More than Scales: Evidence for the Production and Exudation of Mucilage by the Peltate Trichomes of Tillandsia cyanea (Bromeliaceae: Tillandsioideae)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Secretory Activity and Aspects of the Secretion
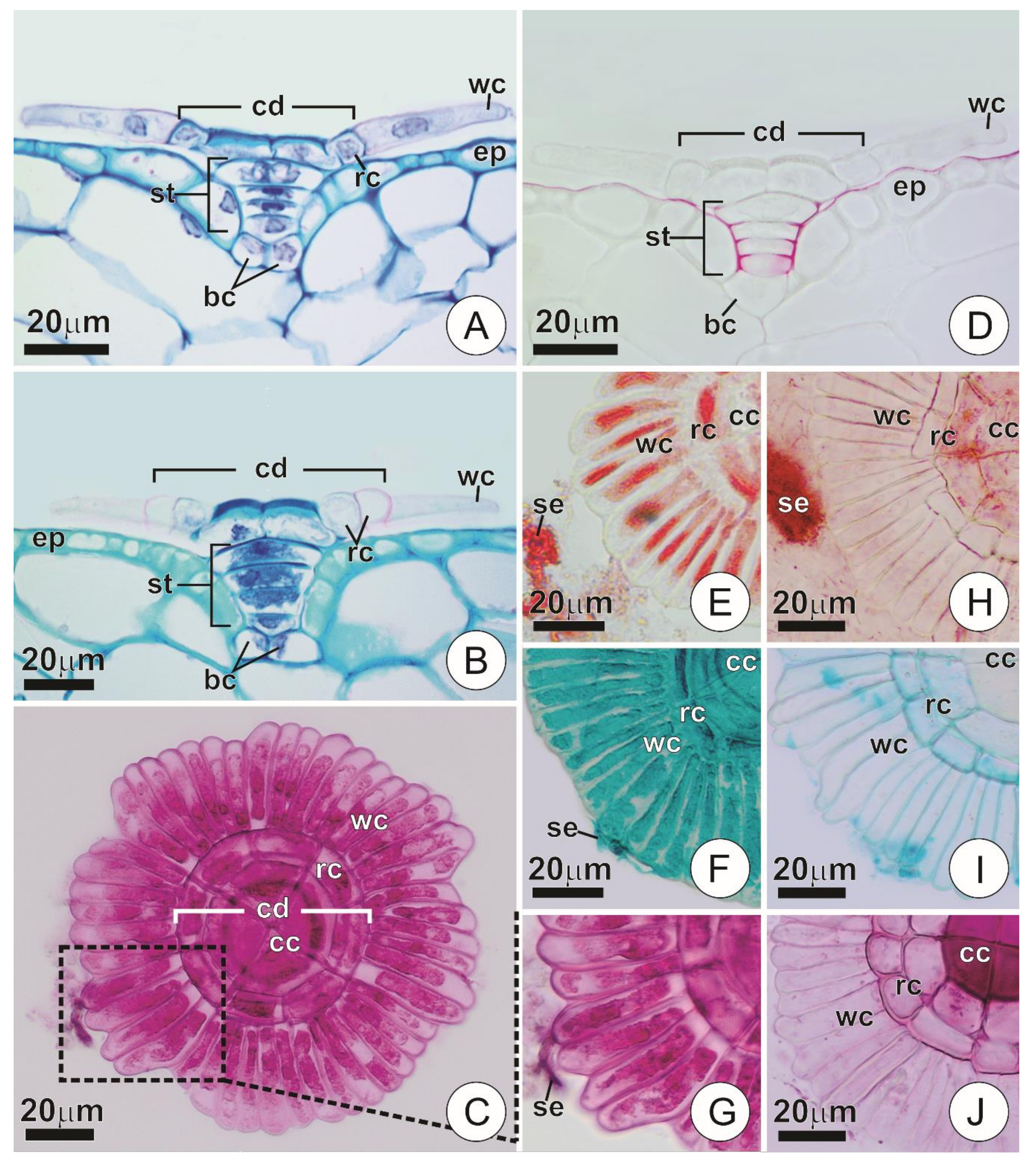
2.2. Anatomy and Histochemistry of the Secretory Trichomes
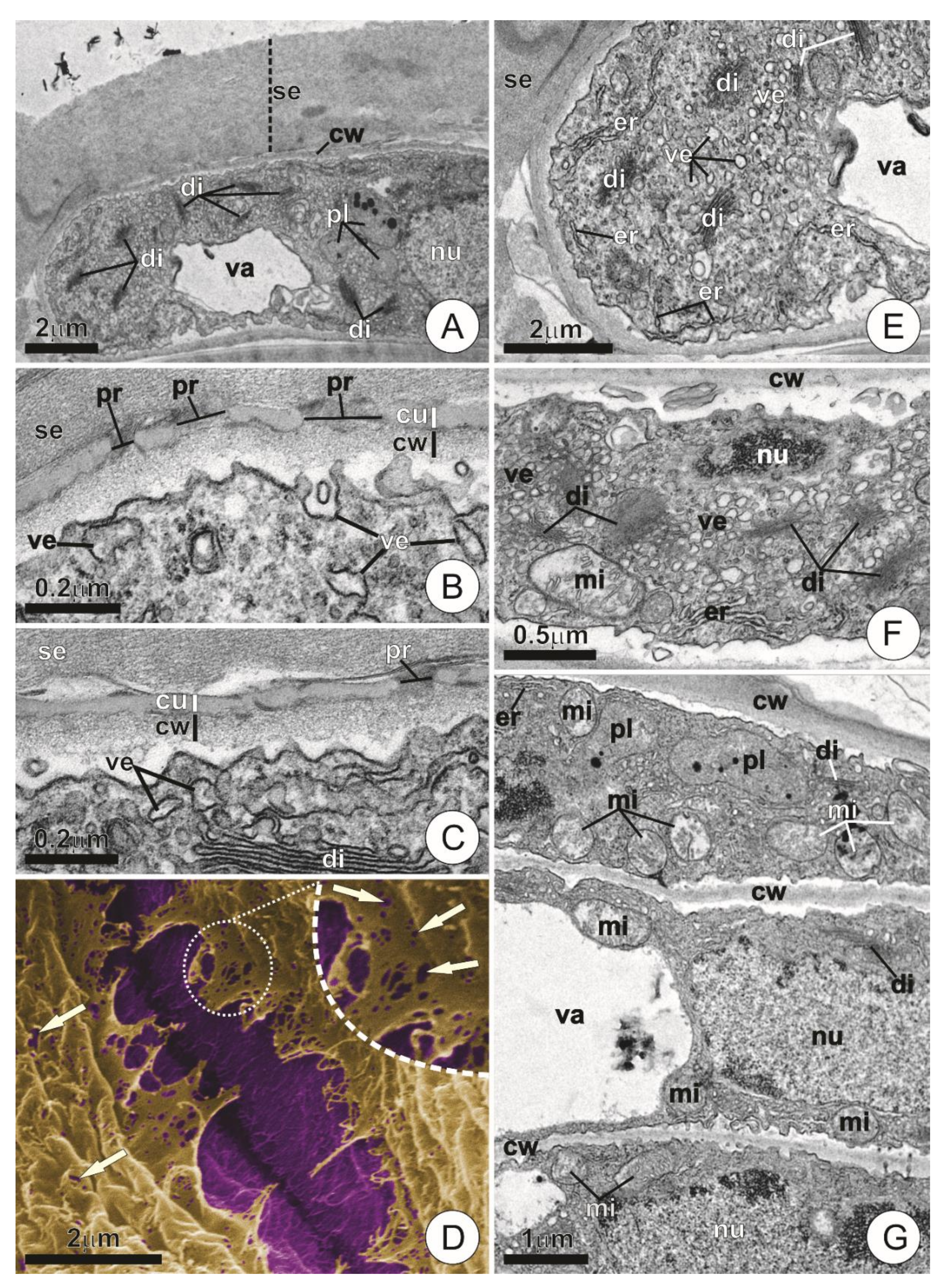
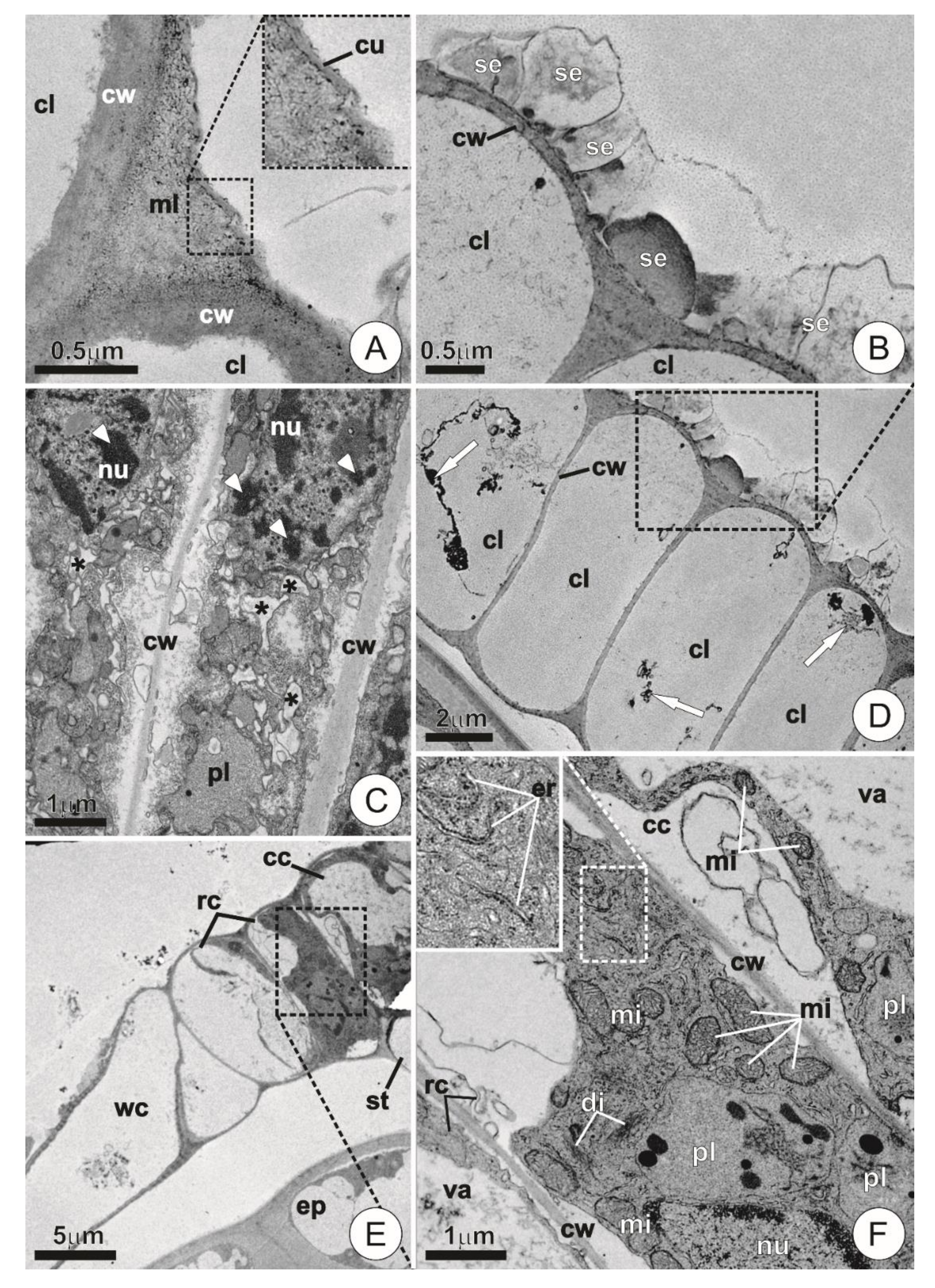
2.3. Ultrastructure
3. Discussion
3.1. Trichome Structure, Distribution, and Activity
3.2. Ultrastructure and Secretory Mechanisms
3.3. Functional Aspects
4. Materials and Methods
4.1. Plant Material and Sampling
4.2. Light Microscopy
4.3. Electron Microscopy
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Schimper, A.F.W. Die epiphytische Vegetation Amerikas. Bot. Mitt. Trop. 1888, 2, 162. [Google Scholar]
- Mez, C. Physiologische Bromeliaceen-Studien. I. Die Wasser—Okonomie der extrem atmosphärischen Tillandsien. Jahrb. Wiss. Bot. 1904, 40, 157–229. [Google Scholar]
- Tietze, M. Physiologische Bromeliaceen Studien II. Die Entwicklung der wasseraufnehmenden Bromeliaceen Trichome. Z. Naturwiss. 1906, 78, 1–50. [Google Scholar]
- Wetzel, K. Beitrag zur Anatomie der Saughaare von Bromeliaceen. Flora Oder Allg. Bot. Zeitung 1924, 117, 133–143. [Google Scholar] [CrossRef]
- Dolzmann, P. Elektronenmikroskopische Untersuchungen an den Saughaaren von Tillandsia Usneoides (Bromeliaceae)—I. Feinstruktur der Kuppelzelle. Planta 1964, 60, 461–472. [Google Scholar] [CrossRef]
- Dolzmann, P. Elektronenmikroskopische untersuchungen an den saughaaren von Tillandsia usneoides (Bromeliaceae): II—Einige beobachtungen zur feinstruktur der plasmodesmen. Planta 1965, 64, 76–80. [Google Scholar] [CrossRef]
- Tomlinson, P.B. Anatomy of the Monocotyledons: III Commelinales-Zingiberales; Claredon Press: Oxford, UK, 1969. [Google Scholar]
- Benzing, D.H. Bromeliad trichomes: Structure, function, and ecological significance. J. Marie Selby Bot. Gard. 1976, 1, 330–348. [Google Scholar]
- Benzing, D.H.; Henderson, K.; Kessel, B.; Sulak, J. The absortive capacities of bromeliad trichomes. Am. J. Bot. 1976, 63, 1009–1014. [Google Scholar] [CrossRef]
- Benzing, D.H.; Givnish, T.J.; Bermudes, D. Absorptive trichomes in Brocchinia reducta (Bromeliaceae) and their evolutionary and systematic significance. Syst. Bot. 1985, 10, 81–91. [Google Scholar] [CrossRef]
- Pierce, S.; Maxwell, K.; Griffiths, H.; Winter, K. Hydrophobic trichome layers and epicuticular wax powders in Bromeliaceae. Am. J. Bot. 2001, 88, 1371–1389. [Google Scholar] [CrossRef] [Green Version]
- Ohrui, T.; Nobira, H.; Sakata, Y.; Taji, T.; Yamamoto, C.; Nishida, K.; Yamakawa, T.; Sasuga, Y.; Yaguchi, Y.; Takenaga, H.; et al. Foliar trichome and aquaporin-aided water uptake in a drought-resistant epiphyte Tillandsia ionantha Planchon. Planta 2007, 227, 47–56. [Google Scholar] [CrossRef]
- Papini, A.; Tani, G.; Di Falco, P.; Brighigna, L. The ultrastructure of the development of Tillandsia (Bromeliaceae) trichome. Flora 2010, 205, 94–100. [Google Scholar] [CrossRef]
- Kowalski, V.K.; Pereira, P.P.D.A.; De Oliveira, F.M.C.; Costa, M.E.; Tardivo, R.C. Are the wings cells alive? Study case in Vriesea trichomes. Rodriguesia 2016, 67, 427–435. [Google Scholar] [CrossRef] [Green Version]
- Kleingesinds, C.K.; Gobara, B.N.K.; Mancilha, D.; Rodrigues, M.A.; Demarco, D.; Mercier, H. Impact of tank formation on distribution and cellular organization of trichomes within Guzmania monostachia rosette. Flora 2018, 243, 11–18. [Google Scholar] [CrossRef]
- Ballego-Campos, I.; Paiva, E.A.S. Mucilage secretion in the inflorescences of Aechmea blanchetiana: Evidence of new functions of scales in Bromeliaceae. Flora 2018, 246–247, 1–9. [Google Scholar] [CrossRef]
- Benzing, D.H. Foliar permeability and the absorption of minerals and organic nitrogen by certain tank bromeliads. Bot. Gaz. 1970, 131, 23–31. [Google Scholar] [CrossRef]
- Benzing, D.H. Bromeliaceae: Profile of an Adaptive Radiation; Cambridge University Press: New York, NY, USA, 2000. [Google Scholar]
- Givnish, T.J.; Burkhardt, E.L.; Happel, R.E.; Weintraub, J.D. Carnivory in the bromeliad Brocchinia reducta, with a cost/benefit model for the general restriction of carnivorous plants to sunny, moist, nutrient-poor habitats. Am. Nat. 1984, 124, 479–497. [Google Scholar] [CrossRef]
- Sakai, W.S.; Sanford, W.G. Ultrastructure of the water-absorbing trichomes of pineapple (Ananas comosus, Bromeliaceae). Ann. Bot. 1980, 46, 7–11. [Google Scholar] [CrossRef]
- Brighigna, L.; Palandri, M.R.; Giuffrida, M.; Macchi, C.; Tani, G. Ultrastructural features of the Tillandsia usneoides L. absorbing trichome during conditions moisture and aridity. Caryologia 1988, 41, 111–129. [Google Scholar] [CrossRef]
- Owen, T.P.; Thomson, W.W. Structure and function of a specialized cell-wall in the trichomes of the carnivorous bromeliad Brocchinia reducta. Can. J. Bot. 1991, 69, 1700–1706. [Google Scholar] [CrossRef]
- Owen, T.P. Apoplastic and ultrastructural characterizations of the trichomes from the carnivorous bromeliad Brocchinia reducta. Can. J. Bot. 1988, 66, 941–948. [Google Scholar] [CrossRef]
- Pierce, S. The jeweled armor of Tillandsia—Multifaceted or elongated trichomes provide photoprotection. Aliso 2007, 23, 44–52. [Google Scholar] [CrossRef] [Green Version]
- Gouda, E.J.; Butcher, D. A List of Accepted Bromeliaceae Names. Available online: http://bromeliad.nl/bromNames/ (accessed on 20 February 2020).
- Givnish, T.J.; Millam, K.C.; Berry, P.E.; Sytsma, K.J. Phylogeny, adaptive radiation, and historical biogeography of bromeliaceae inferred from ndhF sequence data. Aliso 2007, 23, 3–26. [Google Scholar] [CrossRef]
- Givnish, T.J.; Barfuss, M.H.J.; van Ee, B.; Riina, R.; Schulte, K.; Horres, R.; Gonsiska, P.A.; Jabaily, R.S.; Crayn, D.M.; Smith, J.A.C.; et al. Phylogeny, adaptive radiation, and historical biogeography in Bromeliaceae: Insights from an eight-locus plastid phylogeny. Am. J. Bot. 2011, 98, 872–895. [Google Scholar] [CrossRef] [Green Version]
- Barfuss, M.H.J.; Till, W.; Leme, E.M.C.; Pinzón, J.P.; Manzanares, J.M.; Halbritter, H.; Samuel, R.; Brown, G.K. Taxonomic revision of bromeliaceae subfam. Tillandsioideae based on a multi-locus DNA sequence phylogeny and morphology. Phytotaxa 2016, 279. [Google Scholar] [CrossRef]
- Benzing, D.H.; Seemann, J.; Renfrow, A. The foliar epidermis in Tillandsioideae (Bromeliaceae) and its role in habitat selection. Am. J. Bot. 1978, 65, 359. [Google Scholar] [CrossRef]
- Givnish, T.J.; Barfuss, M.H.J.; Ee, B.V.; Riina, R.; Schulte, K.; Horres, R.; Gonsiska, P.A.; Jabaily, R.S.; Crayn, D.M.; Smith, J.A.C.; et al. Adaptive radiation, correlated and contingent evolution, and net species diversification in Bromeliaceae. Mol. Phylogenet. Evol. 2014, 71, 55–78. [Google Scholar] [CrossRef]
- Mayer, J.L.S.; Cardoso-Gustavson, P.; Appezzato-da-Glória, B. Colleters in monocots: New record for Orchidaceae. Flora 2011, 206, 185–190. [Google Scholar] [CrossRef]
- Mayer, J.L.S.; Carmello-Guerreiro, S.M.; Mazzafera, P. A functional role for the colleters of coffee flowers. AoB Plants 2013, 5, 1–13. [Google Scholar] [CrossRef]
- Oriani, A.; Scatena, V.L. The taxonomic value of floral characters in Rapateaceae (Poales-Monocotyledons). Plant. Syst. Evol. 2013, 299, 291–303. [Google Scholar] [CrossRef]
- Ferrari, R.C.; Oriani, A. Floral anatomy and development of Saxofridericia aculeata (Rapateaceae) and its taxonomic and phylogenetic significance. Plant. Syst. Evol. 2017, 303, 187–201. [Google Scholar] [CrossRef] [Green Version]
- Ballego-Campos, I.; Paiva, E.A.S. Colleters in the vegetative axis of Aechmea blanchetiana (Bromeliaceae): Anatomical, ultrastructural and functional aspects. Aust. J. Bot. 2018, 66, 379–387. [Google Scholar] [CrossRef]
- Thomas, V. Structural, functional and phylogenetic aspects of the colleter. Ann. Bot. 1991, 68, 287–305. [Google Scholar] [CrossRef]
- Leitão, C.A.E.; Cortelazzo, A.L. Structural and histochemical characterization of the colleters of Rodriguezia venusta (Orchidaceae). Aust. J. Bot. 2008, 56, 161–165. [Google Scholar] [CrossRef]
- Cardoso-Gustavson, P.; Campbell, L.M.; Mazzoni-Viveiros, S.C.; De Barros, F. Floral colleters in Pleurothallidinae (Epidendroideae: Orchidaceae). Am. J. Bot. 2014, 101, 587–597. [Google Scholar] [CrossRef]
- Cassola, F.; Nunes, C.E.P.; Lusa, M.G.; Garcia, V.L.; Mayer, J.L.S. Deep in the jelly: Histochemical and functional aspects of mucilage-secreting floral colleters in the orchids Elleanthus brasiliensis and E. crinipes. Front. Plant. Sci. 2019, 10, 518. [Google Scholar] [CrossRef]
- Bernardello, M.; Galleto, L.; Juliani, H.R. Floral nectar, nectary structure and pollinators in some argentinean Bromeliaceae. Ann. Bot. 1991, 67, 401–411. [Google Scholar] [CrossRef]
- Vesprini, J.L.; Galetto, L.; Bernardello, G. The beneficial effect of ants on the reproductive success of Dyckia floribunda (Bromeliaceae), an extrafloral nectary plant. Can. J. Bot. 2003, 81, 24–27. [Google Scholar] [CrossRef]
- Mosti, S.; Friedman, C.R.; Pacini, E.; Brighigna, L.; Papini, A. Nectary ultrastructure and secretory modes in three species of tillandsia (Bromeliaceae) that have different pollinators. Botany 2013, 91, 786–798. [Google Scholar] [CrossRef]
- Smith, L.B.; Downs, R.J. Tillandsioideae (Bromeliaceae). Flora Neotrop. 1977, 14, 663–1492. [Google Scholar]
- Vervaeke, I.; Wouters, J.; Londers, E.; De Proft, M.P.; Deroose, R. Morphology of artificial hybrids of Vriesea splendens x Tillandsia cyanea and V. splendens x Guzmania lingulata (Bromeliaceae). Ann. Bot. Fenn. 2004, 41, 201–208. [Google Scholar]
- Paiva, E.A.S. Occurrence, structure and functional aspects of the colleters of Copaifera langsdorffii Desf. (Fabaceae, Caesalpinioideae). C. R. Biol. 2009, 332, 1078–1084. [Google Scholar] [CrossRef] [PubMed]
- Paiva, E.A.S. Colleters in Cariniana estrellensis (Lecythidaceae): Structure, secretion and evidences for young leaf protection. J. Torrey Bot. Soc. 2012, 139, 1–8. [Google Scholar] [CrossRef]
- Mercadante-Simões, M.O.; Paiva, E.A.S. Leaf colleters in Tontelea micrantha (Celastraceae, Salacioideae): Ecological, morphological and structural aspects. C. R. Biol. 2013, 336, 400–406. [Google Scholar] [CrossRef]
- Fernandes, V.F.; Thadeo, M.; Dalvi, V.C.; Meira, R.M.S.A. Secretory structures in Casearia sylvestris sw. (Salicaceae): Diversity, mechanisms of secretion, and exudate complexity. Int. J. Plant. Sci. 2017, 178, 288–301. [Google Scholar] [CrossRef]
- Da Silva, C.J.; Barbosa, L.C.D.A.; Marques, A.E.; Baracat-Pereira, M.C.; Pinheiro, A.L.; Meira, R.M.S.A. Anatomical characterisation of the foliar colleters in Myrtoideae (Myrtaceae). Aust. J. Bot. 2012, 60, 707–717. [Google Scholar] [CrossRef]
- De Barros, T.C.; Marinho, C.R.; Pedersoli, G.D.; Paulino, J.V.; Teixeira, S.P. Beyond pollination: Diversity of secretory structures during flower development in different legume lineages. Acta Bot. Bras. 2017, 31, 358–373. [Google Scholar] [CrossRef] [Green Version]
- Machado, S.R.; Paleari, L.M.; Paiva, É.A.S.; Rodrigues, T.M. Colleters on the inflorescence Axis of Croton glandulosus (Euphorbiaceae): Structural and functional characterization. Int. J. Plant. Sci. 2015, 176, 86–93. [Google Scholar] [CrossRef] [Green Version]
- Almeida, A.L.; Paiva, E.A.S. Colleters in Mabea fistulifera Mart. (Euphorbiaceae): Anatomy and biology of the secretory process. Flora 2019, 258, 151439. [Google Scholar] [CrossRef]
- Mohan, J.S.S.; Inamdar, J.A. Ultrastructure and secretion of extrafloral nectaries of Plumeria rubra L. Ann. Bot. 1986, 57, 389–401. [Google Scholar] [CrossRef]
- Fahn, A. Secretory Tissue in Plants; Academic Press: London, UK, 1979. [Google Scholar]
- Fahn, A. Structure and function of secretory cells. Adv. Bot. Res. 2000, 31, 37–75. [Google Scholar]
- Oliveira, C.S.; Salino, A.; Paiva, E.A.S. Colleters in Thelypteridaceae: Unveiling mucilage secretion and its probable role in ferns. Flora 2017, 228, 65–70. [Google Scholar] [CrossRef]
- Paiva, E.A.S. How does the nectar of stomata-free nectaries cross the cuticle? Acta Bot. Bras. 2017, 31, 525–530. [Google Scholar] [CrossRef] [Green Version]
- Antoń, S.; Kamińska, M. Comparative floral spur anatomy and nectar secretion in four representatives of Ranunculaceae. Protoplasma 2015, 252, 1587–1601. [Google Scholar] [CrossRef] [Green Version]
- Weryszko-Chmielewska, E.; Chwil, M. Flowering biology and structure of floral nectaries in Galanthus nivalis L. Acta Soc. Bot. Pol. 2016, 85, 3486. [Google Scholar] [CrossRef] [Green Version]
- Findlay, N.; Mercer, F.V. Nectar production in Abutilon I. Movement of nectar through the cuticle. Aust. J. Biol. Sci. 1971, 24, 647–656. [Google Scholar] [CrossRef] [Green Version]
- Jeffree, C.E. The Fine Structure of the Plant Cuticle. In Annual Plant Reviews; Riederer, M., Müller, C., Eds.; Blackwell Publishing: Oxford, UK, 2006; Volume 23, pp. 11–125. [Google Scholar]
- Papini, A.; Mosti, S.; Brighigna, L. Programmed-cell-death events during tapetum development of angiosperms. Protoplasma 1999, 207, 213–221. [Google Scholar] [CrossRef]
- Bakeeva, L.E.; Dzyubinskaya, E.V.; Samuilov, V.D. Programmed cell death in plants: Ultrastructural changes in pea guard cells. Biochemistry 2005, 70, 972–979. [Google Scholar] [CrossRef]
- Nobel, P.S.; Cavelier, J.; Andrade, J.L. Mucilage in cacti: Its apoplastic capacitance, associated solutes, and influence on tissue 5. J. Exp. Bot. 1992, 43, 641–648. [Google Scholar] [CrossRef]
- Chatchawal, C.; Nualkaew, N.; Preeprame, S.; Supatra Porasuphatana, A.P. Physical and biological properties of mucilage from Basella alba L. Stem and its gel formulation. Isan J. Pharm. Sci. 2010, 6, 104–112. [Google Scholar]
- Western, T.L. The sticky tale of seed coat mucilages: Production, genetics, and role in seed germination and dispersal. Seed Sci. Res. 2012, 22, 1–25. [Google Scholar] [CrossRef]
- Tresmondi, F.; Canaveze, Y.; Guimarães, E.; Machado, S.R. Colleters in Rubiaceae from forest and savanna: The link between secretion and environment. Sci. Nat. 2017, 104, 17. [Google Scholar] [CrossRef] [PubMed]
- Monteiro, R.F.; Macedo, M.V. First report on the diversity of insects trapped by a sticky exudate of the inflorescences of Vriesea bituminosa Wawra (Bromeliaceae: Tillandsioideae). Arthropod. Plant. Interact. 2014, 8, 519–523. [Google Scholar] [CrossRef]
- Paiva, E.A.S.; Martins, L.C. Calycinal trichomes in Ipomoea cairica (Convolvulaceae): Ontogenesis, structure and functional aspects. Aust. J. Bot. 2011, 59, 91–98. [Google Scholar] [CrossRef]
- Klein, D.E.; Gomes, V.M.; Da Silva-Neto, S.J.; Da Cunha, M. The structure of colleters in several species of Simira (Rubiaceae). Ann. Bot. 2004, 94, 733–740. [Google Scholar] [CrossRef] [Green Version]
- Miguel, E.C.; Moreira Gomes, V.; De Oliveira, M.A.; Da Cunha, M. Colleters in Bathysa nicholsonii K. Schum. (Rubiaceae): Ultrastructure, secretion protein composition, and antifungal activity. Plant. Biol. 2006, 8, 715–722. [Google Scholar] [CrossRef]
- Karnovsky, M.J. A formaldehyde-glutaraldehyde fixative of high osmolarity for use in electron microscopy. J. Cell Biol. 1965, 27, 137–138A. [Google Scholar]
- O’Brien, T.P.; Feder, N.; McCully, M.E. Polychromatic staining of plant cell walls by toluidine blue O. Protoplasma 1964, 59, 368–373. [Google Scholar] [CrossRef]
- Johansen, D.A. Plant Microtechnique; McGraw-Hill Book Co.: New York, NY, USA; London, UK, 1940. [Google Scholar]
- Pearse, A. Histochemistry: Theoretical and Applied, 4th ed.; Longman Group Limited: London, UK, 1968. [Google Scholar]
- Brundrett, M.C.; Kendrick, B.; Peterson, C.A. Efficient lipid staining in plant material with Sudan red 7B or fluoral yellow 088 in polyethylene glycol-glycerol. Biotech. Histochem. 1991, 66, 111–116. [Google Scholar] [CrossRef]
- Vidal, B.C. Dichroism in collagen bundles stained with Xylidine-Ponceau 2R. Ann. D’histochim. 1970, 15, 289–296. [Google Scholar]
- David, R.; Carde, J. Coloration différentialle des pseudophylles de Pin maritime au moyen du reactif de Nadi. C. R. Hebd. Seanc. Acad. Sci. 1964, 258, 1338–1340. [Google Scholar]
- Furr, M.; Mahlberg, P.G. Histochemical analyses of laticifers and glandular trichomes in Cannabis sativa. J. Nat. Prod. 1981, 44, 153–159. [Google Scholar] [CrossRef]
- Spurr, A.R. A low-viscosity epoxy resin embedding medium for electron microscopy. J. Ultrastruct. Res. 1969, 26, 31–43. [Google Scholar] [CrossRef]
- Roland, J.C. General preparation and staining of thin sections. In Electron Microscopy of Plant Cells; Hall, J., Ed.; Elsevier: New York, NY, USA, 1978; pp. 1–62. [Google Scholar]
- Watson, M.L. Staining of tissue sections for electron microscopy with heavy metals. J. Cell Biol. 1958, 4, 475–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, E.S. The use of lead citrate at high pH as an electron-opaque stain in electron microscopy. J. Cell Biol. 1963, 17, 208–212. [Google Scholar] [CrossRef] [Green Version]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ballego-Campos, I.; Forzza, R.C.; Paiva, E.A.S. More than Scales: Evidence for the Production and Exudation of Mucilage by the Peltate Trichomes of Tillandsia cyanea (Bromeliaceae: Tillandsioideae). Plants 2020, 9, 763. https://doi.org/10.3390/plants9060763
Ballego-Campos I, Forzza RC, Paiva EAS. More than Scales: Evidence for the Production and Exudation of Mucilage by the Peltate Trichomes of Tillandsia cyanea (Bromeliaceae: Tillandsioideae). Plants. 2020; 9(6):763. https://doi.org/10.3390/plants9060763
Chicago/Turabian StyleBallego-Campos, Igor, Rafaela C. Forzza, and Elder A. S. Paiva. 2020. "More than Scales: Evidence for the Production and Exudation of Mucilage by the Peltate Trichomes of Tillandsia cyanea (Bromeliaceae: Tillandsioideae)" Plants 9, no. 6: 763. https://doi.org/10.3390/plants9060763