QTL Mapping and Prediction of Haploid Male Fertility Traits in Maize (Zea mays L.)

 , ,
, ,
Abstract
:1. Introduction
2. Results
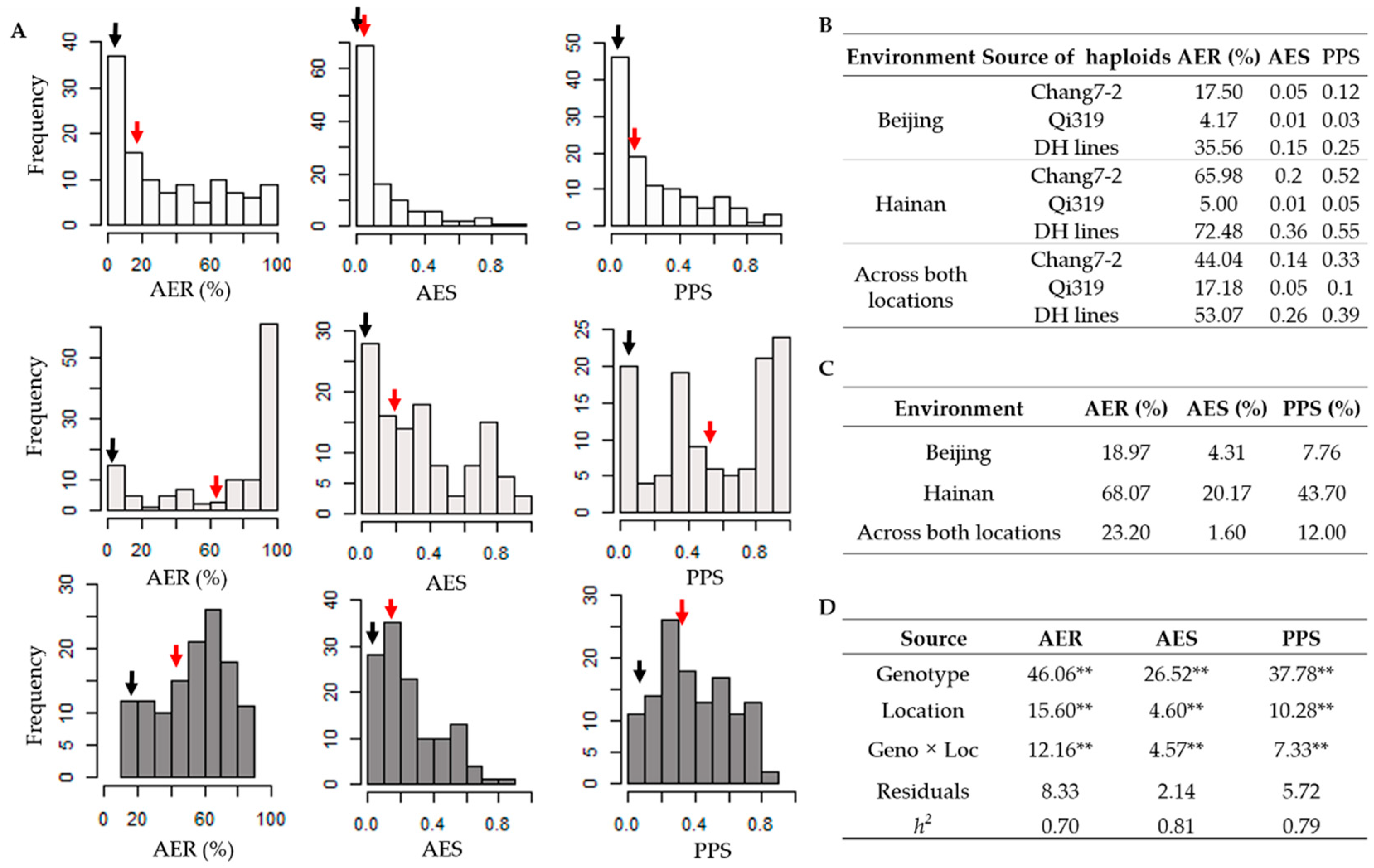
2.1. Phenotypic Evaluation of HMF Traits for DH Population
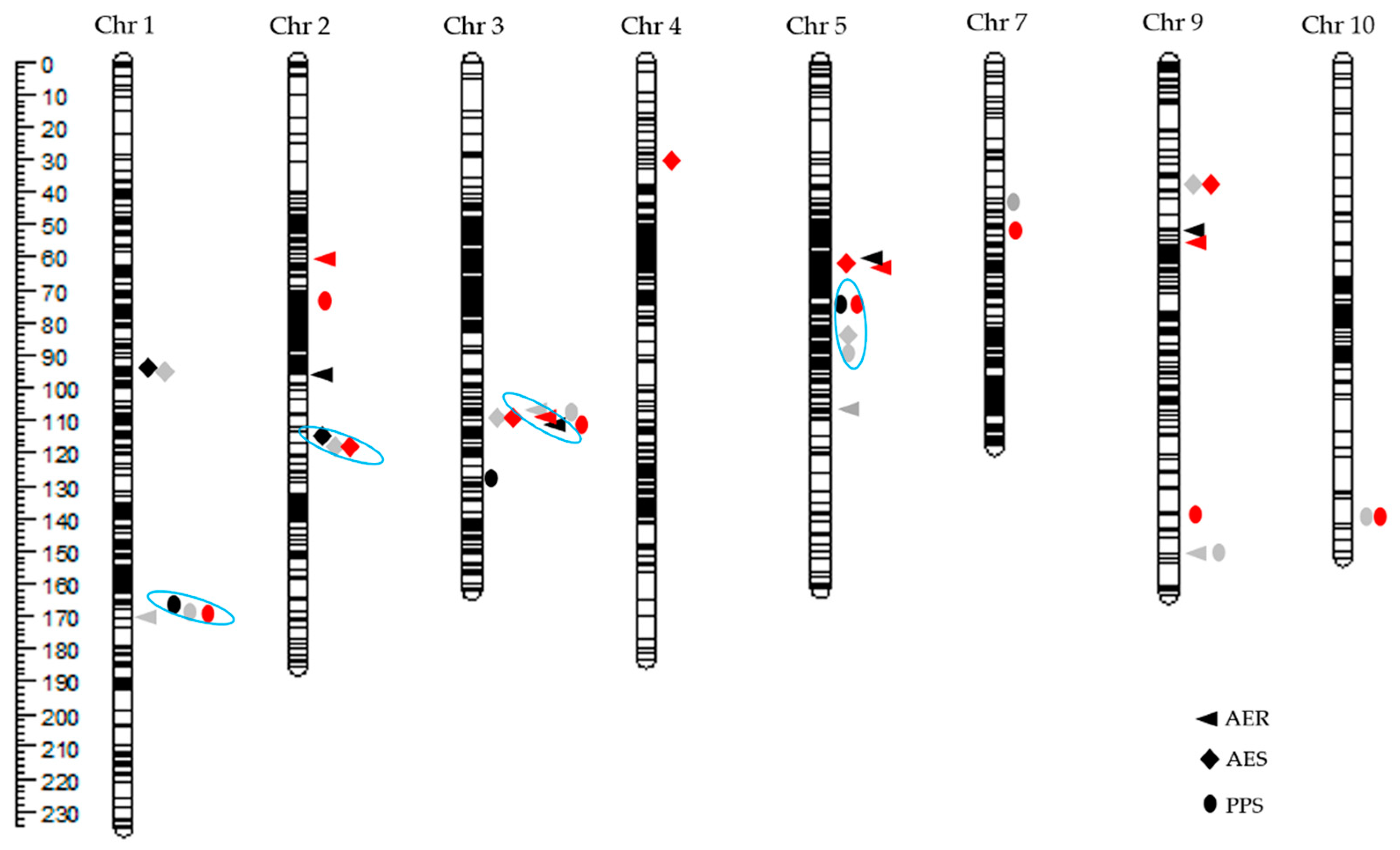
2.2. QTL Analyses of Three HMF-Related Traits
2.2.1. QTL for AER
2.2.2. QTL for AES
2.2.3. QTL for PPS
2.3. The Segregation Distortion of the Putative QTL for Three HMF-Related Traits
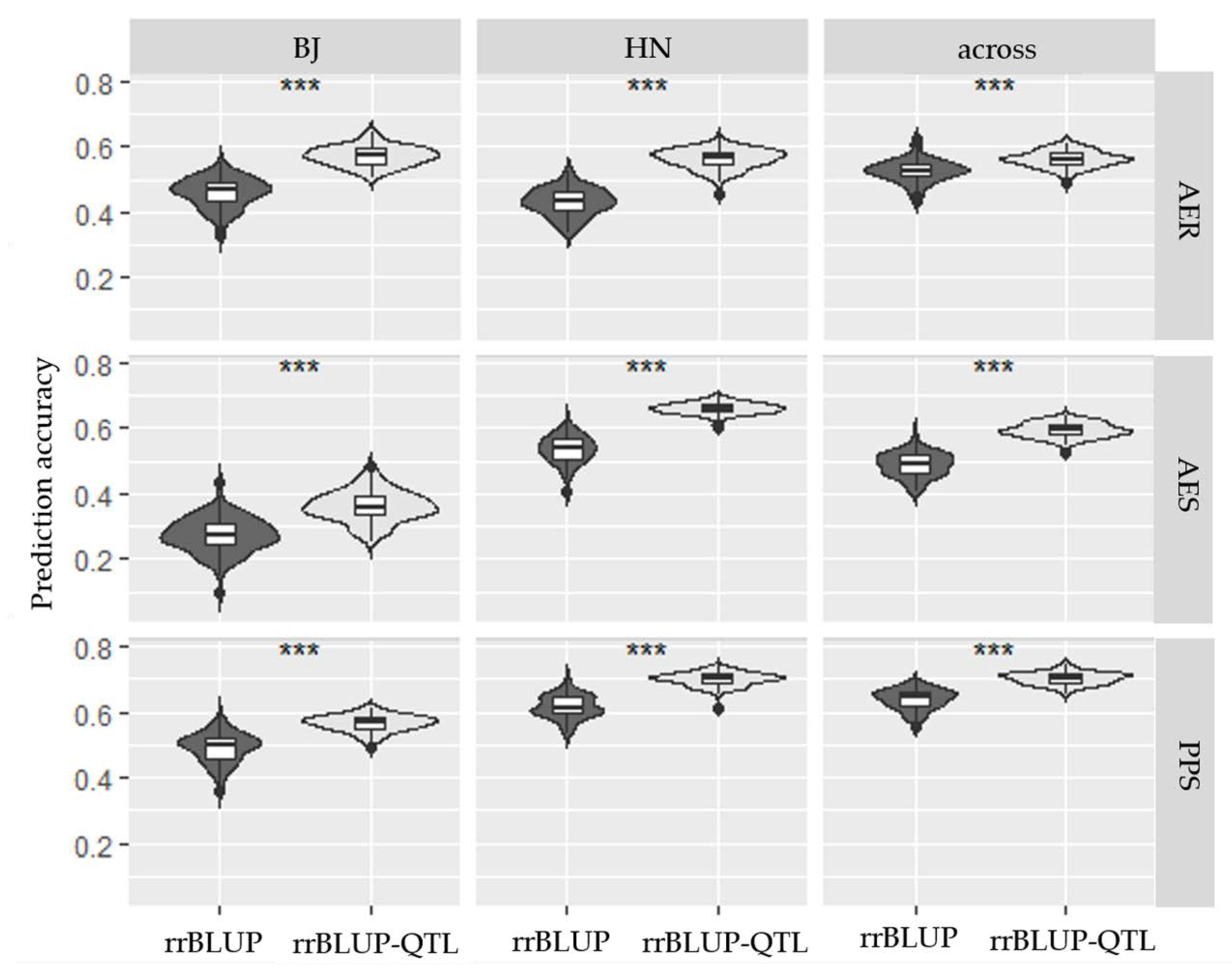
2.4. Genomic Prediction (GP) for Three HMF-Related Traits
3. Discussion
3.1. The Evaluation Standard of HMF
3.2. Factors Affecting HMF
3.3. QTL for HMF
3.4. GP for HMF Application in Breeding Schedule
4. Materials and Methods
4.1. Plant Materials and Haploid Induction
4.2. Phenotypic Evaluation and Statistical Analysis
4.3. Genotyping and QTL Mapping
4.4. GP Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Chaikam, V.; Molenaar, W.; Melchinger, A.E.; Boddupalli, P.M. Doubled haploid technology for line development in maize: Technical advances and prospects. Theor. Appl. Genet. 2019, 132, 3227–3243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prigge, V.; Xu, X.; Li, L.; Babu, R.; Chen, S.; Atlin, G.N.; Melchinger, A.E. New Insights into the Genetics of in Vivo Induction of Maternal Haploids, the Backbone of Doubled Haploid Technology in Maize. Genetics 2012, 190, 781–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.J. Maize Haploid Breeding Technology; China Agricultural University Press: Beijing, China, 2012. [Google Scholar]
- Molenaar, W.S.; Schipprack, W.; Brauner, P.C.; Melchinger, A.E. Haploid male fertility and spontaneous chromosome doubling evaluated in a diallel and recurrent selection experiment in maize. Theor. Appl. Genet. 2019, 132, 2273–2284. [Google Scholar] [CrossRef]
- Chaikam, V.; Prasanna, B. Maternal Haploid Detection Using Anthocyanin Markers. In B.M. prasanna, editor, Doubled haploid technology in maize breeding; International Maize and Wheat Improvement Center: DF, Mexico, 2012; pp. 20–23. [Google Scholar]
- Nanda, D.K.; Chase, S.S. An embryo marker for detecting monoploids of maize (Zea Mays L.). Crop Sci. 1966, 2, 213–215. [Google Scholar] [CrossRef]
- Melchinger, A.E.; Schipprack, W.; Würschum, T.; Chen, S.; Technow, F. Rapid and accurate identification of in vivo-induced haploid seeds based on oil content in maize. Sci. Rep. 2013, 3, 2129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Song, T. Identification Haploid with High Oil Xenia Effect in Maize. Acta Agron. Sin. 2003, 4, 587–590. [Google Scholar]
- Qin, H.; Ma, J.Y.; Chen, S.J.; Yan, Y.L.; Li, W.; Wang, P.; Liu, J. Identification of Haploid Maize Kernel Using NIR Spectroscopy in Reflectance and Transmittance Modes: A Comparative Study. Guang Pu Xue Yu Guang Pu Fen Xi= Guang Pu 2016, 1, 292–297. [Google Scholar]
- Wang, Y.; Lv, Y.; Liu, H.; Wei, Y.; Zhang, J.; An, D.; Wu, J. Identification of maize haploid kernels based on hyperspectral imaging technology. Comput. Electron. Agric. 2018, 153, 188–195. [Google Scholar] [CrossRef]
- Aslam, M.; Farid, B.; Khakwani, K.; Maqbool, M.A.; Zou, H. In vivo Maternal Haploid Seed Production and Chromosome Doubling with different Anti-microtubular Agents in Maize. Int. J. Agric. Biol. 2017, 19, 114–120. [Google Scholar] [CrossRef]
- Jiang, L.; Yang, X.Y.; Li, X.Y.; Deng, P.K.; Jing, G.X.; Wang, X.Q.; Xing, Z.; Zhao, R.G. A study on natural recovery of tassel fertilization and doubling method in maize haploids. Genet. Mol. Res. 2017, 22, gmr16019173. [Google Scholar] [CrossRef]
- Häntzschel, K.R.; Weber, G. Blockage of mitosis in maize root tips using colchicine-alternatives. Protoplasma 2010, 241, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Gayen, P.; Madan, J.K.; Kumar, R.; Sarkar, K.R. Chromosome doubling in haploids through colchicine. Maize Genet. Coop. Newsl. 1994, 68, 65. [Google Scholar]
- Eder, J.; Chalyk, S. In vivo haploid induction in maize. Theor. Appl. Genet. 2002, 4, 703–708. [Google Scholar] [CrossRef]
- Ke, W.; Liang, L.; Yuqiang, L.; Shaojiang, C. Study on bio-haploid inducing and doubling efficiency in maize. J. China Agric. Univ. 2006, 11, 17–20. [Google Scholar]
- Melchinger, A.E.; Molenaar, W.S.; Mirdita, V.; Schipprack, W. Colchicine Alternatives for Chromosome Doubling in Maize Haploids for Doubled-Haploid Production. Crop Sci. 2016, 56, 559. [Google Scholar] [CrossRef]
- Kitamura, S.; Akutsu, M.; Okazaki, K. Mechanism of action of nitrous oxide gas applied as a polyploidizing agent during meiosis in lilies. Sex. Plant Reprod. 2009, 1, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Molenaar, W.S.; Schipprack, W.; Melchinger, A.E. Nitrous Oxide-Induced Chromosome Doubling of Maize Haploids. Crop Sci. 2018, 58, 650–659. [Google Scholar] [CrossRef]
- Geiger, H.H. Doubled haploids. In Maize Handbook—Volume II: Genetics and Genomics; Spring: New York, NY, USA, 2009; pp. 641–657. [Google Scholar]
- Ren, J.; Wu, P.; Trampe, B.; Tian, X.; Lübberstedt, T.; Chen, S. Novel technologies in doubled haploid line development. Plant Biotechnol. J. 2017, 15, 1361–1370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geiger, H.H.; Braun, M.D.; Gordillo, G.A.; Koch, S.; Jesse, J.; Krutzfeldt, B.A. Variation for female fertility among haploid maize lines. Maize Genet. Coop. Newsl. 2006, 80, 28. [Google Scholar]
- Chalyk, S.T. Properties of maternal haploid maize plants and potential application to maize breeding. Euphytica 1994, 79, 13–18. [Google Scholar] [CrossRef]
- Wu, P.; Ren, J.; Tian, X.; Lübberstedt, T.; Li, W.; Li, G.; Li, X.; Chen, S. New Insights into the Genetics of Haploid Male Fertility in Maize. Crop Sci. 2017, 57, 637. [Google Scholar] [CrossRef] [Green Version]
- Kleiber, D.; Prigge, V.; Melchinger, A.E.; Burkard, F.; San Vicente, F.; Palomino, G.; Gordillo, G.A. Haploid Fertility in Temperate and Tropical Maize Germplasm. Crop Sci. 2012, 52, 623. [Google Scholar] [CrossRef]
- Ma, H.; Li, G.; Würschum, T.; Zhang, Y.; Zheng, D.; Yang, X.; Li, J.; Liu, W.; Yan, J.; Chen, S. Genome-Wide Association Study of Haploid Male Fertility in Maize (Zea Mays L.). Front. Plant Sci. 2018, 9, 974. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Wu, P.; Tian, X.; Lübberstedt, T.; Chen, S. QTL mapping for haploid male fertility by a segregation distortion method and fine mapping of a key QTL qhmf4 in maize. Theor. Appl. Genet. 2017, 130, 1349–1359. [Google Scholar] [CrossRef]
- Chaikam, V.; Gowda, M.; Nair, S.K.; Melchinger, A.E.; Boddupalli, P.M. Genome-wide association study to identify genomic regions influencing spontaneous fertility in maize haploids. Euphytica 2019, 215, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Qu, Y.; Chen, Q.; Tang, J.; Lübberstedt, T.; Li, H.; Liu, Z. Genetic dissection of haploid male fertility in maize (Zea mays L.). Plant Breed. 2019, 138, 259–265. [Google Scholar] [CrossRef] [Green Version]
- Ren, J.; Boerman, N.A.; Liu, R.; Wu, P.; Trampe, B.; Vanous, K.; Frei, U.K.; Chen, S.; Lübberstedt, T. Mapping of QTL and identification of candidate genes conferring spontaneous haploid genome doubling in maize (Zea mays L.). Plant Sci. 2020, 293, 110337. [Google Scholar] [CrossRef]
- Li, D.; Xu, Z.; Gu, R.; Wang, P.; Lyle, D.; Xu, J.; Zhang, H.; Wang, G. Enhancing genomic selection by fitting large-effect SNPs as fixed effects and a genotype-by-environment effect using a maize BC1F3:4 population. PLoS ONE 2019, 14, e0223898. [Google Scholar] [CrossRef] [PubMed]
- Nyaga, C.; Gowda, M.; Beyene, Y.; Muriithi, W.T.; Makumbi, D.; Olsen, M.S.; Suresh, L.M.; Bright, J.M.; Das, B.; Prasanna, B.M. Genome-Wide Analyses and Prediction of Resistance to MLN in Large Tropical Maize Germplasm. Genes 2020, 11, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W. Analysis on In Vivo Haploid Induction Effects and Machine Learning Application in Doubled Haploid Technology of Maize; China Agricultural University: Beijing, China, 2017. [Google Scholar]
- Broman, K.W.; Wu, H.; Sen, S.; Churchill, G.A. R/qtl: QTL mapping in experimental crosses. Bioinformatics 2003, 19, 889–890. [Google Scholar] [CrossRef] [Green Version]
- Visscher, P.M.; Thompson, R.; Haley, C.S. Confidence Intervals in QTL Mapping by Bootstrapping. Genetics 1996, 143, 1013–1020. [Google Scholar] [PubMed]
- Endelman, J.B. Ridge Regression and Other Kernels for Genomic Selection with R Package rrBLUP. Plant Genome 2011, 4, 250–255. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Environment | QTL Name | Marker | Chr | Position (cM) | LOD | Add | PVE (%) | Allele | χ2 | p-Value | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Q | C | ||||||||||
| Beijing | qAER2-1 | PZE-102130712 | 2 | 98.24 | 2.68 | −0.08 | 0.21 | 14 | 8 | 1.64 | 0.201 |
| qAER3 | PZE-103145047 | 3 | 113.47 | 5.70 | −0.26 | 8.02 | 17 | 5 | 6.55 | 0.011 | |
| qAER5-1 | PZE-105077018 | 5 | 64.39 | 3.56 | 0.27 | 14.60 | 2 | 19 | 13.76 | <0.001 | |
| qAER9-1 | SYN34182 | 9 | 51.12 | 2.90 | 0.14 | 4.72 | 7 | 15 | 2.91 | 0.088 | |
| Hainan | qAER1 | PZE-101207960 | 1 | 173.69 | 3.49 | 0.17 | 4.22 | 26 | 55 | 10.38 | 0.001 |
| qAER3 | PZE-103135808 | 3 | 108.06 | 2.51 | −0.17 | 6.41 | 46 | 30 | 3.37 | 0.066 | |
| qAER5-2 | SYN1878 | 5 | 109.29 | 8.21 | 0.34 | 19.69 | 19 | 62 | 22.83 | <0.001 | |
| qAER9-2 | PZE-109115897 | 9 | 152.41 | 2.82 | 0.14 | 4.00 | 33 | 48 | 2.78 | 0.096 | |
| Across both locations | qAER2-2 | PZE-102055945 | 2 | 62.11 | 2.83 | −0.12 | 5.13 | 19 | 10 | 2.79 | 0.095 |
| qAER3 | PZE-103138981 | 3 | 111.82 | 4.92 | −0.16 | 7.80 | 21 | 8 | 5.83 | 0.016 | |
| qAER5-1 | PZE-105094949 | 5 | 67.29 | 4.71 | 0.20 | 19.51 | 3 | 26 | 16.33 | <0.001 | |
| qAER9-1 | PZE-109026940 | 9 | 55.90 | 2.54 | 0.10 | 5.64 | 10 | 19 | 2.79 | 0.095 | |
| Environment | QTL Name | Marker | Chr | Position (cM) | LOD | Add | PVE(%) | Allele | χ2 | p-Value | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Q | C | ||||||||||
| Beijing | qAES1 | PZE-101098199 | 1 | 93.80 | 2.65 | −0.04 | 1.17 | 5 | 0 | 5.00 | 0.025 |
| qAES2 | PZE-102141171 | 2 | 116.96 | 2.66 | −0.11 | 6.71 | 4 | 1 | 1.80 | 0.18 | |
| Hainan | qAES1 | PZE-101111793 | 1 | 96.70 | 3.25 | −0.14 | 4.59 | 19 | 5 | 8.17 | 0.004 |
| qAES2 | SYN216 | 2 | 120.65 | 2.71 | −0.13 | 4.55 | 18 | 5 | 7.35 | 0.007 | |
| qAES3 | PZE-103138981 | 3 | 111.81 | 6.93 | −0.26 | 15.61 | 19 | 5 | 8.17 | 0.004 | |
| qAES5-1 | PZE-105126493 | 5 | 86.97 | 5.16 | 0.26 | 11.42 | 2 | 22 | 16.67 | <0.001 | |
| qAES9 | PZE-109019490 | 9 | 39.03 | 4.23 | 0.18 | 10.28 | 4 | 20 | 10.67 | 0.001 | |
| Across both locations | qAES2 | SYN216 | 2 | 120.66 | 4.41 | −0.13 | 6.66 | 2 | 0 | 2.00 | 0.157 |
| qAES3 | PZE-103138981 | 3 | 111.82 | 5.44 | −0.13 | 10.59 | 1 | 1 | 0.00 | 1.000 | |
| qAES4 | PZE-104016598 | 4 | 31.62 | 3.70 | −0.13 | 8.60 | 2 | 0 | 2.00 | 0.157 | |
| qAES5-2 | PZE-105079012 | 5 | 64.36 | 3.46 | 0.13 | 11.29 | 0 | 2 | 2.00 | 0.157 | |
| qAES9 | PZE-109019490 | 9 | 39.03 | 3.89 | 0.11 | 8.40 | 0 | 2 | 2.00 | 0.157 | |
| Environment | QTL Name | Marker | Chr | Position (cM) | LOD | Add | PVE (%) | Allele | χ2 | p-Value | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Q | C | ||||||||||
| Beijing | qPPS1 | PZE-101204728 | 1 | 168.12 | 5.09 | 0.19 | 12.66 | 0 | 9 | 9.00 | 0.003 |
| qPPS3-1 | PZE-103162736 | 3 | 133.82 | 2.92 | −0.15 | 8.07 | 7 | 2 | 2.78 | 0.096 | |
| qPPS5 | SYN32709 | 5 | 77.05 | 3.34 | 0.19 | 9.60 | 0 | 9 | 9.00 | 0.003 | |
| Hainan | qPPS1 | PZE-101206252 | 1 | 170.81 | 4.95 | 0.21 | 12.36 | 11 | 41 | 17.31 | <0.001 |
| qPPS3-2 | PZE-103135808 | 3 | 108.06 | 3.96 | −0.21 | 4.49 | 31 | 18 | 3.45 | 0.063 | |
| qPPS5 | PZE-105136417 | 5 | 91.34 | 6.79 | 0.34 | 15.56 | 5 | 47 | 33.92 | <0.001 | |
| qPPS7 | PZE-107110443 | 7 | 45.60 | 3.27 | 0.21 | 8.48 | 10 | 42 | 19.69 | <0.001 | |
| qPPS9 | PZE-109115897 | 9 | 152.41 | 3.16 | 0.16 | 6.46 | 21 | 31 | 1.92 | 0.166 | |
| qPPS10 | PZE-110102769 | 10 | 141.81 | 4.04 | −0.19 | 7.70 | 35 | 17 | 6.23 | 0.013 | |
| Across both locations | qPPS1 | PZE-101206252 | 1 | 170.82 | 7.22 | 0.14 | 16.14 | 0 | 15 | 15.00 | <0.001 |
| qPPS2 | SYN24086 | 2 | 74.71 | 2.67 | −0.09 | 7.27 | 12 | 3 | 5.40 | 0.020 | |
| qPPS3-2 | PUT-163a-13490777-226 | 3 | 112.24 | 5.33 | −0.15 | 3.70 | 11 | 4 | 3.27 | 0.071 | |
| qPPS5 | SYN32709 | 5 | 77.05 | 8.96 | 0.22 | 14.31 | 0 | 15 | 15.00 | <0.001 | |
| qPPS7 | PZE-107104221 | 7 | 53.17 | 3.17 | 0.12 | 6.26 | 3 | 12 | 5.40 | 0.020 | |
| qPPS9 | PZE-109112516 | 9 | 142.84 | 2.78 | 0.08 | 4.22 | 5 | 10 | 1.67 | 0.197 | |
| qPPS10 | PZE-110102769 | 10 | 141.81 | 4.64 | −0.13 | 9.12 | 11 | 4 | 3.27 | 0.071 | |
| Trait | AER | AES | PPS |
|---|---|---|---|
| AER | 1 | 0.80 *** | 0.88 *** |
| AES | 1 | 0.73 *** | |
| PPS | 1 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiao, Y.; Li, J.; Li, W.; Chen, M.; Li, M.; Liu, W.; Liu, C.; Chen, S. QTL Mapping and Prediction of Haploid Male Fertility Traits in Maize (Zea mays L.). Plants 2020, 9, 836. https://doi.org/10.3390/plants9070836
Jiao Y, Li J, Li W, Chen M, Li M, Liu W, Liu C, Chen S. QTL Mapping and Prediction of Haploid Male Fertility Traits in Maize (Zea mays L.). Plants. 2020; 9(7):836. https://doi.org/10.3390/plants9070836
Chicago/Turabian StyleJiao, Yanyan, Jinlong Li, Wei Li, Ming Chen, Mengran Li, Wenxin Liu, Chenxu Liu, and Shaojiang Chen. 2020. "QTL Mapping and Prediction of Haploid Male Fertility Traits in Maize (Zea mays L.)" Plants 9, no. 7: 836. https://doi.org/10.3390/plants9070836