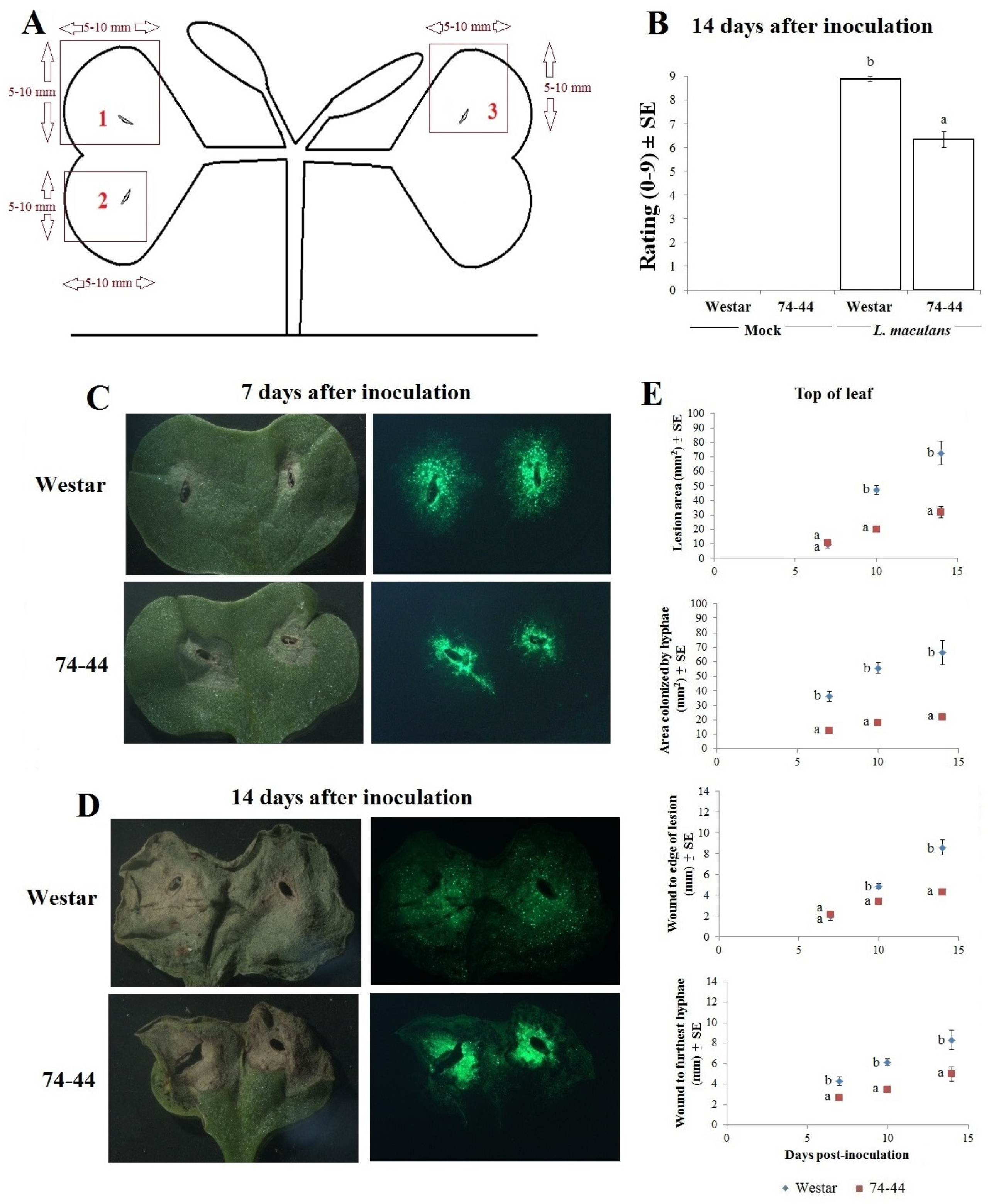
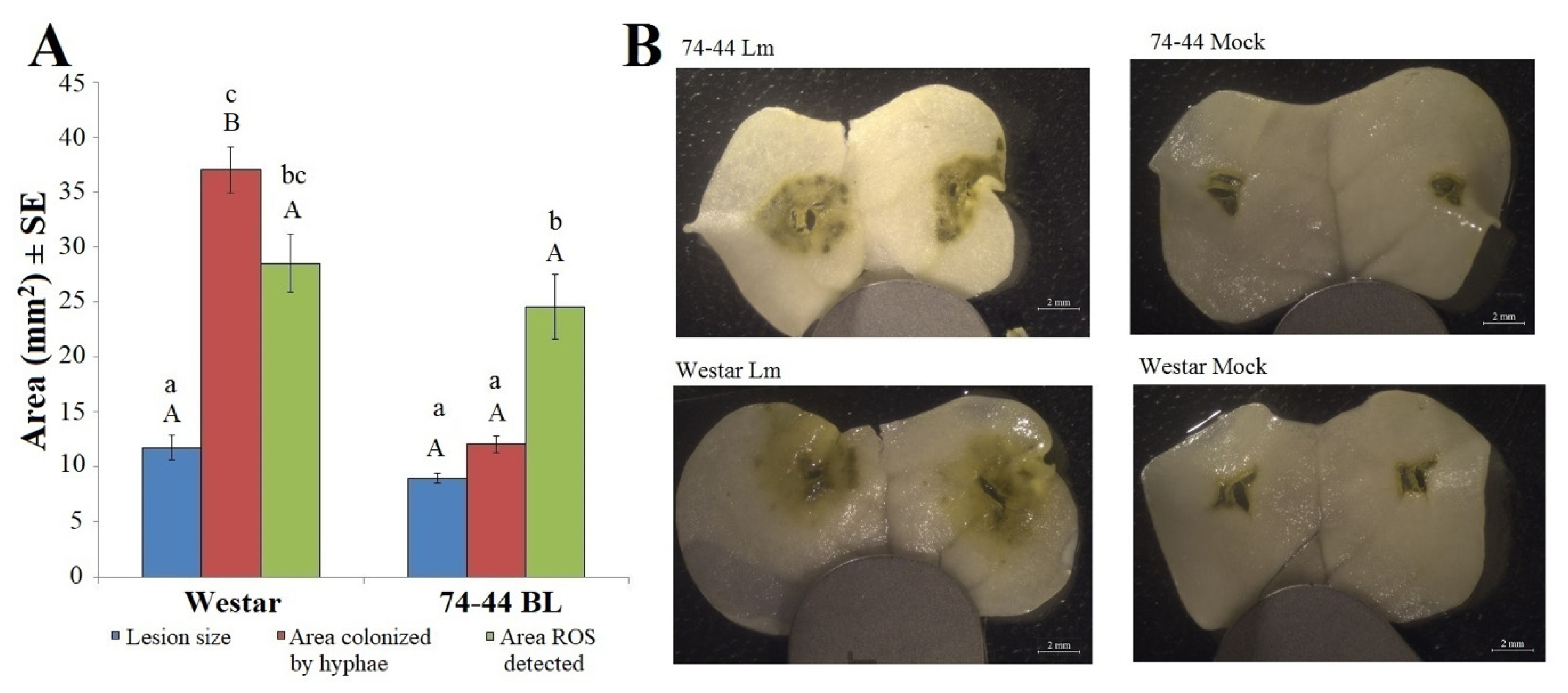
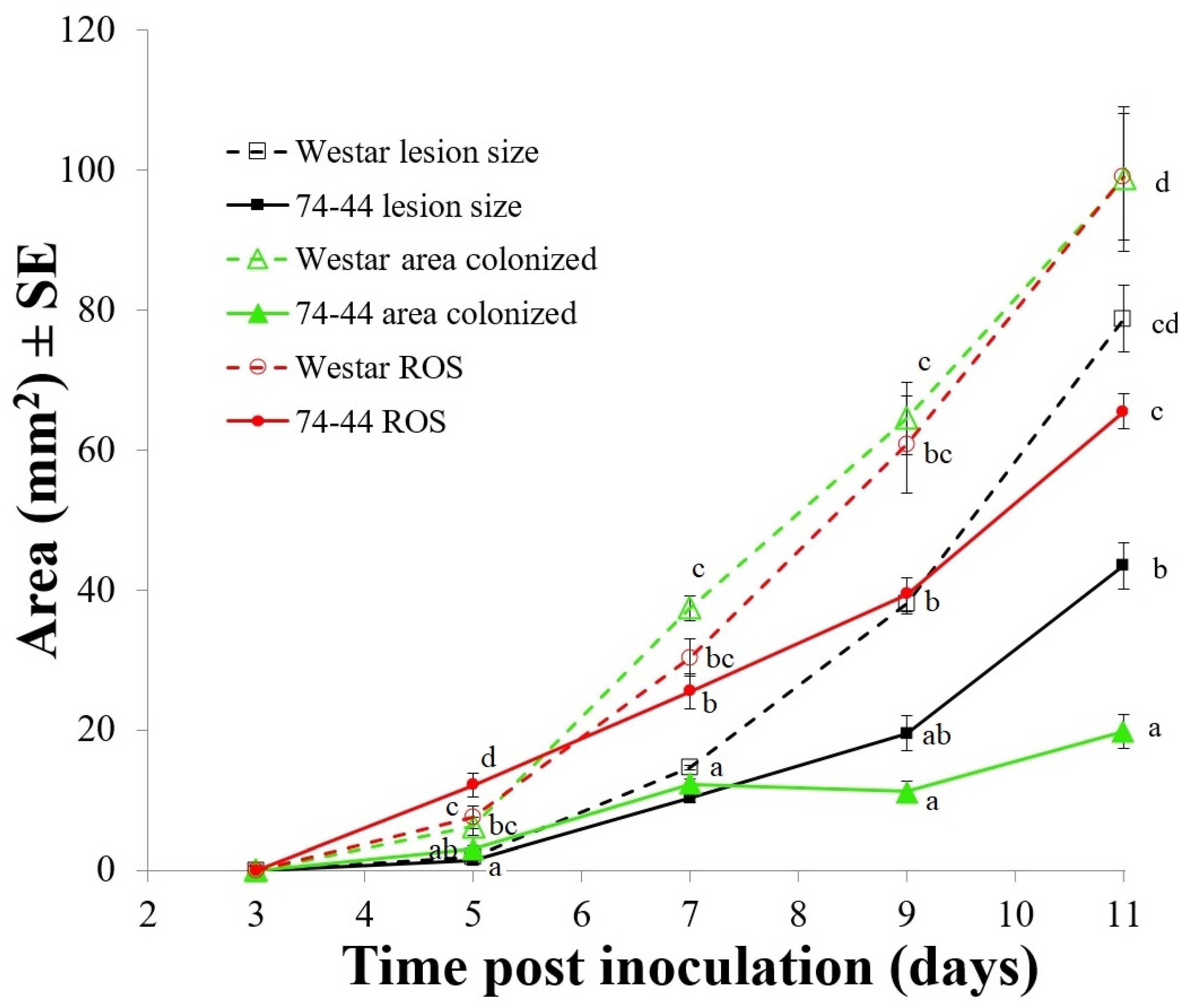
2.1. Infection Symptoms and Lm Hyphal Growth in the Cotyledons of Susceptible and QR Canola Cultivars
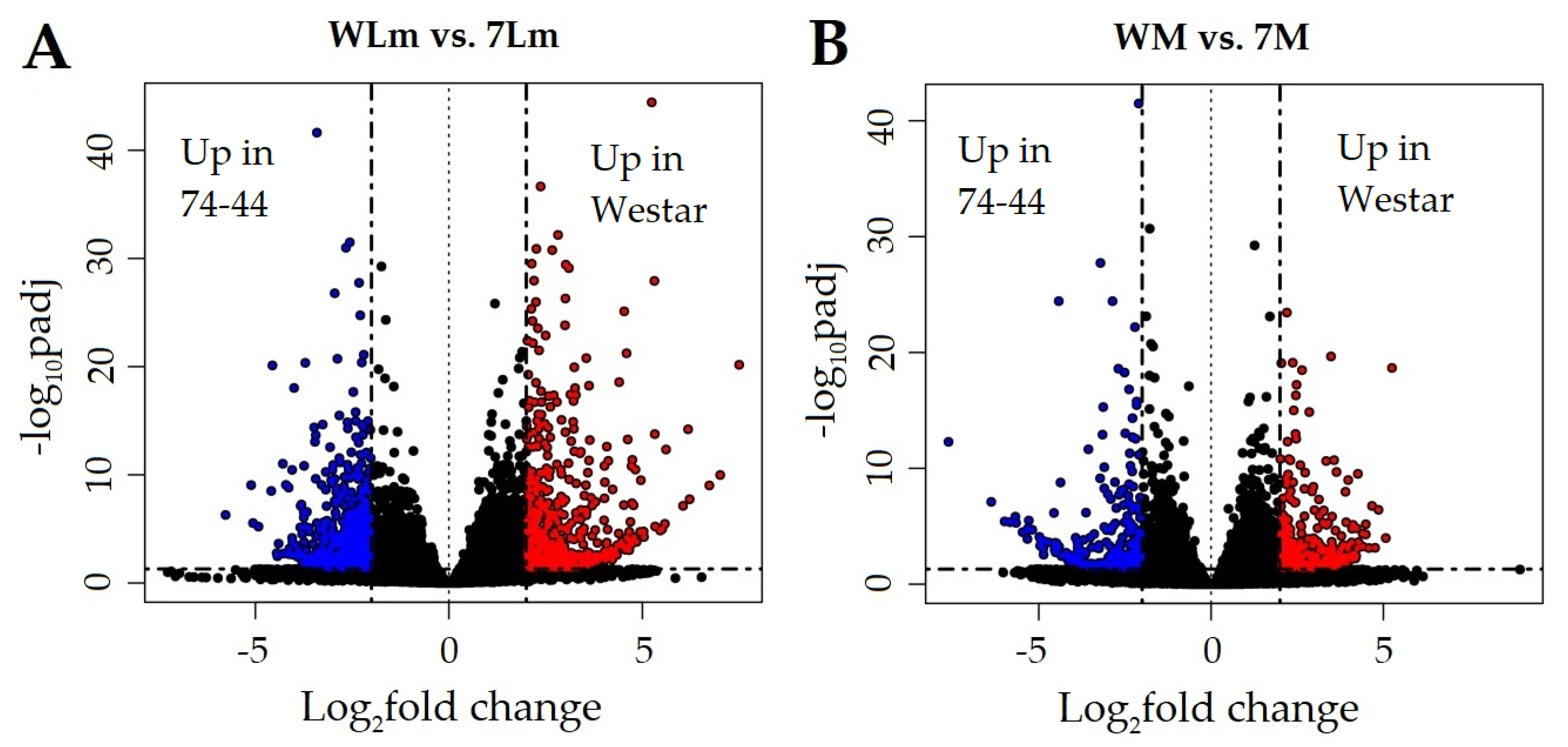
2.2. RNA-Seq Analyses
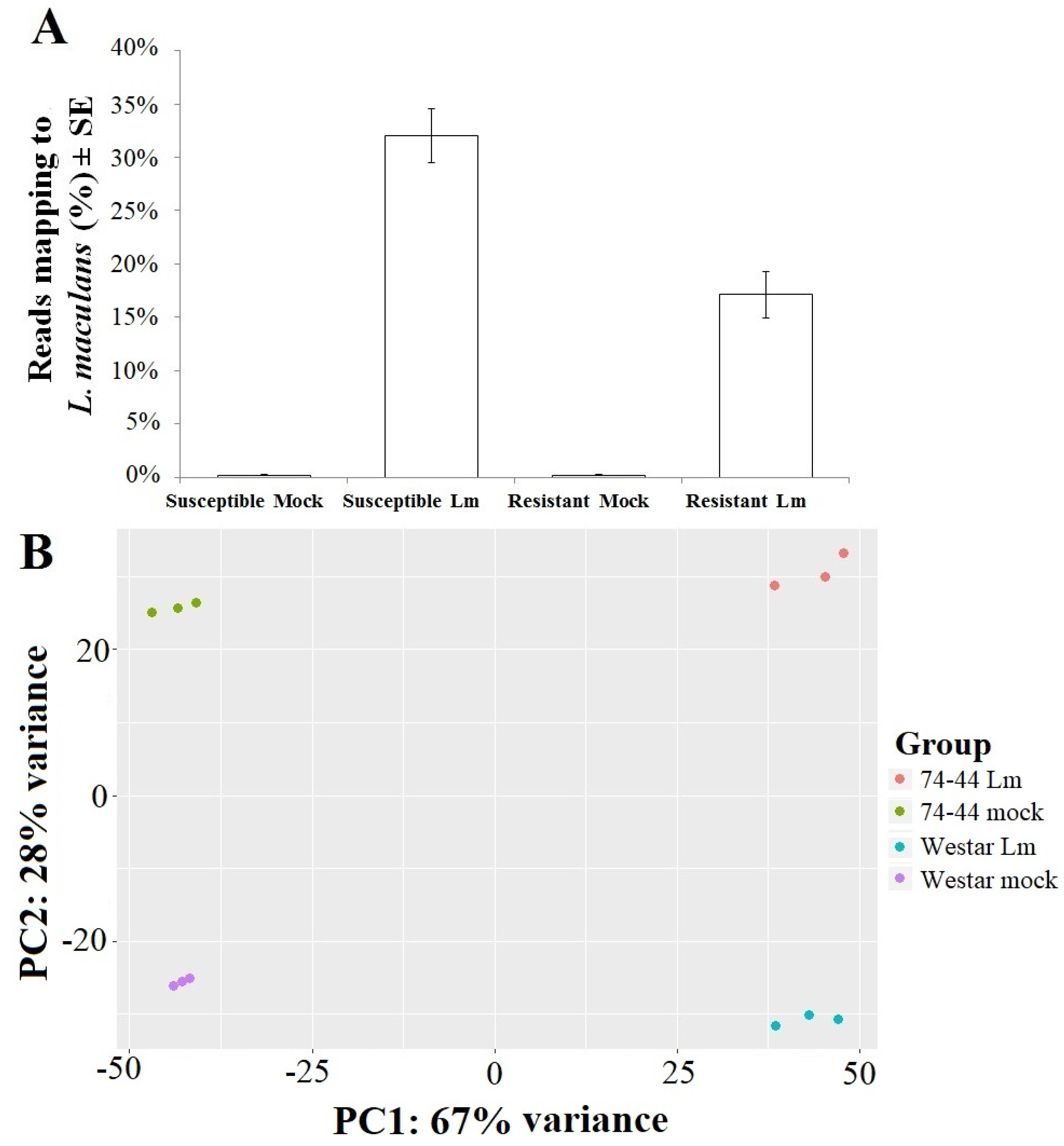
2.2.1. Expression of L. maculans Genes in Inoculated B. napus
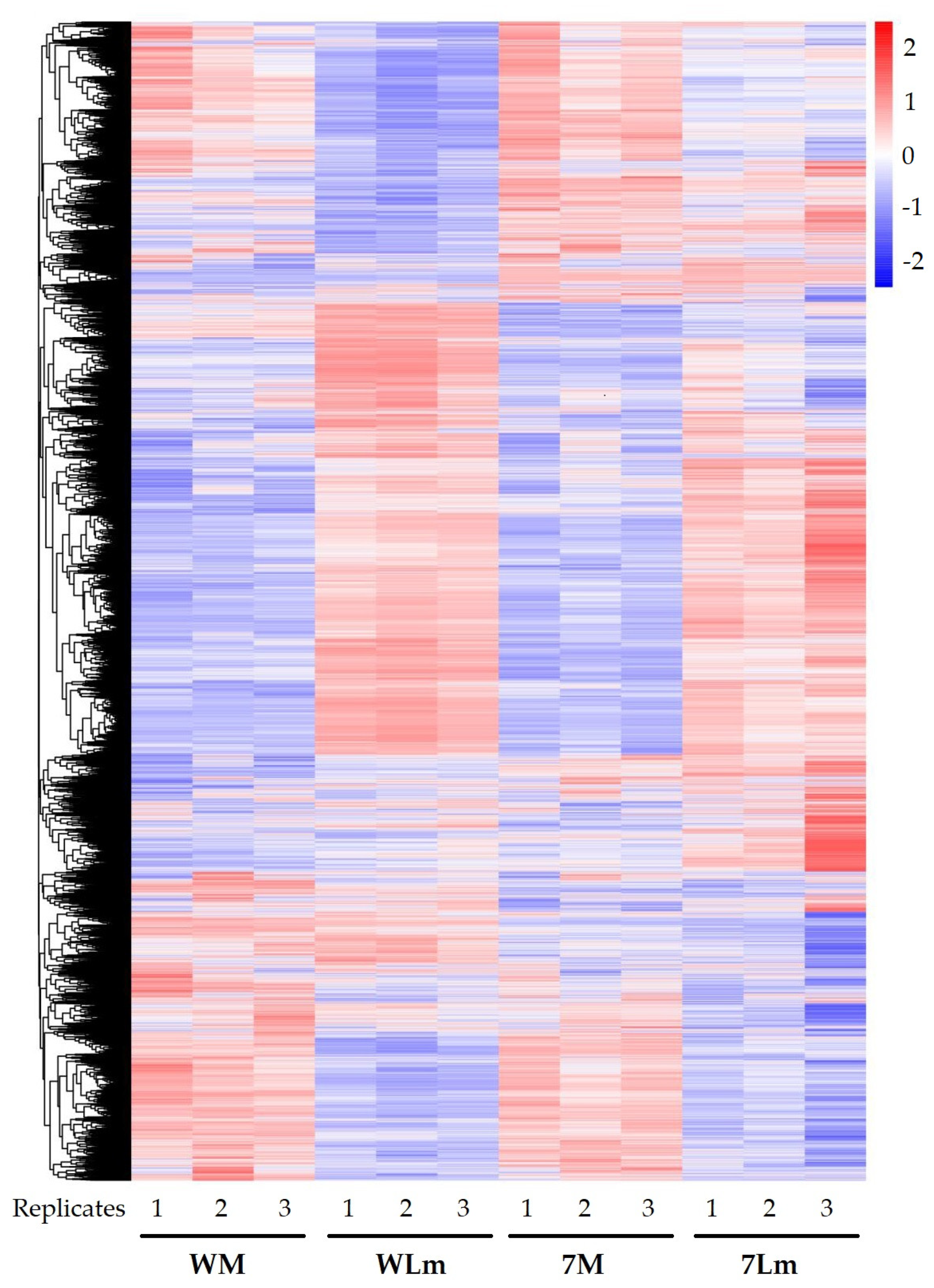
2.2.2. Expression of the B. napus Genes
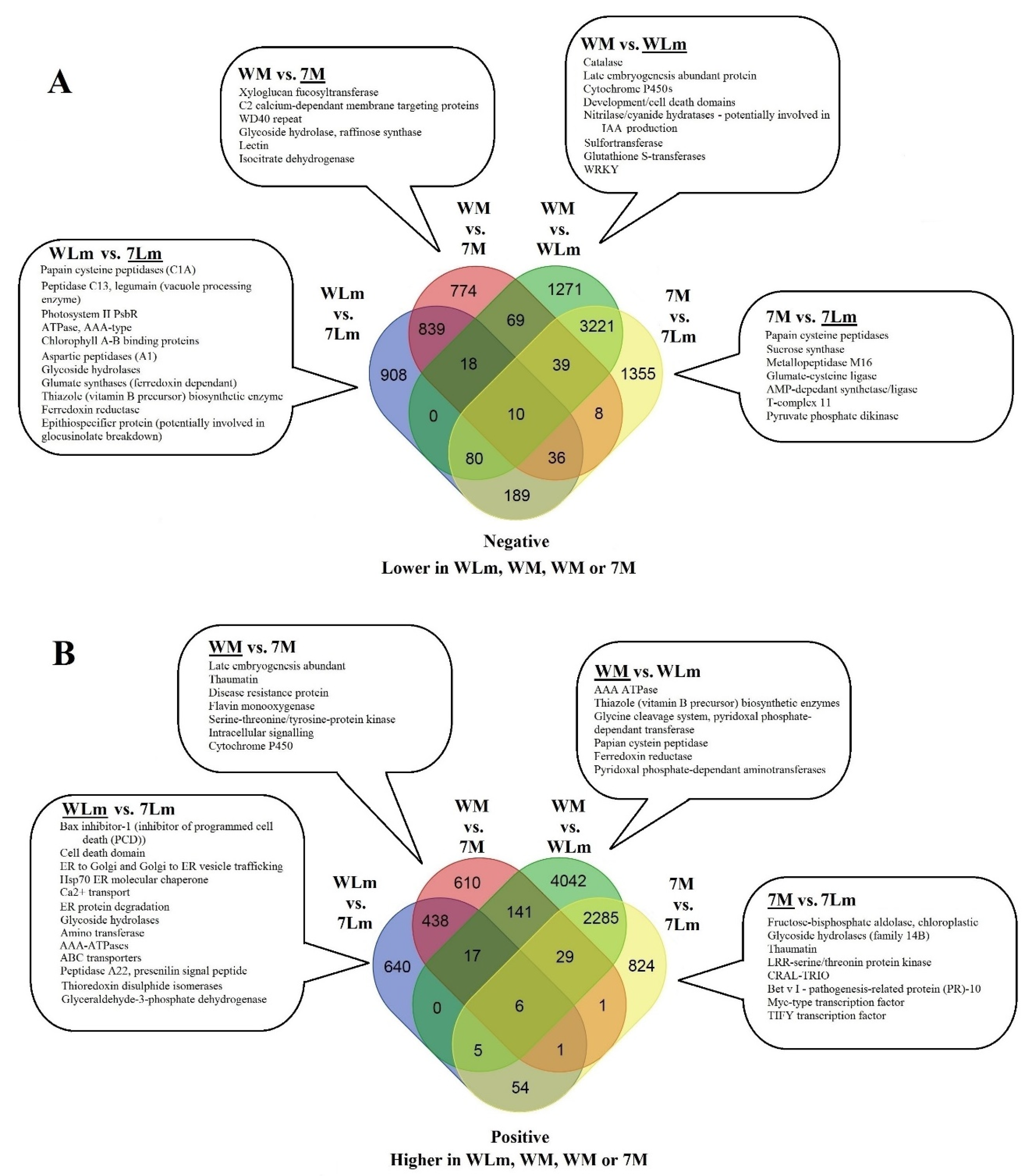
Upregulated B. napus Genes in Inoculated 74-44BL
2.2.3. Upregulated B. napus Genes in Inoculated Westar
2.2.4. Upregulated B. napus Genes in Mock-Inoculated 74-44 BL
2.2.5. Upregulated B. napus Genes in Mock-Inoculated Westar
2.3. HP Production in Cotyledons
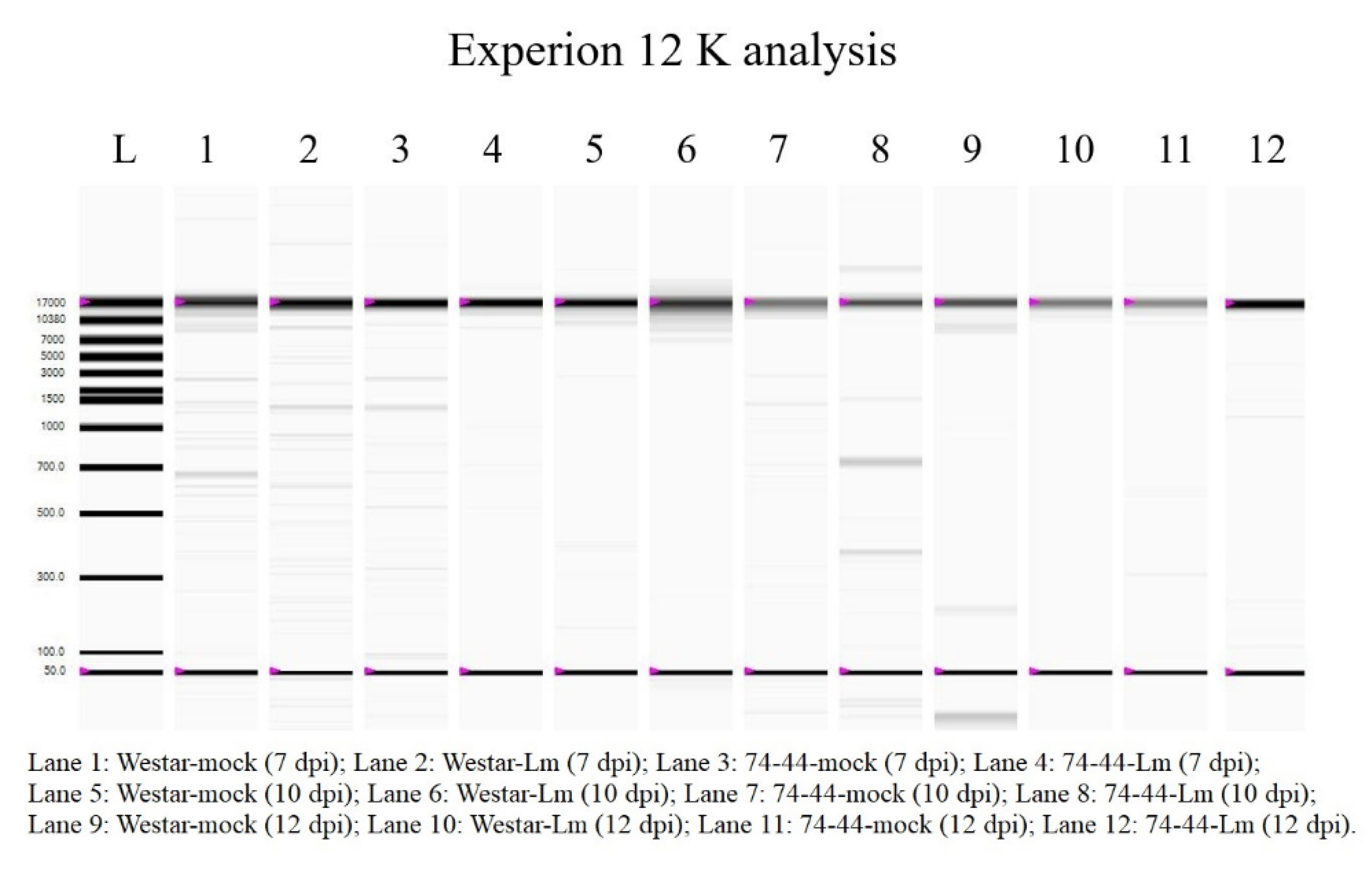
2.4. Genomic DNA Degradation as an Indicator of PCD
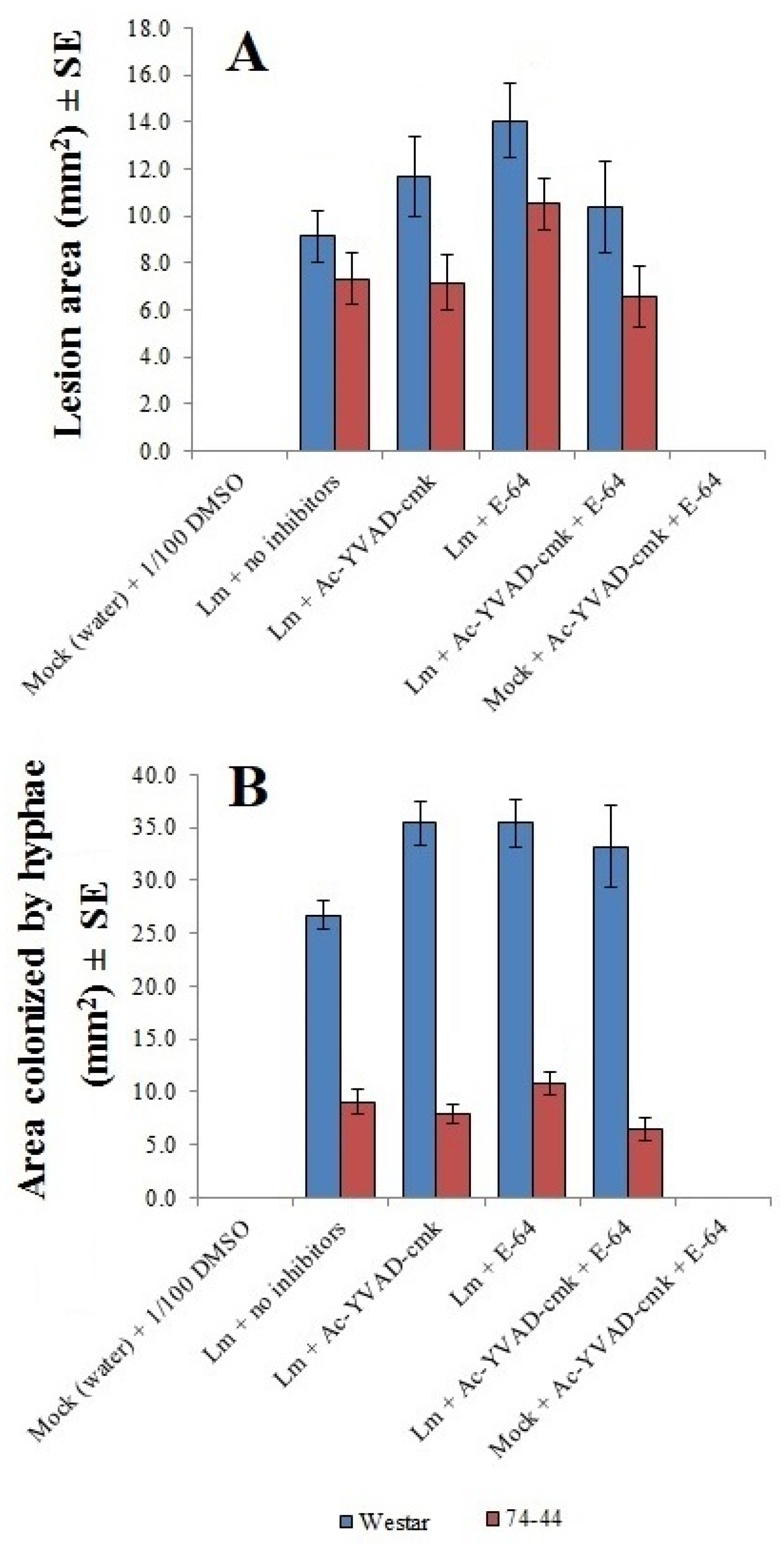
2.5. Impact of Protease Inhibitors on Lm Infection of Cotyledons
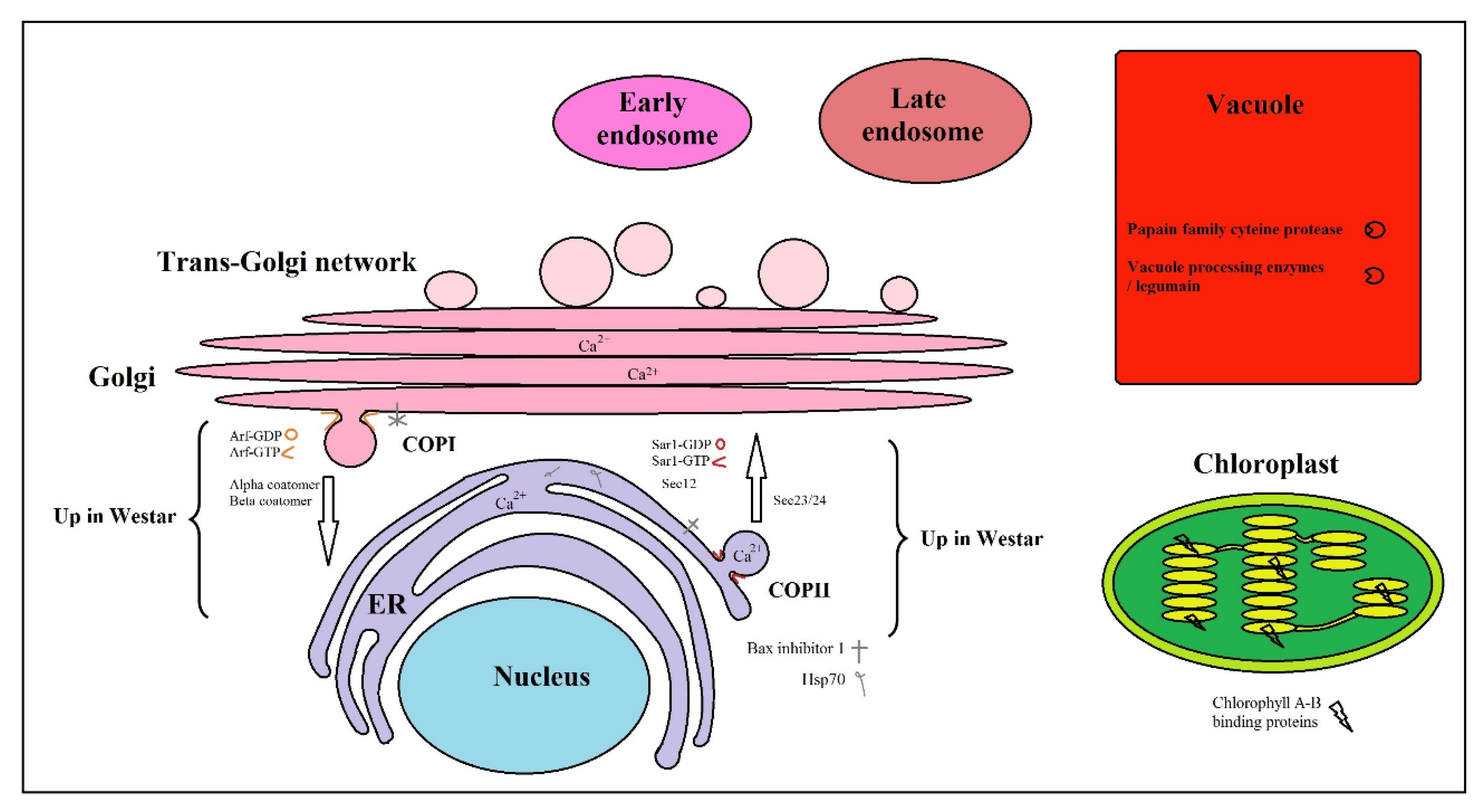
3. Discussion
4. Materials and Methods
4.1. Fungal and Plant Material
4.2. RNA Extraction, Library Preparation, and Sequencing
4.3. RNA-Seq Data Analysis
4.4. Colorimetric Detection of Hydrogen Peroxide
4.5. Time-Series Infection in Cotyledons
4.6. Assessment of Genomic DNA Degradation as a Marker of Programmed Cell Death
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Fitt, B.D.L.; Brun, H.; Barbetti, M.J.; Rimmer, S.R. World-Wide Importance of Phoma Stem Canker (Leptosphaeria maculans and L. biglobosa) on Oilseed Rape (Brassica napus). Eur. J. Plant Pathol. 2006, 114, 3–15. [Google Scholar] [CrossRef]
- Delourme, R.; Chevre, A.-M.; Brun, H.; Rouxel, T.; Balesdent, M.H.; Dias, J.S.; Salisbury, P.; Renard, M.; Rimmer, S.R. Major Gene and Polygenic Resistance to Leptosphaeria maculans in Oilseed Rape (Brassica napus). Eur. J. Plant Pathol. 2006, 114, 41–52. [Google Scholar] [CrossRef]
- Cowger, C.; Brown, J.K.M. Durability of Quantitative Resistance in Crops: Greater Than We Know? Annu. Rev. Phytopathol. 2019, 57, 253–277. [Google Scholar] [CrossRef] [PubMed]
- Larkan, N.; Lydiate, D.J.; Parkin, I.A.P.; Nelson, M.N.; Epp, D.J.; Rimmer, S.R.; Cowling, W.A.; Borhan, H. The Brassica napus blackleg resistance gene lepr3 encodes a receptor-like protein triggered by the Leptosphaeria maculans effector AVRLM1. New Phytol. 2012, 197, 595–605. [Google Scholar] [CrossRef] [PubMed]
- Yu, F.; Lydiate, D.J.; Rimmer, S.R. Identification of two novel genes for blackleg resistance in Brassica napus. Theor. Appl. Genet. 2005, 110, 969–979. [Google Scholar] [CrossRef]
- Yu, F.; Gugel, R.K.; Kutcher, H.R.; Peng, G.; Rimmer, S.R. Identification and mapping of a novel blackleg resistance locus LepR4 in the progenies from Brassica napus × B. rapa subsp. Sylvestris. Theor. Appl. Genet. 2012, 126, 307–315. [Google Scholar] [CrossRef] [PubMed]
- Raman, R.; Taylor, B.; Lindbeck, K.; Coombes, N.; Barbulescu, D.; Salisbury, P.; Raman, H. Molecular and Molecular mapping and validation of Rlm1 gene for resistance to Leptosphaeria maculans in canola (Brassica napus L.). Crop. Pasture Sci. 2012, 63, 1007. [Google Scholar] [CrossRef]
- Larkan, N.; Lydiate, D.J.; Yu, F.; Rimmer, S.R.; Borhan, H. Co-localisation of the blackleg resistance genes Rlm2 and LepR3 on Brassica napus chromosome A10. BMC Plant Biol. 2014, 14, 387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parlange, F.; Daverdin, G.; Fudal, I.; Kuhn, M.-L.; Balesdent, M.-H.; Blaise, F.; Grezes-Besset, B.; Rouxel, T. Leptosphaeria maculans avirulence gene AvrLm4-7 confers a dual recognition specificity by theRlm4andRlm7resistance genes of oilseed rape, and circumventsRlm4-mediated recognition through a single amino acid change. Mol. Microbiol. 2009, 71, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.; Paillard, S.; Fopa-Fomeju, B.; Falentin, C.; Deniot, G.; Baron, C.; Vallée, P.; Manzanares-Dauleux, M.J.; Delourme, R. Multi-year linkage and association mapping confirm the high number of genomic regions involved in oilseed rape quantitative resistance to blackleg. Theor. Appl. Genet. 2018, 131, 1627–1643. [Google Scholar] [CrossRef]
- Raman, H.; Raman, R.; Diffey, S.; Qiu, Y.; McVittie, B.; Barbulescu, D.M.; Salisbury, P.A.; Marcroft, S.; Delourme, R. Stable Quantitative Resistance Loci to Blackleg Disease in Canola (Brassica napus L.) Over Continents. Front. Plant Sci. 2018, 9, 9. [Google Scholar] [CrossRef] [Green Version]
- Soomro, W.M. Characterizing Avr Genes of Leptosphaeria maculans and Resistance Responses among Commercial Canola Cultivars in Western; Canada University of Saskatchewan: Saskatoon, SK, Canada, 2016. [Google Scholar]
- Poland, J.; Balint-Kurti, P.; Wisser, R.J.; Pratt, R.C.; Nelson, R.J. Shades of gray: The world of quantitative disease resistance. Trends Plant Sci. 2009, 14, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Larkan, N.; Raman, H.; Lydiate, D.J.; Robinson, S.J.; Yu, F.; Barbulescu, D.M.; Rosy, R.; Luckett, D.J.; Burton, W.; Wratten, N.; et al. Multi-environment QTL studies suggest a role for cysteine-rich protein kinase genes in quantitative resistance to blackleg disease in Brassica napus. BMC Plant Biol. 2016, 16, 183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raman, H.; Raman, R.; Coombes, N.; Song, J.; Diffey, S.; Kilian, A.; Lindbeck, K.; Barbulescu, D.M.; Batley, J.; Edwards, D.; et al. Genome-wide Association Study Identifies New Loci for Resistance to Leptosphaeria maculans in Canola. Front. Plant Sci. 2016, 7, 1513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fomeju, B.F.; Falentin, C.; Lassalle, G.; Manzanares-Dauleux, M.J.; Delourme, R. Homoeologous duplicated regions are involved in quantitative resistance of Brassica napus to stem canker. BMC Genom. 2014, 15, 498. [Google Scholar] [CrossRef] [Green Version]
- Jestin, C.; Lode, M.; Vallee, P.; Domin, C.; Falentin, C.; Horvais, R.; Coedel, S.; Manzanares-Dauleux, M.J.; Delourme, R. Association mapping of quantitative resistance for Leptosphaeria maculans in oilseed rape (Brassica napus L.). Mol. Breed. 2010, 27, 271–287. [Google Scholar] [CrossRef]
- Haddadi, P.; Ma, L.; Wang, H.; Borhan, M.H. Genome-wide transcriptome analyses provides insights into the lifestyle transition and effector repertoire of Leptosphaeria maculans during colonization of Brassica napus seedlings. Mol. Plant Pathol. 2015. [Google Scholar] [CrossRef] [Green Version]
- Sonah, H.; Zhang, X.; Deshmukh, R.; Borhan, M.H.; Fernando, W.G.D.; Bélanger, R.R. Comparative Transcriptomic Analysis of Virulence Factors in Leptosphaeria maculans during Compatible and Incompatible Interactions with Canola. Front. Plant Sci. 2016, 7, 86. [Google Scholar] [CrossRef] [Green Version]
- Becker, M.G.; Zhang, X.; Walker, P.L.; Wan, J.C.; Millar, J.L.; Khan, D.; Granger, M.J.; Cavers, J.D.; Chan, A.C.; Fernando, D.W.; et al. Transcriptome analysis of the Brassica napus-Leptosphaeria maculans pathosystem identifies receptor, signaling and structural genes underlying plant resistance. Plant J. 2017, 90, 573–586. [Google Scholar] [CrossRef] [Green Version]
- Zhou, T.; Xu, W.; Hirani, A.H.; Liu, Z.; Tuan, P.A.; Ayele, B.T.; Daayf, F.; McVetty, P.B.E.; Duncan, R.W.; Li, G. Transcriptional Insight Into Brassica napus Resistance Genes LepR3 and Rlm2-Mediated Defense Response Against the Leptosphaeria maculans Infection. Front. Plant Sci. 2019, 10, 823. [Google Scholar] [CrossRef]
- Hao, Y.; Wang, T.; Wang, K.; Wang, X.; Fu, Y.; Huang, L.; Kang, Z. Transcriptome Analysis Provides Insights into the Mechanisms Underlying Wheat Plant Resistance to Stripe Rust at the Adult Plant Stage. PLoS ONE 2016, 11, e0150717. [Google Scholar] [CrossRef] [PubMed]
- Joshi, R.K.; Megha, S.; Rahman, M.H.; Basu, U.; Kav, N.N. A global study of transcriptome dynamics in canola (Brassica napus L.) responsive to Sclerotinia sclerotiorum infection using RNA-Seq. Gene 2016, 590, 57–67. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.-J.; Jestin, C.; Welham, S.J.; King, G.J.; Manzanares-Dauleux, M.J.; Fitt, B.D.L.; Delourme, R. Identification of environmentally stable QTL for resistance against Leptosphaeria maculans in oilseed rape (Brassica napus). Theor. Appl. Genet. 2015, 129, 169–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.-J.; Qi, A.; King, G.J.; Fitt, B.D.L. Assessing Quantitative Resistance against Leptosphaeria maculans (Phoma Stem Canker) in Brassica napus (Oilseed Rape) in Young Plants. PLoS ONE 2014, 9, e84924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wendenbaum, S.; Demange, P.; Dell, A.; Meyer, J.; Abdallah, M. The structure of pyoverdine Pa, the siderophore of Pseudomonas aeruginosa. Tetrahedron Lett. 1983, 24, 4877–4880. [Google Scholar] [CrossRef]
- Zwiers, L.-H.; Roohparvar, R.; De Waard, M.A. MgAtr7, a new type of ABC transporter from Mycosphaerella graminicola involved in iron homeostasis. Fungal Genet. Boil. 2007, 44, 853–863. [Google Scholar] [CrossRef]
- Kim, Y.; Park, S.-Y.; Kim, D.; Choi, J.; Lee, Y.-H.; Lee, J.-H.; Choi, W. Genome-scale analysis of ABC transporter genes and characterization of the ABCC type transporter genes in Magnaporthe oryzae. Genomics 2013, 101, 354–361. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Wang, Z.; Cheng, D.; Chen, X.; Chen, Y.; Ma, Z. The ATP-binding protein FgArb1 is essential for penetration, infectious and normal growth ofFusarium graminearum. New Phytol. 2018, 219, 1447–1466. [Google Scholar] [CrossRef] [Green Version]
- Yorimitsu, T.; Sato, K.; Takeuchi, M. Molecular mechanisms of Sar/Arf GTPases in vesicular trafficking in yeast and plants. Front. Plant Sci. 2014, 5, 411. [Google Scholar] [CrossRef] [Green Version]
- Bioinformatics & Evolutionary Genomics. Available online: http://bioinformatics.psb.ugent.be/webtools/Venn/ (accessed on 3 March 2017).
- Hubbard, M.; Peng, G. Quantitative resistance against an isolate of Leptosphaeria maculans (blackleg) in selected Canadian canola cultivars remains effective under increased temperatures. Plant Pathol. 2018, 67, 1329–1338. [Google Scholar] [CrossRef]
- Huang, Y.-J.; Paillard, S.; Kumar, V.; King, G.J.; Fitt, B.D.L.; Delourme, R. Oilseed rape (Brassica napus) resistance to growth of Leptosphaeria maculans in leaves of young plants contributes to quantitative resistance in stems of adult plants. PLoS ONE 2019, 14, e0222540. [Google Scholar] [CrossRef] [PubMed]
- Tenhaken, R.; Doerks, T.; Bork, P. DCD—A novel plant specific domain in proteins involved in development and programmed cell death. BMC Bioinform. 2005, 6, 169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reis, P.A.B.; Carpinetti, P.A.; Freitas, P.P.; Santos, E.G.; Camargos, L.F.; De Oliveira, I.H.T.; Silva, J.C.F.; Carvalho, H.H.; Dal-Bianco, M.; Soares-Ramos, J.R.; et al. Functional and regulatory conservation of the soybean ER stress-induced DCD/NRP-mediated cell death signaling in plants. BMC Plant Biol. 2016, 16, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, Y.; Wang, H.; Zou, Y.; Liu, C.; Liu, Y.; Wang, Y.; Zhang, W. Over-expression of mitochondrial heat shock protein 70 suppresses programmed cell death in rice. FEBS Lett. 2010, 585, 231–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoeberichts, F.A.; De Jong, A.J.; Woltering, E.J. Apoptotic-like cell death marks the early stages of gypsophila (Gypsophila paniculata) petal senescence. Postharvest Boil. Technol. 2005, 35, 229–236. [Google Scholar] [CrossRef]
- Ruberti, C.; Lai, Y.; Brandizzi, F. Recovery from temporary endoplasmic reticulum stress in plants relies on the tissue-specific and largely independent roles of bZIP28 and bZIP60, as well as an antagonizing function of BAX-Inhibitor 1 upon the pro-adaptive signaling mediated by bZIP28. Plant J. 2017, 93, 155–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babaeizad, V.; Imani, J.; Kogel, K.-H.; Eichmann, R.; Huckelhoven, R. Over-expression of the cell death regulator BAX inhibitor-1 in barley confers reduced or enhanced susceptibility to distinct fungal pathogens. Theor. Appl. Genet. 2008, 118, 455–463. [Google Scholar] [CrossRef]
- Scotton, D.C.; Azevedo, M.D.S.; Sestari, I.; Da Silva, J.S.; Souza, L.A.; Peres, L.E.P.; Leal, G.A.; Figueira, A. Expression of the Theobroma cacao Bax-inhibitor-1 gene in tomato reduces infection by the hemibiotrophic pathogen Moniliophthora perniciosa. Mol. Plant Pathol. 2016, 18, 1101–1112. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Mendoza, M.; Velasco-Arroyo, B.; González-Melendi, P.; Martinez, M.; Diaz, I. C1A cysteine protease–cystatin interactions in leaf senescence. J. Exp. Bot. 2014, 65, 3825–3833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okamoto, T.; Shimada, T.; Hara-Nishimura, I.; Nishimura, M.; Minamikawa, T. C-Terminal KDEL Sequence of A KDEL-Tailed Cysteine Proteinase (Sulfhydryl-Endopeptidase) Is Involved in Formation of KDEL Vesicle and in Efficient Vacuolar Transport of Sulfhydryl-Endopeptidase1. Plant Physiol. 2003, 132, 1892–1900. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara-Nishimura, I.; Inoue, K.; Nishimura, M. A unique vacuolar processing enzyme responsible for conversion of several proprotein precursors into the mature forms. FEBS Lett. 1991, 294, 89–93. [Google Scholar] [CrossRef] [Green Version]
- Hara-Nishimura, I.; Nishimura, M. Proglobulin Processing Enzyme in Vacuoles Isolated from Developing Pumpkin Cotyledons. Plant Physiol. 1987, 85, 440–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zamyatnin, A.A., Jr. Plant Proteases Involved in Regulated Cell Death. Biochemistry (Moscow) 2015, 80, 1701–1715. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Zhang, H.; Deng, X.; Liu, J.; Chen, H. The relationship between vacuolation and initiation of PCD in rice (Oryza sativa) aleurone cells. Sci. Rep. 2017, 7, 41245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sueldo, D.J.; Van Der Hoorn, R.A.L. Plant life needs cell death, but does plant cell death need Cys proteases? FEBS J. 2017, 284, 1577–1585. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.-H.; Liu, R.; Yan, L.; Liu, Z.-Q.; Jiang, S.-C.; Shen, Y.-Y.; Wang, X.-F.; Zhang, D.-P. Light-harvesting chlorophyll a/b-binding proteins are required for stomatal response to abscisic acid in Arabidopsis. J. Exp. Bot. 2011, 63, 1095–1106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gálvez-Valdivieso, G.; Mullineaux, P.M. The role of reactive oxygen species in signalling from chloroplasts to the nucleus. Physiol. Plant. 2010, 138, 430–439. [Google Scholar] [CrossRef]
- Ahn, I.-P.; Kim, S.; Lee, Y.-H. Vitamin B1 Functions as an Activator of Plant Disease Resistance1. Plant Physiol. 2005, 138, 1505–1515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahn, I.-P.; Kim, S.; Lee, Y.-H.; Suh, S.-C. Vitamin B1-Induced Priming Is Dependent on Hydrogen Peroxide and the NPR1 Gene in Arabidopsis. Plant Physiol. 2006, 143, 838–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boubakri, H.; Wahab, M.A.; Chong, J.; Bertsch, C.; Mliki, A.; Soustre-Gacougnolle, I. Thiamine induced resistance to plasmopara viticola in grapevine and elicited host–defense responses, including HR like-cell death. Plant Physiol. Biochem. 2012, 57, 120–133. [Google Scholar] [CrossRef]
- Jain, B.P.; Pandey, S. WD40 Repeat Proteins: Signalling Scaffold with Diverse Functions. Protein J. 2018, 37, 391–406. [Google Scholar] [CrossRef] [PubMed]
- Nalefski, E.A.; Wisner, M.A.; Chen, J.Z.; Sprang, S.R.; Fukuda, M.; Mikoshiba, K.; Falke, J.J. C2 Domains from Different Ca2+ Signaling Pathways Display Functional and, Mechanistic Diversity. Biochemistry 2001, 40, 3089–3100. [Google Scholar] [CrossRef]
- Andersen, E.J.; Ali, S.; Byamukama, E.; Yen, Y.; Nepal, M.P. Disease Resistance Mechanisms in Plants. Genes 2018, 9, 339. [Google Scholar] [CrossRef] [Green Version]
- Williams, B.; Verchot, J.; Dickman, M. When supply does not meet demand-ER stress and plant programmed cell death. Front. Plant Sci. 2014, 5, 211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, H.-H.; Wang, M.-H.; Lee, Y.-L.; Tsai, Y.; Li, Y.-H.; Yang, F.-J.; Liao, Y.-C.; Lin, S.; Lai, E.-M. Agrobacterium-produced and exogenous cytokinin-modulated Agrobacterium-mediated plant transformation. Mol. Plant Pathol. 2010, 11, 677–690. [Google Scholar] [CrossRef] [PubMed]
- Kutcher, H.; Balesdent, M.H.; Rimmer, S.R.; Rouxel, T.; Chevre, A.-M.; Delourme, R.; Brun, H. Frequency of avirulence genes in Leptosphaeria maculans in western Canada. Can. J. Plant Pathol. 2010, 32, 77–85. [Google Scholar] [CrossRef]
- Koch, E.; Badawy, H.M.A.; Hoppe, H.H. Differences Between Aggressive and Non-Aggressive Single Spore Lines of Leptosphaeria maculans in Cultural Characteristics and Phytotoxin Production. J. Phytopathol. 1989, 124, 52–62. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genoscope. Available online: http://www.genoscope.cns.fr/brassicanapus/data/ (accessed on 18 October 2016).
- Joint Genome Institute, Genome Portal. Available online: http://genome.jgi.doe.gov/ (accessed on 2 November 2016).
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2012, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, M.; Huber, W.; Pagès, H.; Aboyoun, P.; Carlson, M.; Gentleman, R.; Morgan, M.; Carey, V.J. Software for Computing and Annotating Genomic Ranges. PLoS Comput. Biol. 2013, 9, e1003118. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conesa, A.; Götz, S. Blast2GO: A Comprehensive Suite for Functional Analysis in Plant Genomics. Int. J. Plant Genom. 2007, 2008, 1–12. [Google Scholar] [CrossRef] [PubMed]










© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hubbard, M.; Zhai, C.; Peng, G. Exploring Mechanisms of Quantitative Resistance to Leptosphaeria maculans (Blackleg) in the Cotyledons of Canola (Brassica napus) Based on Transcriptomic and Microscopic Analyses. Plants 2020, 9, 864. https://doi.org/10.3390/plants9070864
Hubbard M, Zhai C, Peng G. Exploring Mechanisms of Quantitative Resistance to Leptosphaeria maculans (Blackleg) in the Cotyledons of Canola (Brassica napus) Based on Transcriptomic and Microscopic Analyses. Plants. 2020; 9(7):864. https://doi.org/10.3390/plants9070864
Chicago/Turabian StyleHubbard, Michelle, Chun Zhai, and Gary Peng. 2020. "Exploring Mechanisms of Quantitative Resistance to Leptosphaeria maculans (Blackleg) in the Cotyledons of Canola (Brassica napus) Based on Transcriptomic and Microscopic Analyses" Plants 9, no. 7: 864. https://doi.org/10.3390/plants9070864