Mitigation of the Adverse Effects of the El Niño (El Niño, La Niña) Southern Oscillation (ENSO) Phenomenon and the Most Important Diseases in Avocado cv. Hass Crops



Abstract
:1. Introduction
2. Results
2.1. Adaptability of Native Rootstock Genotypes to Drought, Waterlogging, Tolerance to P. cinamomi and Verticillium sp. and Grafting Compatibility under Net House Condition
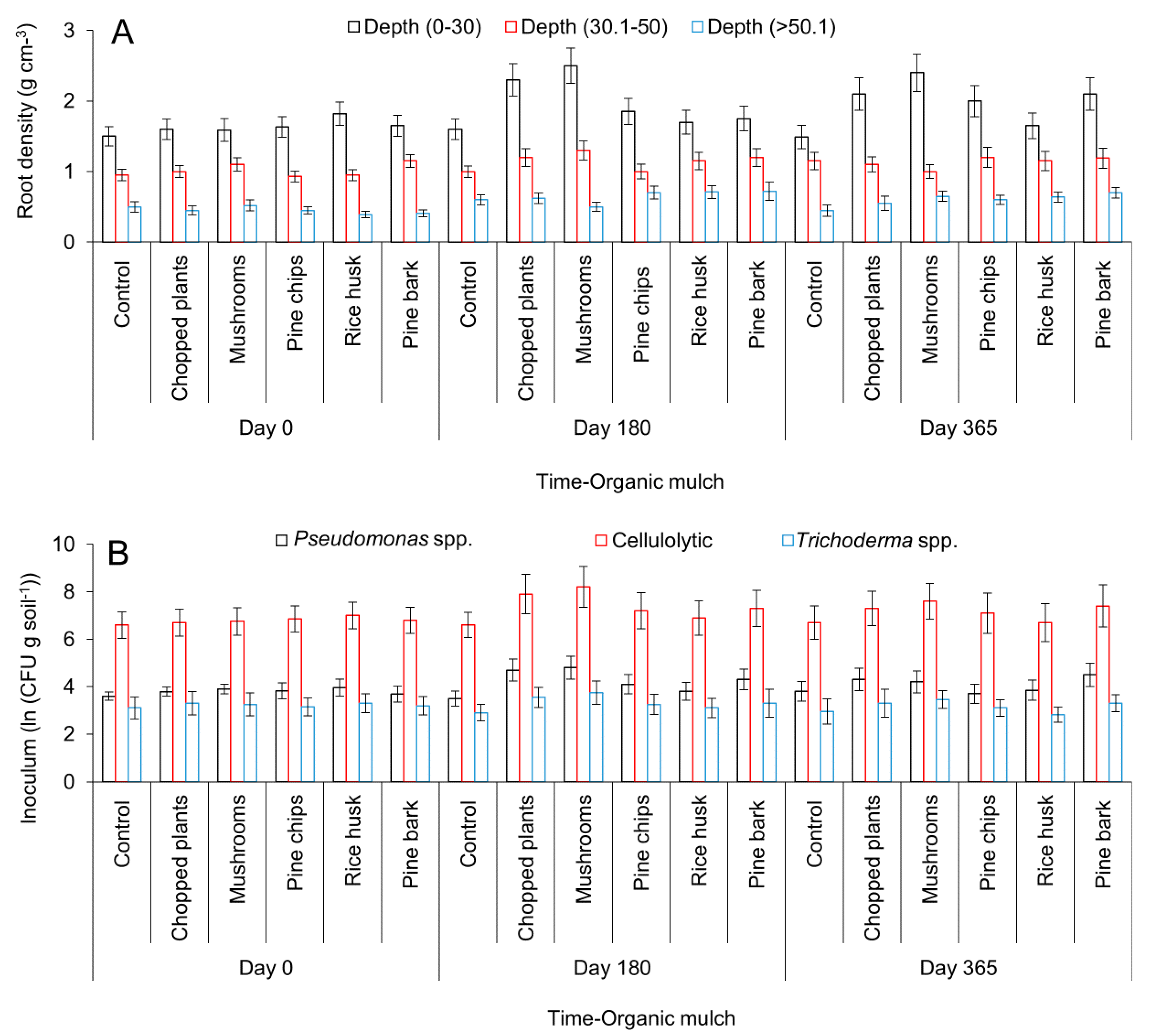
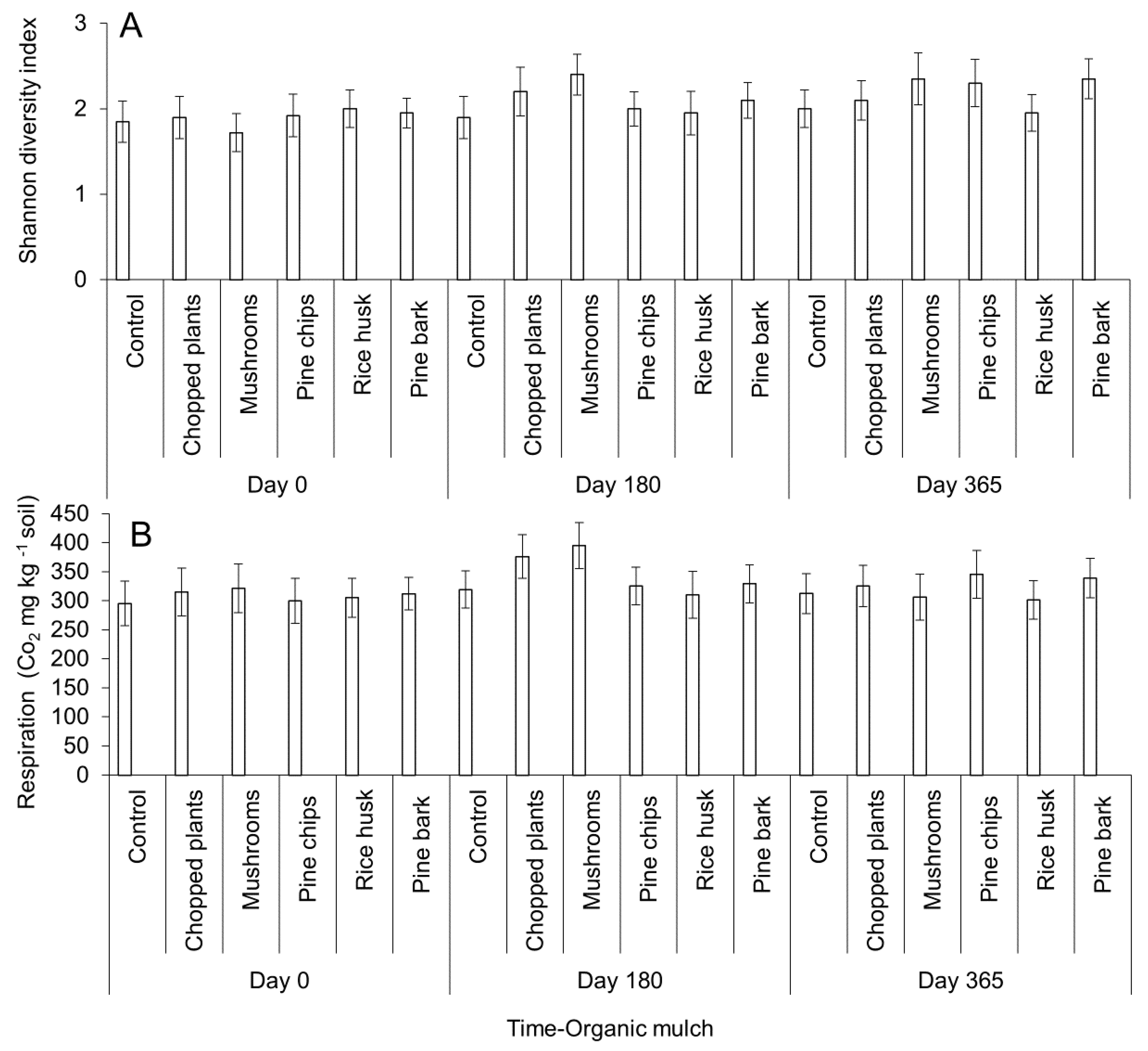
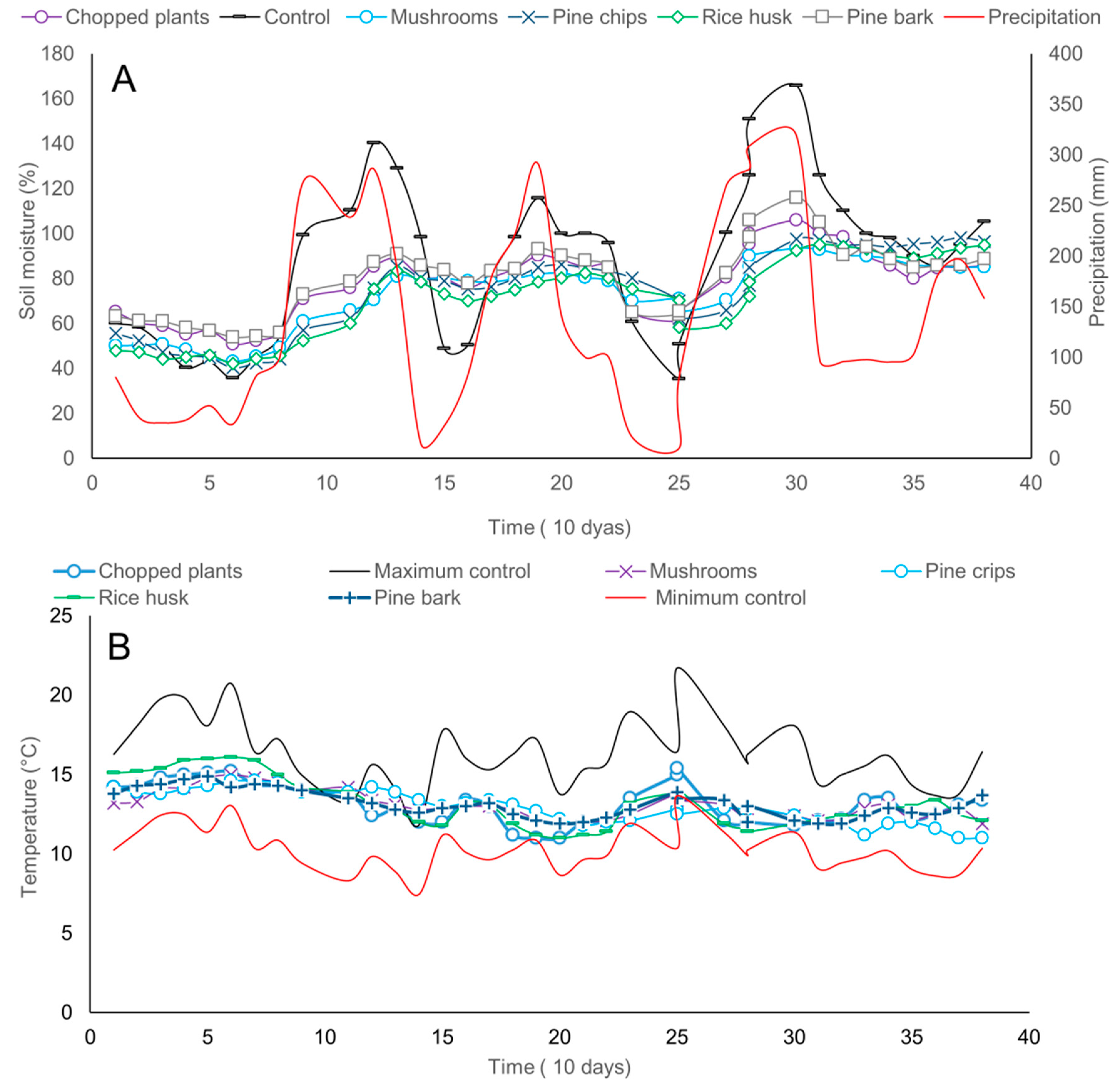
2.2. Effect of Organic Mulch Addition on Avocados Roots Growth, Microbial Populations, and Reduction of Stress Associated with Environmental Conditions under Field Conditions
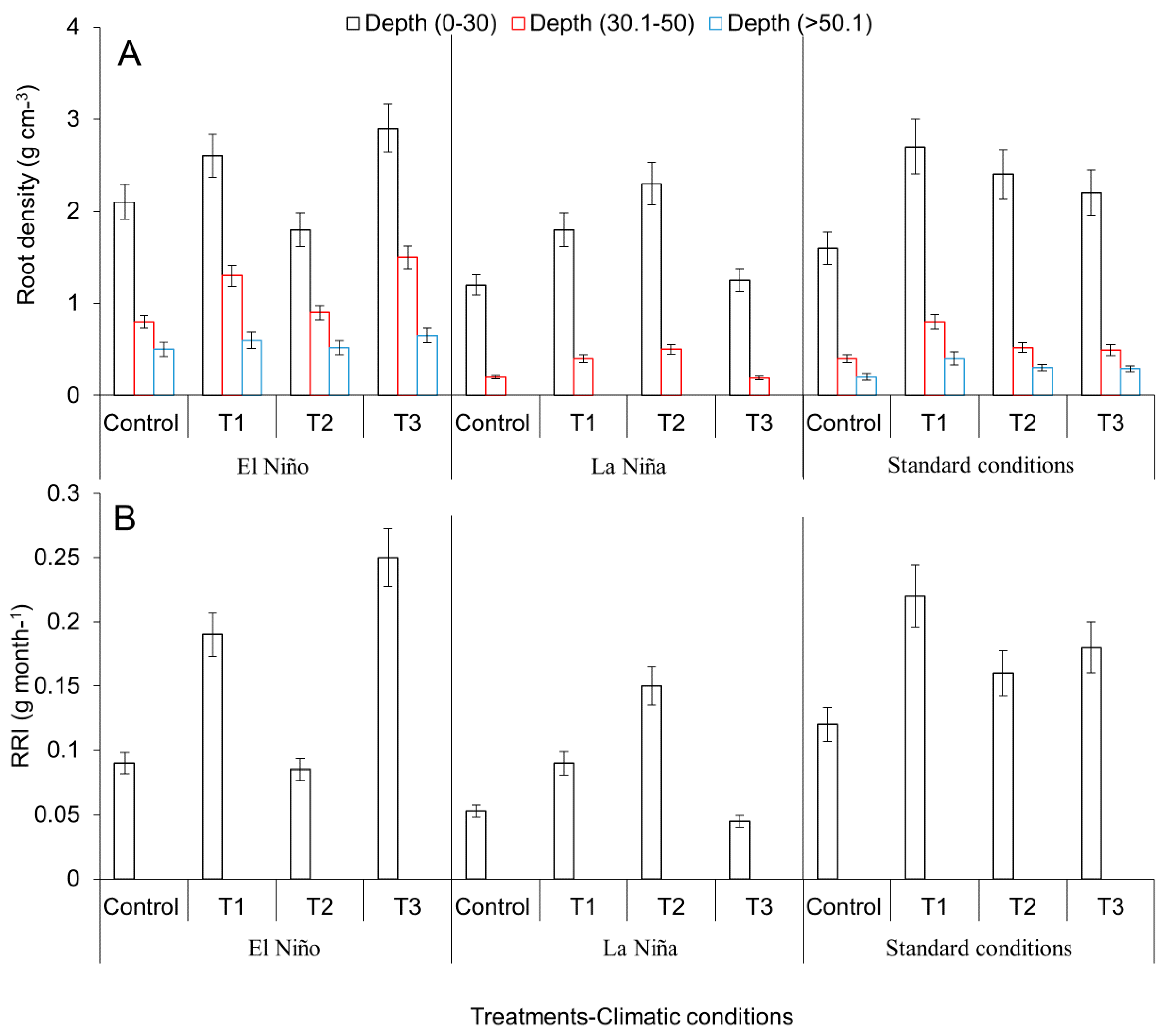
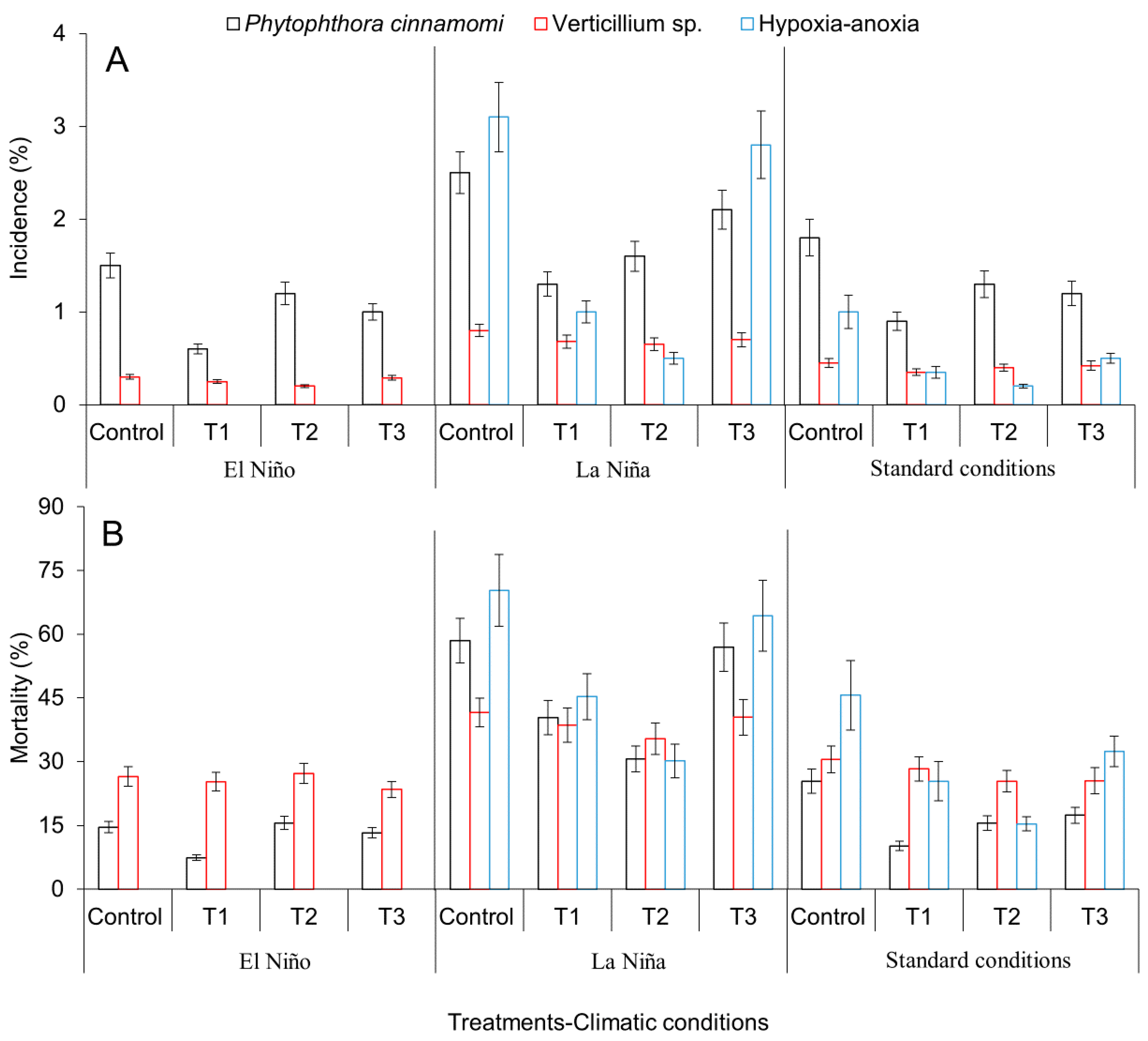
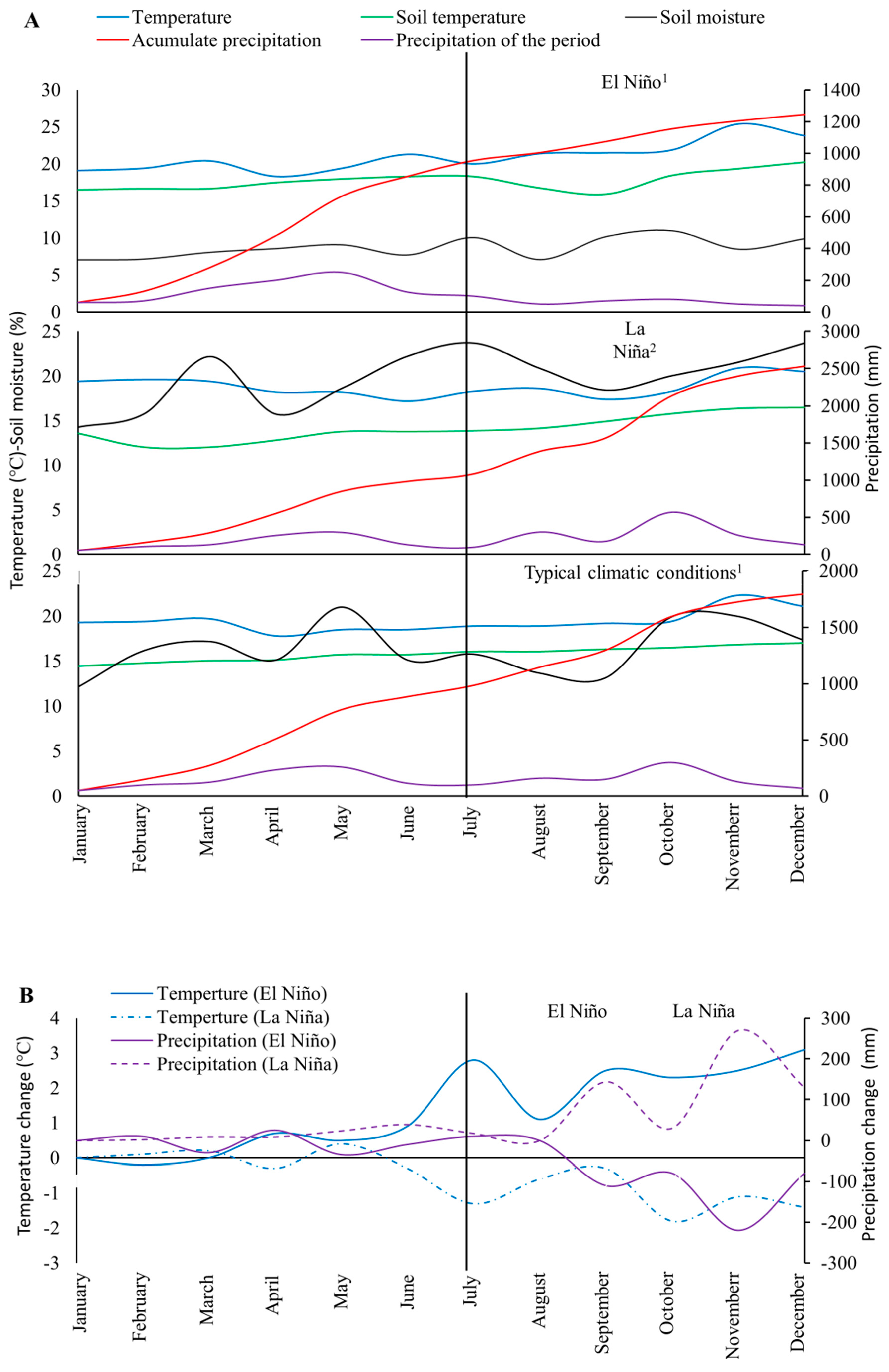
2.3. Evaluation of Agronomical Management Practices of Avocado Crops Aimed at Mitigating Adverse Effects of Climate Variability under the ENSO Phenomenon: El Niño and La Niña and Reduction of Incidence and Mortality of Diseases under Field Conditions
3. Discussion
3.1. Evaluation of Native Rootstock Adaptability to Different Soil Moisture Regimes, Tolerance to P. cinamomi and Verticillium sp., and Grafting Compatibility with Avocado cv. Hass under Net House Conditions
3.2. Evaluation of Agronomical Management Practices to Mitigate the Adverse Effects of Climatic Variability Associated with the ENSO Phenomenon (El Niño and La Niña) and Reduction of Incidence and Mortality of Diseases under Field Conditions
4. Materials and Methods
4.1. Study Location
4.2. Phase One
4.2.1. Evaluation of the Adaptability of Native Avocado Rootstocks to Different Soil Moisture Regimes, Resistance to P. cinamomi and Verticillium sp., and Grafting Compatibility with Avocado cv. Hass under Net House Conditions
4.2.2. Effect of Organic Mulch Addition on Avocados Roots Growth, Microbial Populations, and Reduction of Stress Associated with Environmental Conditions under Field Conditions
4.3. Phase Two
Agronomical Practices in Avocado to Mitigating Adverse Effects of Climate Variability under the ENSO Phenomenon and Incidence and Mortality of Diseases under Field Conditions
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Howden, S.M.; Soussana, J.-F.; Tubiello, F.N.; Chhetri, N.; Dunlop, M.; Meinke, H. Adapting agriculture to climate change. Proc. Natl. Acad. Sci. USA 2007, 104, 19691–19696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- IPCC. Fifth Assessment Report-Impacts, Adaptation and Vulnerability. Available online: http://www.ipcc.ch/report/ar5/wg2/ (accessed on 4 September 2017).
- Mall, R.K.; Gupta, A.; Sonkar, G. 2-Effect of climate change on agricultural crops. In Current Developments in Biotechnology and Bioengineering; Elsevier: Amsterdam, The Netherlands, 2017; pp. 23–46. ISBN 978-0-444-63661-4. [Google Scholar]
- Challinor, A.J.; Watson, J.; Lobell, D.B.; Howden, S.M.; Smith, D.R.; Chhetri, N. A meta-analysis of crop yield under climate change and adaptation. Nat. Clim. Chang. 2014, 4, 287–291. [Google Scholar] [CrossRef]
- Cerri, C.E.P.; Sparovek, G.; Bernoux, M.; Easterling, W.E.; Melillo, J.M.; Cerri, C.C. Tropical agriculture and global warming: Impacts and mitigation options. Sci. Agric. 2007, 64, 83–99. [Google Scholar] [CrossRef]
- Dixon, G.R. Climate change–mpact on crop growth and food production, and plant pathogens. Can. J. Plant. Pathol. 2012, 34, 362–379. [Google Scholar] [CrossRef]
- Chakraborty, S.; Newton, A.C. Climate change, plant diseases and food security: An overview. Plant. Pathol. 2011, 60, 2–14. [Google Scholar] [CrossRef]
- Ghini, R.; Bettiol, W.; Hamada, E. Diseases in tropical and plantation crops as affected by climate changes: Current knowledge and perspectives. Plant. Pathol. 2011, 60, 122–132. [Google Scholar] [CrossRef]
- Mesa-Sánchez, Ó.J.; Peñaranda-Vélez, V.M. Complejidad de la estructura espacio-temporal de la precipitación. Rev. Acad. Colomb. Cienc. Exactas Físicas Nat. 2015, 39, 304–320. [Google Scholar] [CrossRef] [Green Version]
- Mesa, O.; Poveda, G.; Carvajal, L. Introducción al Clima de Colombia; Universidad Nacional de Colombia: Medellín, Colombia, 1997; p. 390. ISBN 958-628-144-2. [Google Scholar]
- Pérez, C.; Poveda, G.; Mesa, O.; Carvajal, L.F.; Ochoa, A. Evidencias de cambio climático en Colombia: Tendencias y cambios de fase y amplitud de los ciclos anual y semianual. Bull. Inst. Fr. Détudes Andin. 1998, 27, 537–546. [Google Scholar]
- Poveda, G.; Pineda, K. Reassessment of Colombia’s tropical glaciers retreat rates: Are they bound to disappear during the 2010–2020 decade? Adv. Geosci. 2009, 22, 107–116. [Google Scholar] [CrossRef] [Green Version]
- Ruiz, D.; Poveda, G.; Vélez, I.D.; Quiñones, M.L.; Rúa, G.L.; Velásquez, L.E.; Zuluaga, J.S. Modelling entomological-climatic interactions of Plasmodium falciparum malaria transmission in two Colombian endemic-regions: Contributions to a National Malaria Early Warning System. Malar. J. 2006, 5, 66. [Google Scholar] [CrossRef]
- Capotondi, A.; Wittenberg, A.T.; Newman, M.; Di Lorenzo, E.; Yu, J.-Y.; Braconnot, P.; Cole, J.; Dewitte, B.; Giese, B.; Guilyardi, E.; et al. Understanding ENSO diversity. Bull. Am. Meteorol. Soc. 2014, 96, 921–938. [Google Scholar] [CrossRef]
- McPhaden, M.J.; Zebiak, S.E.; Glantz, M.H. ENSO as an integrating concept in earth science. Science 2006, 314, 1740–1745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iizumi, T.; Luo, J.-J.; Challinor, A.J.; Sakurai, G.; Yokozawa, M.; Sakuma, H.; Brown, M.E.; Yamagata, T. Impacts of El Niño Southern Oscillation on the global yields of major crops. Nat. Commun. 2014, 5, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minchio, C.A.; Canteri, M.G.; Fantin, L.H.; De Aguiar, E.S.M. Soybean rust epidemics in Rio Grande do Sul explained by ENSO phenomenon and off-season disease incidence. Summa Phytopathol. 2016, 42, 321–326. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Gil, J.G.; Morales-Osorio, J.G. Microbial dynamics in the soil and presence of the avocado wilt complex in plots cultivated with avocado cv. Hass under ENSO phenomena (El Niño-La Niña). Sci. Hortic. 2018, 240, 273–280. [Google Scholar] [CrossRef]
- Bost, J.B.; Smith, N.J.; Crane, J.H. History, distribution and uses. In The Avocado: Botany, Production and Uses, 2nd ed.; CABI: Wallingford, UK, 2013; pp. 10–30. [Google Scholar]
- Chen, H.; Morrell, P.L.; Ashworth, V.E.T.M.; de la Cruz, M.; Clegg, M.T. Tracing the geographic origins of major avocado cultivars. J. Hered. 2009, 100, 56–65. [Google Scholar] [CrossRef] [Green Version]
- Crane, J.H.; Douhan, G.; Faber, B.A.; Arpaia, M.L.; Bender, G.S.; Balerdi, C.F.; Barrientos-Priego, A.F. Cultivars and rootstocks. In The Avocado Botany, Production and Uses, 2nd ed.; CABI: Wallingford, UK, 2013; pp. 200–233. [Google Scholar]
- FAO. FAOSTAT. Available online: http://www.fao.org/faostat/es/#home (accessed on 9 January 2020).
- Rebolledo, A.; Romero, M.A. Avances en investigación sobre el comportamiento productivo del aguacate (Persea americana Mill.) bajo condiciones subtropicales. Cienc. Tecnol. Agropecu. 2011, 12, 113–120. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Gil, J.G.; Cobos, M.E.; Jiménez-García, D.; Morales-Osorio, J.G.; Peterson, A.T. Current and potential future distributions of Hass avocados in the face of climate change across the Americas. Crop. Pasture Sci. 2019, 70, 694–708. [Google Scholar] [CrossRef]
- Schaffer, B.; Gil, P.M.; Mickelbart, M.V.; Whiley, A.W. Ecophysiology. In The Avocado: Botany, Production and Uses, 2nd ed.; CABI: Wallingford, UK, 2013; pp. 168–199. [Google Scholar]
- Wolstenholme, B.N. Ecology: Climate and soils. In The Avocado: Botany, Production and Uses, 2nd ed.; CABI: Wallingford, UK, 2013; pp. 86–117. [Google Scholar]
- Ramírez-Gil, J.G.; Gilchrist Ramelli, E.; Morales Osorio, J.G. Economic impact of the avocado (cv. Hass) wilt disease complex in Antioquia, Colombia, crops under different technological management levels. Crop. Prot. 2017, 101, 103–115. [Google Scholar] [CrossRef]
- Ramírez-Gil, J.G.; Peterson, A.T. Current and potential distributions of most important diseases affecting Hass avocado in Antioquia Colombia. J. Plant. Prot. Res. 2019, 59, 214–228. [Google Scholar] [CrossRef]
- Hardham, A.R.; Blackman, L.M. Phytophthora cinnamomi. Mol. Plant. Pathol. 2018, 19, 260–285. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Gil, J.; Morales-Osorio, J.G. Polyphasic identification of preharvest pathologies and disorders in avocado cv. Hass. Agron. Colomb. 2019, 37, 213–227. [Google Scholar] [CrossRef]
- Ramírez-Gil, J. Avocado wilt complex disease, implications and management in Colombia. Rev. Fac. Nac. Agron. Medellín 2018, 71, 8525–8541. [Google Scholar] [CrossRef]
- Zentmyer, G. Phytophthora cinnamomi and diseases it causes. In Monograph, American Phytopathological Society; American Phytopathological Society: St. Paul, MN, USA, 1980; p. 96. [Google Scholar]
- Schaffer, B.; Davies, F.S.; Crane, J.H. Responses of subtropical and tropical fruit trees to flooding in calcareous soil. Am. Soc. Hort. Sci. 2006, 41, 549–555. [Google Scholar] [CrossRef] [Green Version]
- Gisi, U.; Zentmyer, G.A.; Klure, L.J. Production of sporangia by Phytophthora cinnamomi and P. palmivora in soils at different matric potentials. Phytopathology 1980, 70, 301–306. [Google Scholar] [CrossRef]
- Sterne, R.E.; Zentmyer, G.A.; Kaufmann, M.R. Effect of matric and osmotic potential of soil on Phytophthora root disease of Persea indica. Phytopathology 1977, 67, 1491–1494. [Google Scholar] [CrossRef] [Green Version]
- Castro, V.M.; Iturrieta, E.R.; Fassio, O.C. Rootstock effect on the tolerance of cv. Hass avocado plants to NaCl stress. Chil. J. Agric. Res. 2009, 69, 316–324. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Gil, J.G.; Castañeda-Sánchez, D.; Morales-Osorio, J. Production of avocado trees infected with Phytophthora cinnamomi under different management regimes. Plant. Pathol. 2017, 66, 623–632. [Google Scholar] [CrossRef]
- Deng, L.; Meng, X.; Yu, R.; Wang, Q. Assessment of the effect of mulch film on crops in the arid agricultural region of China under future climate scenarios. Water 2019, 11, 1819. [Google Scholar] [CrossRef] [Green Version]
- Iriany, A.; Chanan, M.; Djoyowasito, G. Organic mulch sheet formulation as an effort to help plants adapt to climate change. Int. J. Recycl. Org. Waste Agric. 2018, 7, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Ben-Ya’acov, A.; Michelson, E. Avocado rootstocks. Hortic. Rev. 1995, 17, 381–429. [Google Scholar]
- Zilberstaine, M.; Ben-Ya’acov, A.; Sela, I. Selection of avocado rootstocks resistant to root-rot in Israel. Proc. Interam. Soc. Trop. Hortic. 1992, 36, 15–19. [Google Scholar]
- Steward, P.R.; Dougill, A.J.; Thierfelder, C.; Pittelkow, C.M.; Stringer, L.C.; Kudzala, M.; Shackelford, G.E. The adaptive capacity of maize-based conservation agriculture systems to climate stress in tropical and subtropical environments: A meta-regression of yields. Agric. Ecosyst. Environ. 2018, 251, 194–202. [Google Scholar] [CrossRef]
- GalindoTovar, M.E.; Ogata Aguilar, N.; Alzate Fernández, A.M. Some aspects of avocado (Persea americana Mill.) diversity and domestication in Mesoamerica. Genet. Resour. Crop. Evol. 2008, 55, 441–450. [Google Scholar] [CrossRef]
- Frolich, E.; Schroeder, C.; Zentmyer, G. Graft compatibility in the genus Persea. Calif. Avocado Soc. Yearb. USA 1958, 42, 102–105. [Google Scholar]
- Ben-Ya’acov, A.; Frenkel, M. Sensitivity of avocado rootstocks to Verticillium wilt. Calif. Avocado Soc. 1973, 57, 111–113. [Google Scholar]
- Halma, F. Avocado rootstock experiment-a 10-year report. Calif. Avocado Soc. 1954, 38, 78–89. [Google Scholar]
- Khan, M.A.; Gemenet, D.C.; Villordon, A. Root system architecture and abiotic atress tolerance: Current knowledge in root and tuber crops. Front. Plant. Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- García-Pineda, E.; Benezer-Benezer, M.; Gutiérrez-Segundo, A.; Rangel-Sánchez, G.; Arreola-Cortés, A.; Castro-Mercado, E. Regulation of defence responses in avocado roots infected with Phytophthora cinnamomi (Rands). Plant. Soil 2010, 331, 45–56. [Google Scholar] [CrossRef]
- Phillips, D.; Grant, B.R.; Weste, G. Histological changes in the roots of an avocado cultivar, Duke 7, infected with Phytophthora cinnamomi. Phytopathology 1987, 77, 691–698. [Google Scholar] [CrossRef]
- Christie, J.B. Determining the Phenotypic Resistance Mechanisms in Avocado Against Phytophthora cinnamomi. Master’s Thesis, University of Pretoria, Pretoria, South Africa, 2012. [Google Scholar]
- Costa, J.D.S.; Menge, J.A.; Casale, W.L. Investigations on some of the mechanisms by which bioenhanced mulches can suppress Phytophthora root rot of avocado. Microbiol. Res. 1996, 151, 183–192. [Google Scholar] [CrossRef]
- Downer, J.; Faber, B.; Menge, J. Factors affecting root rot control in mulched avocado orchards. HortTechnology 2002, 12, 601–605. [Google Scholar] [CrossRef]
- Ramírez-Gil, J.G.; Martínez, G.O.G.; Morales Osorio, J.G. Design of electronic devices for monitoring climatic variables and development of an early warning system for the avocado wilt complex disease. Comput. Electron. Agric. 2018, 153, 134–143. [Google Scholar] [CrossRef]
- Ramírez-Gil, J.G.; Morales-Osorio, J.G. Development and validation of severity scales of avocado wilt complex caused by Phytophthora cinnamomi, Verticillium dahliae and hypoxia-anoxia disorder and their physiological responses in avocado plants. Agron. Colomb. 2020, 38. [Google Scholar] [CrossRef]
- Madden, L.V.; Hughes, G.; van den Bosch, F. The Study of Plant Disease Epidemics; American Phytopathological Society (APS Press): St. Paul, MN, USA, 2007; ISBN 978-0-89054-505-8. [Google Scholar]
- Hunt, R. Plant. Growth Analysis; Edward Arnold Publishers: London, UK, 1978; ISBN 0713128445. [Google Scholar]
- R Development Core Team. R: The R Project for Statistical Computing. Available online: https://www.r-project.org/ (accessed on 1 June 2020).
- Wider, R.K.; Lang, G.E. A Critique of the analytical methods used in examining decomposition data obtained from litter bags. Ecology 1982, 63, 1636–1642. [Google Scholar] [CrossRef] [Green Version]
- Richter, B.S.; Ivors, K.; Shi, W.; Benson, D.M. Cellulase Activity as a Mechanism for Suppression of Phytophthora Root Rot in Mulches. Phytopathology 2010, 101, 223–230. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, C. Aislamiento y Evaluación de Rizobacterias con Potencial Biocontrolador y Promotor de Crecimiento en Plantas en Banano. Master’s Thesis, Universidad Nacional de Colombia sede Medellín, Medellín, Colombia, 2005. [Google Scholar]
- Gams, W.; Bisset, J. Morphology and Identification of Trichoderma. In Trichodermaand Gliocladium: Basic Biology, Taxonomy and Genetics; Taylor & Francis: London, UK, 1998; pp. 3–34. [Google Scholar]
- Wood, P.J. Specificity in the interaction of direct dyes with polysaccharides. Carbohydr. Res. 1980, 85, 271–287. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.; Ryan, P. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Anderson, J.P.; Domsch, K.H. A physiological method for the quantitative measurement of microbial biomass in soils. Soil Biol. Biochem. 1978, 10, 215–221. [Google Scholar] [CrossRef]
- Instituto De Hidrología, Meteorología Y Estudios Ambientales IDEAM. Condiciones Hidroclimáticas Actuales y Predicción Climática para los Próximos. 2016. Available online: http://www.ideam.gov.co/documents/21021/548388/03_Condiciones+Clim%C3%A1ticas+Actuales_marzo+30+de+2016.pdf/18a91d72-8eec-40f6-964c-ca56eb6fa2de?version=1.0 (accessed on 4 February 2020).
- Lau, M.K. DTK: Dunnett-Tukey-Kramer Pairwise Multiple Comparison Test Adjusted for Unequal Variances and Unequal Sample Sizes. 2013. Available online: https://cran.r-project.org/web/packages/DTK/DTK.pdf (accessed on 20 March 2020).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype/Soil Moisture in Percentage (%) | 10 | 20 | 30 | 40 | 50 | 70 | 90 | 110 | 130 | 150 | ||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| RRI 1 | RRI 1 | RRI 1 | RRI 1 | RRI 1 | RRI 1 | RRI 1 | AUDPC 2 | RRI 1 | AUDPC 2 | RRI 1 | AUDPC 2 | RRI 1 | AUDPC 2 | |
| AX1 | 0.004 c | 0.008 c | 0.015 c | 0.023 c | 0.047 c | 0.032 c | 0.059 a | 12.5 c | 0.04 b | 35.4 e | 0.025 a | 95.4 c | 0.006 a | 200.3 c |
| AX2 | 0.003 d | 0.009 c | 0.016 bc | 0.024 c | 0.050 b | 0.034 c | 0.05 b | 10.1 c | 0.035 b | 30.2 e | 0.021 a | 88.9 c | 0.007 a | 185.4 c |
| AX3 | 0.005 c | 0.010 c | 0.017 b | 0.025 c | 0.055 b | 0.038 b | 0.065 a | 19.3 bc | 0.052 a | 60.3 d | 0.018 ab | 150.4 b | 0.005 a | 230.4 bc |
| AX4 | 0.005 c | 0.011 c | 0.019 b | 0.026 bc | 0.059 ab | 0.04 b | 0.069 a | 7.5.6 c | 0.049 a | 25.8 de | 0.02 ab | 70.3 c | 0.006 a | 165.6 bc |
| AX5 | 0.002 d | 0.008 c | 0.014 c | 0.022 c | 0.045 c | 0.031 c | 0.045 c | 13.8 c | 0.038 b | 50.3 d | 0.015 b | 100.3 c | 0.006 a | 205.8 c |
| GX1 | 0.006 b | 0.013 b | 0.024 a | 0.031 a | 0.065 a | 0.047 ab | 0.050 b | 25.6 b | 0.028 c | 75.4 c | 0.014 b | 160.3 b | 0.001 b | 225 bc |
| GX2 | 0.012 a | 0.025 a | 0.029 a | 0.032 a | 0.066 a | 0.045 b | 0.045 c | 28.4 b | 0.025 c | 89.3 bc | 0.009 c | 185.4 ab | 0.001 b | 250.3 b |
| GX3 | 0.010 a | 0.020 a | 0.027 a | 0.034 a | 0.070 a | 0.052 a | 0.055 b | 20.3 bc | 0.03 c | 80.2 c | 0.008 c | 160 b | 0.001 b | 230.4 bc |
| GX4 | 0.007 b | 0.013 b | 0.026 a | 0.031 a | 0.061 ab | 0.045 b | 0.047 c | 32.4 b | 0.023 d | 100.5 b | 0.011 c | 205.3 a | 0.0009 b | 301.2 a |
| MX1 | 0.008 a | 0.014 b | 0.020 b | 0.023 c | 0.042 c | 0.025 d | 0.032 d | 39.1 a | 0.028 c | 115.8 a | 0.007 c | 215.6 a | 0.0008 b | 320.5 a |
| MX2 | 0.006 b | 0.015 b | 0.018 b | 0.026 bc | 0.045 c | 0.027 d | 0.035 d | 42.3 a | 0.024 cd | 120.3 a | 0.006 c | 225.4 a | 0.006 b | 335.1 a |
| Hass | 0.007 b | 0.013 b | 0.017 b | 0.029 b | 0.052 b | 0.035 c | 0.038 d | 32.1 b | 0.027 c | 100 b | 0.007 c | 190.3 a | 0.0009 b | 250.2 b |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramírez-Gil, J.G.; Henao-Rojas, J.C.; Morales-Osorio, J.G. Mitigation of the Adverse Effects of the El Niño (El Niño, La Niña) Southern Oscillation (ENSO) Phenomenon and the Most Important Diseases in Avocado cv. Hass Crops. Plants 2020, 9, 790. https://doi.org/10.3390/plants9060790
Ramírez-Gil JG, Henao-Rojas JC, Morales-Osorio JG. Mitigation of the Adverse Effects of the El Niño (El Niño, La Niña) Southern Oscillation (ENSO) Phenomenon and the Most Important Diseases in Avocado cv. Hass Crops. Plants. 2020; 9(6):790. https://doi.org/10.3390/plants9060790
Chicago/Turabian StyleRamírez-Gil, Joaquín Guillermo, Juan Camilo Henao-Rojas, and Juan Gonzalo Morales-Osorio. 2020. "Mitigation of the Adverse Effects of the El Niño (El Niño, La Niña) Southern Oscillation (ENSO) Phenomenon and the Most Important Diseases in Avocado cv. Hass Crops" Plants 9, no. 6: 790. https://doi.org/10.3390/plants9060790