1. Introduction
Cotton (
Gossypium spp.) is an economically important fibre crop contributing to approximately 40% of the world’s natural fibre [
1]. Cultivated cotton can be found in a range of ecological niches from arid to semi-arid areas of the tropical and subtropical zones and is mainly derived from Upland cotton (
G. hirsutum L.) and Pima cotton (
G. barbadense L.) due to superior fibre quality and quantity traits [
2]. In Australia, Upland cotton is a major agricultural crop and mainly grown in regional areas of New South Wales (NSW) and Queensland and produced predominantly under irrigated conditions, but smaller areas of dryland production do occur in some seasons [
3]. In the 2017/18 cotton growing season, the industry employed up to 10,000 people across 152 communities and contributed significantly to the economic growth and wealth of these regions. The majority of Australian cotton is exported, which generates an average value of around AUD 1.9 billion annually [
3].
In Australia, cotton is prone to infection with an array of pathogens, including black root rot pathogen
Thielaviopsis basicola (Berk. and Br.) Ferraris [
4], leaf spot pathogen
Alternaria alternata (Fr.) Keissl. [
5], Fusarium wilt pathogen
Fusarium oxysporum f. sp.
vasinfectum Snyder and Hansen, [
6,
7], and Verticillium wilt (VW) pathogen
Verticillium dahliae Kleb. [
8]. Of these, VW of cotton is of a major constraint to the sustainability of the cotton industry due to the lack of complete resistant resources and highly effective management strategies. The disease is associated with vascular discolouration, leaf chlorosis, necrosis, defoliation and plant death in some severe cases [
9]. In NSW, VW incidence in cotton was as little as 3% up to 16% on average between 1984 and 2012 [
8]. During 2016–2019, the highest average incidence of VW in NSW and Queensland was 30% and 4%, respectively [
10]. According to Allen [
11], yield loss caused by the disease in Australia can be up to 25% in years where climatic conditions favoured disease development. On a rare occasion, yield loss was estimated up to 50% in a severely infested field in NSW (unpublished data).
Verticillium dahliae is a soilborne phytopathogen and commonly associated with vascular wilt diseases of up to 400 host plant species; many of these are of economic importance in agriculture, horticulture and forestry [
12].
V. dahliae was reported across many cotton growing regions such as Australia, China, Spain and the USA [
8,
13,
14,
15]. Virulence of
V. dahliae was commonly found associated with its pathotypes, that being defoliating (D) and non-defoliating (ND), based on symptoms induced on host plants [
16,
17]. Unless otherwise stated, the D and ND pathotypes should only be referred to as pathotypes of
V. dahliae in this study. The D pathotype was deemed to be highly aggressive, inciting defoliation, and was lethal to cotton; alternatively, the ND pathotype was considered less aggressive and did not attribute to defoliation [
13]. However,
V. dahliae isolates recovered from Australian cotton and designated as D and ND pathotypes were found to be equally lethal to its host (unpublished data). In the past two survey seasons, the ND pathotype was detected across NSW, while the D pathotype was more prevalent in the northern valleys of NSW [
10]. Additionally, our initial data also indicated the presence of both pathotypes in some fields in NSW (unpublished data). Co-occurrence of both pathotypes in a cotton field were previously reported in Spain [
16]. Similarly, Jiménez-Díaz et al. [
18] found that co-occurrence of both pathotypes was relatively common in olive orchards in Spain. Interestingly, Mercado-Blanco et al. [
19] for the first time demonstrated that co-infection of D and ND pathotypes also occurred naturally in olive trees. This reflects the complexity of the patho-system in
V. dahliae. However, many used single isolates of
V. dahliae for biological and pathogenicity assessments [
20,
21].
Interactions between
V. dahliae with either other
Verticillium spp. or different isolates were assessed in artificially co-inoculated assays. The observed effects varied from none to cross-protection, depending on the inoculation methods, orders and time intervals between inoculations [
22,
23]. For example, VW expressions on lettuce were lower in co-inoculated plants with isolates of
V. tricorpus and
V. dahliae compared with those inoculated with
V. dahliae alone. Additionally, the co-inoculation relatively promoted the growth of lettuce [
23]. Qin et al. [
23] also reported that a soil drench with
V. tricorpus in advance of the inoculation with
V. dahliae appeared to provide better protection compared with simultaneous inoculation using a root-dip method. On the other hand, simultaneous inoculation of the avirulent isolate P6 and the highly virulent isolate VM of
V. dahliae did not result in any difference in disease expression on sunflower; however, sequential inoculation of the isolate VM two days following the challenge with the isolate P6 resulted in a significantly lower disease severity in sunflower [
24]. Shittu et al. [
25] found that disease scores in tomato were significantly lower in plants either preceding inoculation with the non-host isolate Dvd-E6 followed by the virulent isolate Vd1 of
V. dahliae or in simultaneous inoculation. Wheeler and Johnson [
22] reported that co-inoculation with two or three different host-selective
V. dahliae isolates did not alter potato yields, or mustard and barley biomass, but disease severity in potato increased in co-inoculated plants. On cotton, cross-protection was observed in plants grown both under artificially co-inoculated and naturally infested soils with the virulent D pathotype isolate T-1 and mildly virulent ND pathotype isolate SS-4 [
26]. However, at the time, there was no evidence that supported the co-existence of both pathotypes in cotton plants [
26]. To date, only Mercado-Blanco et al. [
19] showed the natural co-infection and co-existence of the D and ND pathotypes in olive. Therefore, our study sought to document for the first time the natural co-occurrence of D and ND pathotypes in field-grown cotton sampled in NSW, Australia, which were also demonstrated again in our co-inoculated pot trials.
3. Discussion
Verticillium wilt of cotton was first reported in NSW in 1959 [
27]. Over 35 years of continuous disease survey since 1984 [
8], we for the first time documented the natural co-occurrence of both D and ND pathotypes of
V. dahliae in cotton in NSW, Australia, sampled from two survey seasons between 2017 and 2019. Additionally, we were able to demonstrate the co-infection in two independently repeated pot trials using a root dip method. Therefore, we propose that co-infections of multiple
V. dahliae isolates in a cotton plant are probably not a rare event under natural field conditions. Previously, Mercado-Blanco et al. [
19] reported the co-infection of D and ND pathotypes occurred naturally in olive trees. Schnathorst and Mathre [
26] artificially co-inoculated cotton with the virulent D isolate T-1 and mildly virulent ND isolate SS-4; however, there was no evidence of co-infection recorded in their study.
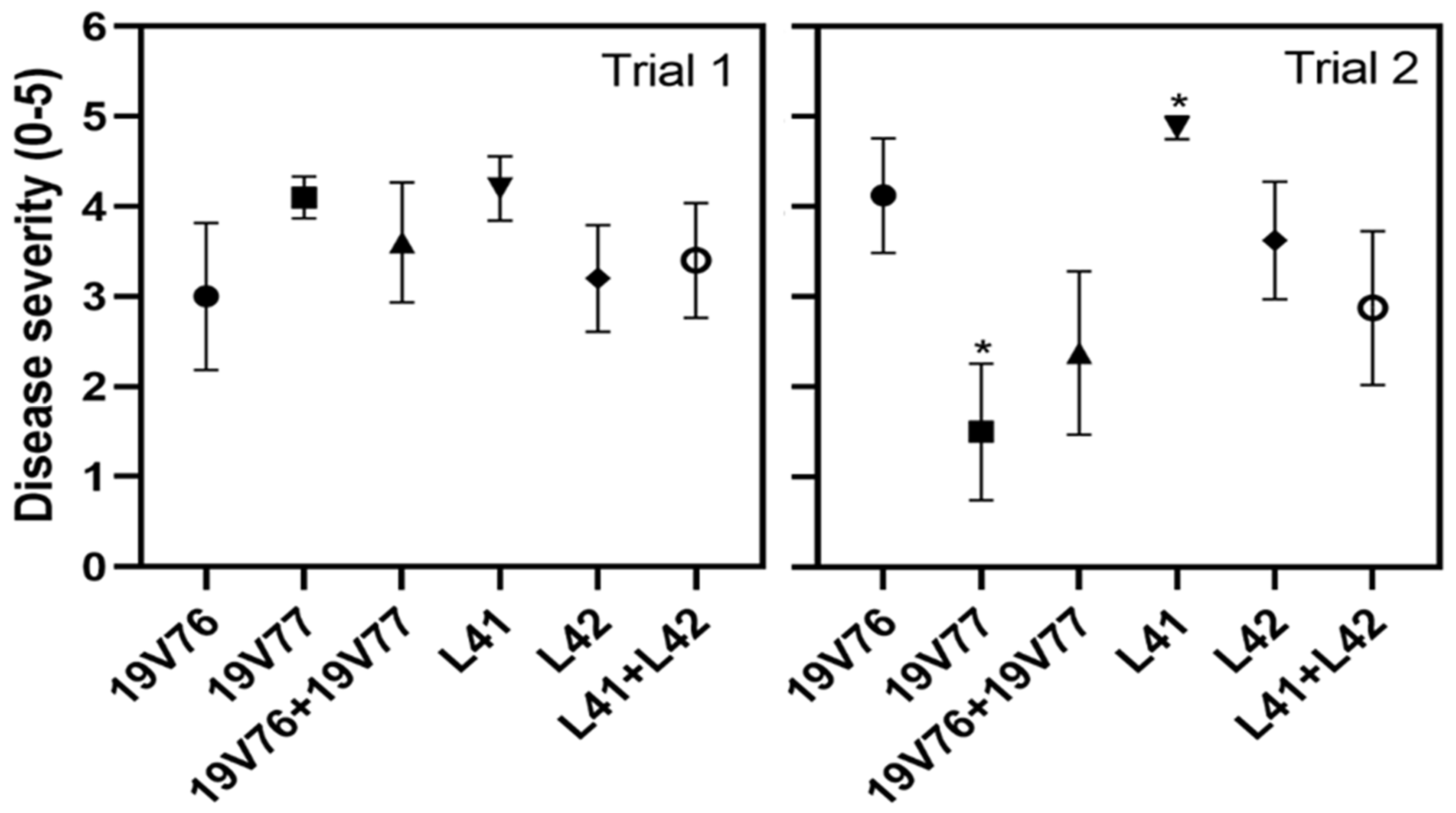
Our pot trials indicated that the D and ND isolates recovered from the same stem were relatively comparable in virulence on cotton, and the VW disease expressions were not significantly different between the single- and co-inoculated plants. Irrespective of treatments, initial VW symptoms appeared first on cotyledons between 7 and 9 days after inoculation and dead plants were observed across the treatments four weeks after inoculation. Similarly, Wheeler and Johnson [
22] reported VW disease in potato plants co-inoculated with three isolates from potato, mint and tomato was highly comparable to plants who received a single inoculation with the potato isolate. Interestingly, Schnathorst and Mathre [
26] found cross-protection on cotton when co-inoculated with virulent D and mildly virulent ND isolates. Cross-protection was more often associated with co-inoculations of avirulent and virulent isolates. Shittu et al. [
25] reported the VW disease scores were reduced by half on tomato plants co-inoculated with the highly virulent Vd1 and endophyte Dvd-E6 isolates compared with those recorded from single-inoculated tomato with Vd1. Sunflower challenged prior with the avirulent isolate P6 was protected from sequential inoculation with a virulent isolate of VM,
V. dahliae; however, simultaneous inoculation of the two isolates did not result in different disease expressions [
24].
We proposed that the D and ND isolates were able to colonise cotton stems equally due to the even number of isolates recovered from the same field-sampled stems. However, in our pot trials, the recovery frequency of D and ND isolates from the confirmed co-infected stems were predominated by one to the other. Mercado-Blanco [
19] directly detected both D and ND isolates from co-inoculated olive roots at 0, 7 and 21 days after inoculation; however, DNA directly obtained from stems of the same plants were only positive with the D marker. Similarly, Shittu et al. [
25] suggested that tomato plants colonised by the endophyte isolate Dvd-E6 ameliorated the effectiveness of colonisation of the pathogenic isolate Vd1. We postulated that vegetative growth competition may play a minor role in co-infection and co-colonisation of the tested
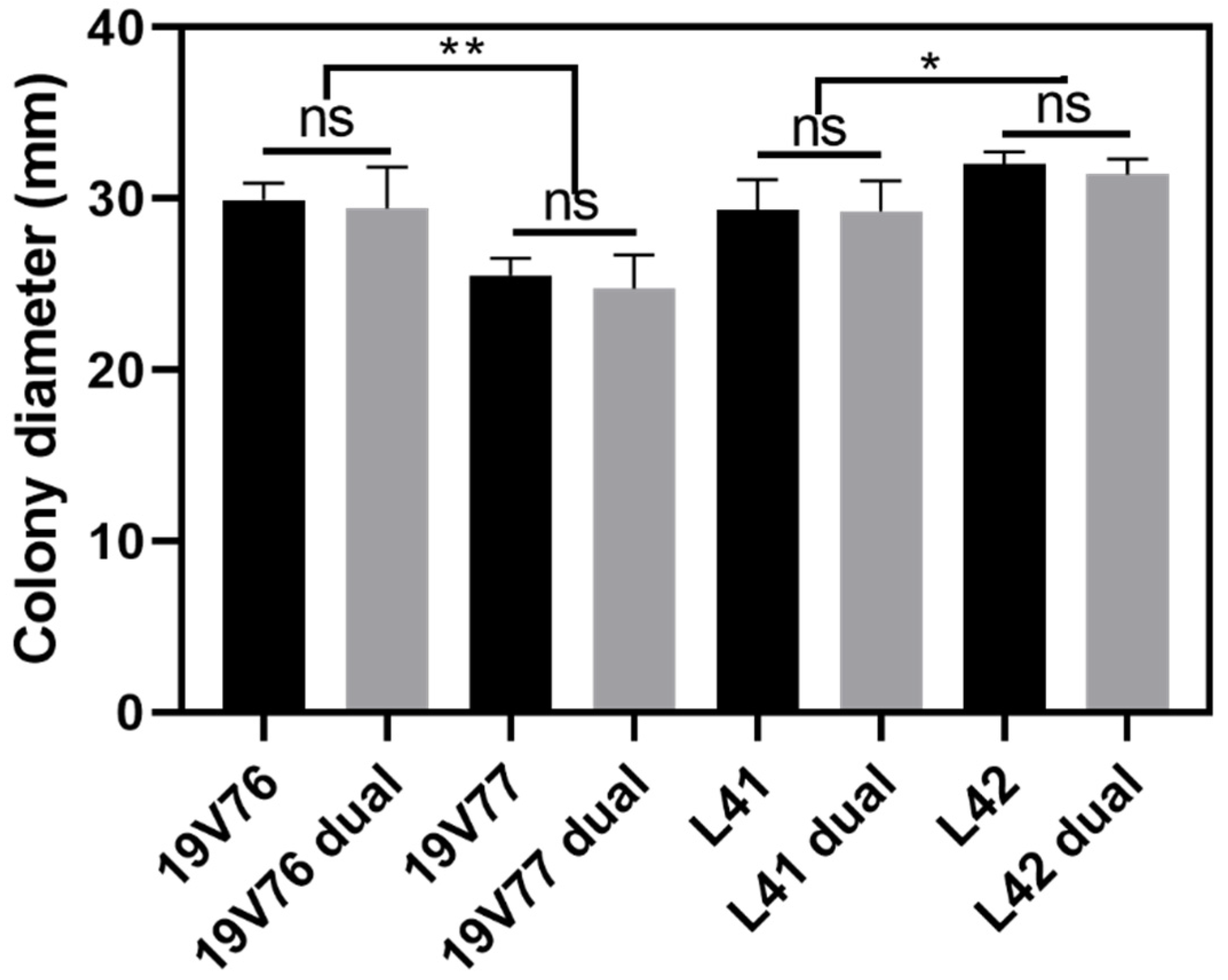
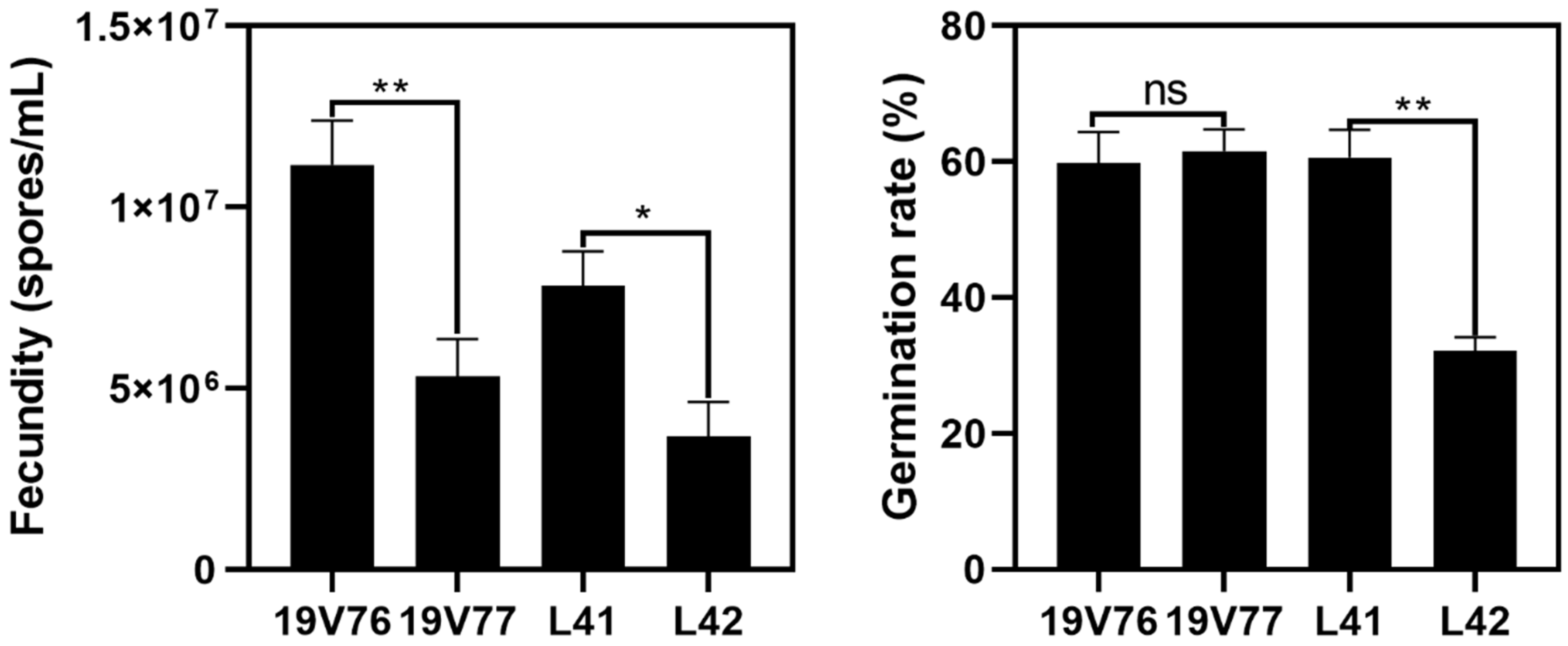
V. dahliae isolates in co-inoculated plants since there were no negative effects of one to the other detected in the dual culture assays. Fecundity and the spore germination rate of the tested isolates could partly play a role in driving this predominant colonisation. For example, the isolate 19V76 produced significantly more spores than 19V77; subsequently, in the two pot trials, the isolate 19V76 was recovered more than the isolate 19V77 in co-inoculated plants. However, we did not see a similar pattern in plants co-inoculated with the L41 and L42 isolates, though L41 produced a higher number of spores which had double the germination rate compared with L42. It is not possible to offer any conclusive recommendations from the current data set since host plant responses also play an important role in a successful colonisation of the pathogen. It will be worth using green fluorescent protein (GFP)-tagged isolates to better understand the interactions between
V. dahliae isolates as well as with the host in planta. GFP-tagged
V. dahliae was studied to understand its capacity to colonise cotton cultivars with different degrees of susceptibility [
28].
The occurrence of both D and ND pathotypes within a single cotton plant under field conditions has also raised concerns regarding the development of disease management strategies. First, commercial Australian cotton germplasms exhibited varietal responses to D and ND pathotypes differently, and to date, there has not been a highly resistant cultivar against both pathotypes, especially to the D pathotype (C. R. Trapero per. comm.). Therefore, there will be limited cultivars available for
V. dahliae-infested fields where the co-occurrence of both D and ND pathotypes in cotton plants were detected. However, before this raises an alarm, a thorough assessment of the damage that co-infection may cause should be pursued. The co-infection incidence under natural field conditions was detected at a low level in the previous two seasons. We continue to carry out disease surveillance, which has been ongoing for over 30 years in NSW, to enable the monitoring of the occurrence and distribution of the co-infection of both D and ND pathotypes on cotton. Second, genetic combination was also questioned. Wheeler and Johnson [
22] recently reported the putative anastomosis in planta when mustard plants were co-inoculated with three isolates: potato 653, mint 111 and tomato 461. Therefore, understanding the diversity of the isolate collection recovered from co-infected cotton in this study will warrant future research.
4. Materials and Methods
4.1. Sampling and Isolating
During the 2017/18 and 2018/19 survey seasons, cotton stems from VW-suspected plants were sampled for further confirmation through isolation and identification of the actual causal agent. Where possible, at least three VW-suspected stem cuts, approximately 10–15 cm long, were sampled from each of the surveyed fields. These stem cuts were double-bagged inside a paper bag and another outside zip-lock plastic bag. The stem cuts were stored in an esky (a portable cooler) away from direct sunlight during the survey trips and immediately transferred to a 4 °C fridge/cold room after each trip until further processed.
Isolation of the putative pathogen was initiated by excising each of the stem cuts into smaller sections, 1–2 cm long, and peeling off the outer bark. Under aseptic conditions, each of the peeled sections was sprayed and left for 30 s with 70% ethanol for surface decontamination. The sprayed section was then plotted dry with paper towel and split in half using a sterile scalpel. Inner vascular discoloured tissues (wood chips) were thinly sliced with the scalpel and embedded into sPDA. sPDA was made of potato dextrose agar (PDA Difco) amended with 100 ppm streptomycin sulfate (Sigma Aldrich) and contained in Petri plates. The plates with embedded vascular tissues were sealed with parafilm and incubated at 25 °C in the dark for 3–5 days. Putative fungal colonies emerging from vascular tissues were individually sub-cultured onto new sPDA plates and single spore cultures were established. Pure cultures were then transferred onto half strength sPDA and incubated at 25 °C in darkness for at least a week before small plugs (0.5 cm2) were excised from the colony margins, submerged in sterile water and stored at room temperature for subsequent experimentation.
4.2. Pathotyping by Duplex PCR
4.2.1. DNA Extraction
Genomic DNA was obtained using a Wizard® Genomic DNA Purification Kit (Promega, Sydney Australia) following the manufacture’s protocol. However, slight modifications were deployed to suit our laboratory conditions. A small amount of mycelia (10–100 mg) was scraped off culture plates and transferred into a 1.5 mL tight-lock Eppendorf tube. Then, 50 µL Nuclei Lysis Solution was added along with two steel beads (3.3 mm dia.) to each tube and shaken to macerate the mycelia on a tissue lyser (Retsch® MM300) for 1 min at a frequency of 28 times per second. Another 450 µL Nuclei Lysis Solution was then added to each of the tubes, vortexed to homogenicity and followed by incubation in a water bath at 65 °C for 30 min. After incubation, 3 µL RNase A Solution was added to each of the tubes and followed by another incubation at 37 °C for 15 min. After cooling down at room temperature for 5 min, 200 µL Protein Precipitation Solution was added and vortexed vigorously to homogenicity and followed by a centrifugation at 13,000 rpm for 5 min. The supernatants were carefully transferred to new 1.5 mL Eppendorf tubes containing 600 µL room temperature isopropanol. The tubes were gently inverted and centrifuged at 13,000 rpm for 1 min. The supernatants were carefully decanted and the visible DNA pellets were washed twice with 70% room temperature ethanol. The DNA pellets were then air-dried under a fume hood for 30–45 min, rehydrated with 50–200 µL DNA Rehydration Solution depending on the size of the pellets and followed with an incubation at 65 °C for 1 h. The DNA solutions were then stored at −20 °C until use.
4.2.2. Duplex PCR Amplification
A duplex PCR assay developed by Mercado-Blanco et al. [
19] was deployed to simultaneously characterise the two defoliating (D) and non-defoliating (ND) pathotypes. All PCR amplifications were carried out using GoTaq
® G2 Green Master Mix (Promega). Each PCR mix contained: 10 µL of Green Master Mix, 8 µL of DNase free water, 1 µL of 10 mM primer mix and 1 µL of DNA template. DNase-free water was included as a negative (no-template) control. The primer mix included 3 portions of DB19 (5′-CGGTGACATAATACTGAGAG-3′), 2 portions of DB22 (5′-GACGATGCGGATTGAACGAA3′) and 1 portion of espdef01 (5′-TGAGACTCGGCTGCCACAC-3′). PCR cycling conditions were slightly modified from Mercado-Blanco et al. [
19] as follows: initial denaturation for 5 min at 94 °C followed by 35 cycles of 30 s at 94 °C, 30 s at 52 °C and 90 s at 72 °C, with a final elongation step of 7 min at 72 °C. The PCR products were run at 100 V in a GelRed (GeneTargetSolutions) pre-stained 1.5% agarose gel for 45 min and visualised under UV light using a UVIDOC HD6 (UVITEC Cambridge). D pathotype isolates were predicted to contain two visible PCR amplicons at sizes of 539 and 334 bp, whereas ND pathotype isolates contained a single amplicon at a size of 523 bp [
19].
4.3. Pathogenicity
Root-dip assays were conducted twice in glasshouse conditions to assess the virulence of D and ND isolates solely and in combination. Two isolates from each of the pathotypes were selected for the pathogenicity assays (
Table 3). Both D and ND isolates recovered from the corresponding year were isolated from a single stem.
Cotton black seeds (cv. Sicot 75RRF, a VW susceptible cultivar) were sown individually in a 100-cell plastic seedling tray containing Searles® potting mix and grown in a glasshouse at 10–25 °C for 15 days prior to inoculation. Upon inoculation, seedlings were gently removed from the seedling trays and washed free of the potting mix. The washed seedlings were root-dipped in conidial suspensions of the four tested isolates at the concentration of 104 conidia/mL for five minutes and then transferred into 140 mL pots containing Searles® potting mix. There were two additional co-inoculum mixtures of L41 and L42, and 19V76 and 19V77. Conidial suspensions were mixed and adjusted to 104 conidia each/mL. There were two seedlings per pot and five and four replicate pots for each of treatment, respectively, in trial 1 and 2. Control seedlings were treated in the same manner; however, the spore suspension was replaced with sterile water. Inoculated seedlings were maintained in the same glasshouse conditions and monitored for disease occurrence and severity. Disease ratings were as follows: 0 = no symptoms, 1 = chlorosis and wilting of cotyledons, 2 = chlorosis and wilting of first true leaf, 3 = symptoms on lower 50% of the foliage, 4 = symptoms on 51–100% of the foliage and 5 = dead plant.
Upon termination of the pathogenicity assays, all plants were subjected to re-isolation of the inoculated pathogens. The isolation of the inoculated V. dahliae from the collar sections was as described previously. All recovered isolates were also subjected to the duplex PCR again for determination of the pathotypes.
4.4. Growth Competition Assays
Relative growth of the four tested isolates was assessed singly at 25 °C. Briefly, actively growing V. dahliae cultures on sPDA were excised into 0.5 cm2 plugs and transferred into the centre of new sPDA plates. Additionally, a dual cultures technique was undertaken to assess the growth competition of the V. dahliae isolates under the influence of one to another. As previously, active cultures of L41 and L42, and 19V76 and 19V77 were excised, transferred in pairs together and placed 5 cm apart onto new sPDA plates. All newly transferred plates were sealed with parafilm and incubated in darkness at 25 °C for seven days before the colony dia. was recorded in perpendicular directions. There were three replicate plates per each isolate/pair and the whole assay was repeated once.
4.5. Fecundity and Germination Assessments
Ten-day-old cultures growing at 25 °C in darkness were subjected to spore collection for fecundity and germination assessments. There were three plates per isolate for the fecundity examination. Each plate was flooded with 10 mL of sterile water and gently interrupted with a disposable L-shaped spreader. The spore concentration per mL of the collected suspension was determined using a haemocytometer. The spore suspension was then adjusted to 104 per mL for the germination assessment. Three 10 µL aliquots from each of the spore suspensions were individually spread onto clean sPDA plates (n = 9 per isolate). The plates were then incubated at 25 °C in darkness for two days, after which the colonies of V. dahliae were recorded. The experiment was repeated once.
Data collected from the pathogenicity, growth rate, fecundity and germination assays were subjected to an analysis of variance (ANOVA) and the separations of means were determined by Tukey’s least significant difference (LSD) test (p ≤ 0.05). The ANOVA and graphing were performed with Graphpad Prism 8.2.0.






{kind=link}
{kind=link}
{kind=link}