In Vitro Propagation of an Endangered Helianthus verticillatus by Axillary Bud Proliferation

 , , and
, , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Induction of Axillary Bud Development
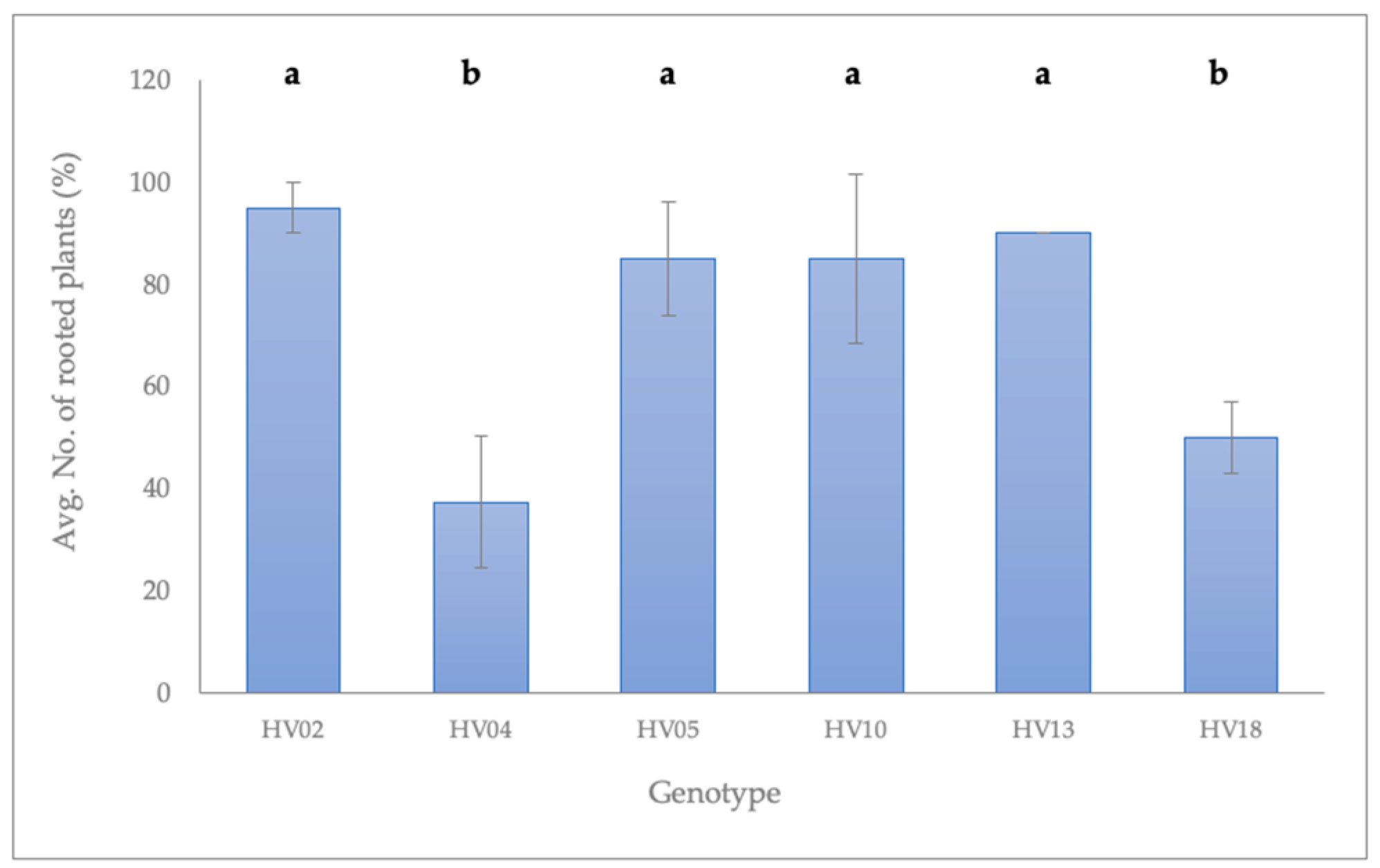
2.2. Rooting of Shoots Induced from the Axillary Buds
2.3. Acclimatization
2.4. Assessment of Genetic Fidelity of Micropropagated Plants
3. Materials and Methods
3.1. Plant Materials and Explant Preparation
3.2. Media and Culture Conditions
3.3. Induction of Axillary Shoots from the Nodal Stem Segments
3.4. Axillary Shoots Elongation and Rooting
3.5. Acclimatization
3.6. Assessment of Genetic Fidelity of Micropropagated Plants
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Matthews, J.F.; Allison, J.R.; Ware Sr, R.T.; Nordman, C. Helianthus verticillatus Small (Asteraceae) rediscovered and redescribed. Castanea 2002, 67, 13–24. [Google Scholar]
- Mandel, J.R. Clonal diversity, spatial dynamics, and small genetic population size in the rare sunflower, Helianthus verticillatus. Conserv. Genet. 2010, 11, 2055–2059. [Google Scholar] [CrossRef]
- Mississippi Plant Conservation Alliance. Available online: https://mississippiplantconservation.org/species-2/ (accessed on 23 February 2020).
- US Fish and Wildlife Service. Endangered and Threatened Wildlife and Plants; Designation of Critical Habitat for Physaria globosa (Short’s bladderpod), Helianthus verticillatus (whorled sunflower), and Leavenworthia crassa (Fleshy-Fruit Gladecress); Final Rule; US Fish and Wildlife Service: Washington, DC, USA, 2014; Volume 79, pp. 50990–51039.
- Trigiano, R.N.; Dominguez McLaughlin, H.; Lawton, C.; Gwinn, K.; Boggess, S.L. First report of a Cercospora species causing leaf spot on the whorled sunflower, Helianthus verticillatus, in the United States. Plant Dis. 2020. [Google Scholar] [CrossRef]
- Seiler, G.; Gulya, T.; Kong, G. Oil concentration and fatty acid profile of wild Helianthus species from the southeastern United States. Ind. Crops Prod. 2010, 31, 527–533. [Google Scholar] [CrossRef]
- Seiler, G.J.; Gulya, T.J. Evaluation of Achenes of Helianthus verticillatus, an Endemic Sunflower of the Southeast US for Oil Concentration and Fatty Acid Composition. Available online: https://www.sunflowernsa.com/uploads/research/572/seiler_achenes_11.pdf (accessed on 23 February 2020).
- Ngezahayo, F.; Liu, B. Axillary bud proliferation approach for plant biodiversity conservation and restoration. Int. J. Biodivers. 2014, 2014. [Google Scholar] [CrossRef] [Green Version]
- Fay, M.F. Conservation of rare and endangered plants using in vitro methods. In Vitro Cell. Dev. Biol.-Plant 1992, 28, 1–4. [Google Scholar] [CrossRef]
- García-Rubio, O.; Malda-Barrera, G. Micropropagation and reintroduction of the endemic Mammillaria mathildae (Cactaceae) to its natural habitat. HortScience 2010, 45, 934–938. [Google Scholar] [CrossRef] [Green Version]
- Misic, D.M.; Ghalawenji, N.A.; Grubisic, D.V.; Konjevic, R.M. Micropropagation and reintroduction of Nepeta rtanjensis, an endemic and critically endangered perennial of Serbia. Phyton-Ann. Rei Bot. 2005, 45, 9–20. Available online: https://www.zobodat.at/pdf/PHY_45_1_0009-0020.pdf (accessed on 23 February 2020).
- Hristova, L.; Damyanova, E.; Doichinova, Z.; Kapchina-Toteva, V. Effect of 6-benzylaminopurine on micropropagation of Artemisia chamaemelifolia Vill.(Asteraceae). Bulg. J. Agric. Sci. 2013, 19, 57–60. Available online: http://agrojournal.org/ARH.htm (accessed on 23 February 2020).
- Malda, G.; Suzán, H.; Backhaus, R. In vitro culture as a potential method for the conservation of endangered plants possessing crassulacean acid metabolism. Sci. Hortic. 1999, 81, 71–87. [Google Scholar] [CrossRef]
- Corral, P.; Mallón, R.; Rodríguez-Oubiña, J.; González, M.L. Multiple shoot induction and plant regeneration of the endangered species Crepis novoana. Plant Cell Tissue Organ Cult. 2011, 105, 211–217. [Google Scholar] [CrossRef]
- Cui, Y.; Deng, Y.; Zheng, K.; Hu, X.; Zhu, M.; Deng, X.; Xi, R. An efficient micropropagation protocol for an endangered ornamental tree species (Magnolia sirindhorniae Noot. & Chalermglin) and assessment of genetic uniformity through DNA markers. Sci. Rep. 2019, 9, 9634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ibanez, M.; Amo-Marco, J. Promotion by phloroglucinol of micropropagation of Minuartia valentina, an endangered and endemic Spanish plant. Plant Growth Regul. 1998, 26, 49–56. [Google Scholar] [CrossRef]
- He, H.; Deng, Z.; Jin, H. An efficient micropropagation system for Morinda officinalis How.(Rubiaceae), an endangered medicinal plant. J. Agric. Sci. Technol. 2015, 17.6, 1609–1618. Available online: http://hdl.handle.net/123456789/3821 (accessed on 23 February 2020).
- Jain, R.; Sinha, A.; Kachhwaha, S.; Kothari, S. Micropropagation of Withania coagulans (Stocks) Dunal: A critically endangered medicinal herb. J. Plant Biochem. Biotechnol. 2009, 18, 249–252. [Google Scholar] [CrossRef]
- Faisal, M.; Ahmad, N.; Anis, M. An efficient micropropagation system for Tylophora indica: An endangered, medicinally important plant. Plant Biotechnol. Rep. 2007, 1, 155–161. [Google Scholar] [CrossRef]
- Jordan, A.M.; Calvo, M.C.; Segura, J. Micropropagation of adult Lavandula dentata plants. J. Hortic. Sci. Biotechnol. 1998, 73, 93–96. [Google Scholar] [CrossRef]
- Rani, V.; Raina, S. Genetic fidelity of organized meristem-derived micropropagated plants: A critical reappraisal. In Vitro Cell. Dev. Biol.—Plant 2000, 36, 319–330. [Google Scholar] [CrossRef]
- George, E.F.; Hall, M.A.; De Klerk, G.-J. Micropropagation: Uses and Methods. In Plant Propagation by Tissue Cultura; George, E.F., Hall, M.A., De Klerk, G.-J., Eds.; The Background; Springer: Dordrecht, The Netherlands, 2008; pp. 29–64. [Google Scholar] [CrossRef]
- Hu, C.; Wang, P. Meristem, shoot tip and bud cultures. In Handbook of Plant Cell Culture; Evans, D., Sharp, W., Ammirato, P., Yamada, Y., Eds.; MacMillan: New York, NY, USA, 1983; Volume 1, pp. 177–227. [Google Scholar]
- Krishna, H.; Alizadeh, M.; Singh, D.; Singh, U.; Chauhan, N.; Eftekhari, M.; Sadh, R.K. Somaclonal variations and their applications in horticultural crops improvement. 3 Biotech 2016, 6, 54. [Google Scholar] [CrossRef] [Green Version]
- George, E.F. Plant Propagation by Tissue Culture. Part 1: The Technology; Exegetics Limited: Westbury, UK, 1993.
- Bairu, M.W.; Aremu, A.O.; Van Staden, J. Somaclonal variation in plants: Causes and detection methods. Plant Growth Regul. 2011, 63, 147–173. [Google Scholar] [CrossRef]
- Bandupriya, H.-D.; Iroshini, W.-A.; Perera, S.-N.; Vidhanaarachchi, V.-M.; Fernando, S.C.; Santha, E.S.; Gunathilake, T.R. Genetic fidelity testing using SSR marker assay confirms trueness to type of micropropagated coconut (Cocos nucifera L.) plantlets derived from unfertilized ovaries. Open Plant Sci. J. 2017, 10. [Google Scholar] [CrossRef] [Green Version]
- Chandrasekera, B.; Fluess, H.; Zhao, Y.; Trigiano, R.N.; Winkelmann, T. In vitro plant regeneration from ovules of Taraxacum officinale and Taraxacum koksaghyz. Afr. J. Biotechnol. 2017, 16, 1764–1775. [Google Scholar] [CrossRef] [Green Version]
- Pandey, R.N.; Singh, S.P.; Rastogi, J.; Sharma, M.L.; Singh, R.K. Early assessment of genetic fidelity in sugarcane (Saccharum officinarum) plantlets regenerated through direct organogenesis with RAPD and SSR markers. Aust. J. Crop Sci. 2012, 6, 618. [Google Scholar]
- Rai, M.K.; Phulwaria, M.; Gupta, A.K.; Shekhawat, N.S.; Jaiswal, U. Genetic homogeneity of guava plants derived from somatic embryogenesis using SSR and ISSR markers. Plant Cell Tissue Organ Cult. 2012, 111, 259–264. [Google Scholar] [CrossRef]
- Espinasse, A.; Lay, C. Shoot regeneration of callus derived from globular to torpedo embryos from 59 sunflower genotypes. Crop Sci. 1989, 29, 201–205. [Google Scholar] [CrossRef]
- Hewezi, T.; Jardinaud, F.; Alibert, G.; Kallerhoff, J. A new approach for efficient regeneration of a recalcitrant genotype of sunflower (Helianthus annuus) by organogenesis induction on split embryonic axes. Plant Cell Tissue Organ Cult. 2003, 73, 81–86. [Google Scholar] [CrossRef]
- Laparra, H.; Stoeva, P.; Ivanov, P.; Hahne, G. Plant regeneration from different explants in Helianthus smithii Heiser. Plant Cell Rep. 1997, 16, 692–695. [Google Scholar] [CrossRef] [PubMed]
- Lupi, M.C.; Bennici, A.; Locci, F.; Gennai, D. Plantlet formation from callus and shoot-tip culture of Helianthus annuus (L.). Plant Cell Tissue Organ Cult. 1987, 11, 47–55. [Google Scholar] [CrossRef]
- Paterson, K.E. Shoot tip culture of Helianthus annuus—Flowering and development of adventitious and multiple shoots. Am. J. Bot. 1984, 71, 925–931. [Google Scholar] [CrossRef]
- Prado, E.; Berville, A. Induction of somatic embryo development by liquid culture in sunflower (Helianthus annuus L.). Plant Sci. 1990, 67, 73–82. [Google Scholar] [CrossRef]
- Pugliesi, C.; Megale, P.; Cecconi, F.; Baroncelli, S. Organogenesis and embryogenesis in Helianthus tuberosus and in the interspecific hybrid Helianthus annuus × Helianthus tuberosus. Plant Cell Tissue Organ Cult. 1993, 33, 187–193. [Google Scholar] [CrossRef]
- Punia, M.S.; Bohorova, N.E. Callus development and plant regeneration from different explants of six wild species of sunflower (Helianthus L.). Plant Sci. 1992, 87, 79–83. [Google Scholar] [CrossRef]
- Weber, S.; Horn, R.; Friedt, W. High regeneration potential in vitro of sunflower (Helianthus annuus L.) lines derived from interspecific hybridization. Euphytica 2000, 116, 271–280. [Google Scholar] [CrossRef]
- Witrzens, B.T.; Scowcroft, W.R.; Downes, R.W.; Larkin, P.J. Tissue culture and plant regeneration from sunflower (Helianthus annuus) and interspecific hybrids (H. tuberosus × H. annuus). Plant Cell Tissue Organ Cult. 1988, 13, 61–76. [Google Scholar] [CrossRef]
- Yordanov, Y.; Yordanova, E.; Atanassov, A. Plant regeneration from interspecific hybrid and backcross progeny of Helianthus eggertii × Helianthus annuus. Plant Cell Tissue Organ Cult. 2002, 71, 7–14. [Google Scholar] [CrossRef]
- Deglene, L.; Lesignes, P.; Alibert, G.; Sarrafi, A. Genetic control of organogenesis in cotyledons of sunflower (Helianthus annuus). Plant Cell Tissue Organ Cult. 1997, 48, 127–130. [Google Scholar] [CrossRef]
- Inoka, K.-I.; Dahanayake, N. Effect of plant growth regulators on micro-propagation of sunflower (Helianthus annuus L.). Int. J. Sci. Res. Publ. 2015, 5, 1–5. [Google Scholar]
- Debnath, S. Growth and development of lingonberry cultivars as affected by in vitro and ex vitro culture methods and source propagule. HortScience 2004, 39, 891A–891. [Google Scholar] [CrossRef]
- Gaspar, T.; Kevers, C.; Penel, C.; Greppin, H.; Reid, D.M.; Thorpe, T.A. Plant hormones and plant growth regulators in plant tissue culture. In Vitro Cell. Dev. Biol.—Plant 1996, 32, 272–289. [Google Scholar] [CrossRef]
- Fatima, N.; Ahmad, N.; Ahmad, I.; Anis, M. Interactive effects of growth regulators, carbon sources, pH on plant regeneration and assessment of genetic fidelity using single primer amplification reaction (SPARS) techniques in Withania somnifera L. Appl. Biochem. Biotechnol. 2015, 177, 118–136. [Google Scholar] [CrossRef]
- Parveen, S.; Shahzad, A. Factors affecting in vitro plant regeneration from cotyledonary node explant of Senna sophera (L.) Roxb.–A highly medicinal legume. Afr. J. Biotechnol. 2014, 13. [Google Scholar] [CrossRef]
- Parveen, S.; Shahzad, A.; Saema, S. In vitro plant regeneration system for Cassia siamea Lam., a leguminous tree of economic importance. Agrofor. Syst. 2010, 80, 109–116. [Google Scholar] [CrossRef]
- Husain, M.K.; Anis, M. Rapid in vitro propagation of Eclipta alba (L.) Hassk. through high frequency axillary shoot proliferation. Acta Physiol. Plant. 2006, 28, 325–330. [Google Scholar] [CrossRef]
- Saha, S.; Adhikari, S.; Dey, T.; Ghosh, P. RAPD and ISSR based evaluation of genetic stability of micropropagated plantlets of Morus alba L. variety S-1. Meta Gene 2016, 7, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, V.; Prakash, S.; Gupta, S. Effective protocol for in vitro shoot production through nodal explants of Simmondsia chinensis. Biol. Plant. 2002, 45, 449–453. [Google Scholar] [CrossRef]
- Sathyanarayana, N.; Vikas, P.; Rajesha, R. In vitro clonal propagation of Mucuna pruriens var. utilis and its evaluation of genetic stability through RAPD markers. Afr. J. Biotechnol. 2008, 7, 973–980. Available online: https://www.ajol.info/index.php/ajb/article/view/58587 (accessed on 23 February 2020).
- Sen, J.; Sharma, A. Micropropagation of Withania somnifera from germinating seeds and shoot tips. Plant Cell Tissue Organ Cult. 1991, 26, 71–73. [Google Scholar] [CrossRef]
- Shahzad, A.; Parveen, S.; Fatema, M. Development of a regeneration system via nodal segment culture in Veronica anagallis-aquatica L.–an amphibious medicinal plant. J. Plant Interact. 2011, 6, 61–68. [Google Scholar] [CrossRef]
- Sivanesan, I.; Jeong, B.R. Direct shoot regeneration from nodal explants of Sida cordifolia Linn. In Vitro Cell. Dev. Biol.—Plant 2007, 43, 436–441. [Google Scholar] [CrossRef]
- Steephen, M.; Nagarajan, S.; Ganesh, D. Phloroglucinol and silver nitrate enhances axillary shoot proliferation in nodal explants of Vitex negundo L.–an aromatic medicinal plant. Iran. J. Biotechnol. 2010, 8, 82–89. Available online: https://www.sid.ir/en/Journal/ViewPaper.aspx?ID=171746 (accessed on 23 February 2020).
- Thomas, T.D.; Yoichiro, H. In vitro propagation for the conservation of a rare medicinal plant Justicia gendarussa Burm. f. by nodal explants and shoot regeneration from callus. Acta Physiol. Plant. 2010, 32, 943–950. [Google Scholar] [CrossRef]
- Amer, A.; Omar, H. In vitro propagation of the multipurpose Egyptian medicinal plant Pimpinella anisum. Egypt. Pharm. J. 2019, 18, 254–262. [Google Scholar] [CrossRef]
- Fatima, N.; Anis, M. Role of growth regulators on in vitro regeneration and histological analysis in Indian ginseng (Withania somnifera L.) Dunal. Physiol. Mol. Biol. Plants 2012, 18, 59–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lema-Rumińska, J.; Kulus, D. Micropropagation of cacti—A review. Haseltonia 2014, 2014, 46–63. [Google Scholar] [CrossRef]
- Park, H.-J.; Lee, H.-R.; Pyee, J.; Cha, H.-C. Regeneration of grape (Vitis labruscana cv. Kyoho) by shoot-tip culture. J. Plant Biol. 2001, 44, 185–192. [Google Scholar] [CrossRef]
- Skoog, F.; Miller, C. Chemical regulation of growth and organ formation in plant tissues cultured in vitro. Symp. Soc. Exp. Biol. 1957, 11, 118–130. [Google Scholar]
- Gamburg, K.Z.; Vysotskaya, E.; Gamanets, L. Microtuber formation in micropropagated Jerusalem artichoke (Helianthus tuberosus). Plant Cell Tissue Organ Cult. 1998, 55, 115–118. [Google Scholar] [CrossRef]
- Esan, E. Micropropagation of cocoa (Theobroma cacao L.). In High-tech and Micropropagation II; Springer: Berlin/Heidelberg, Germany, 1992; pp. 96–122. [Google Scholar] [CrossRef]
- Banilas, G.; Korkas, E. Rapid micropropagation of grapevine cv. Agiorgitiko through lateral bud development. J. Sci. Technol. 2007, 2, 31–38. [Google Scholar] [CrossRef]
- Qiu, Y.; Guan, S.C.; Wen, C.; Li, P.; Gao, Z.; Chen, X. Auxin and cytokinin coordinate the dormancy and outgrowth of axillary bud in strawberry runner. BMC Plant Biol. 2019, 19, 528. [Google Scholar] [CrossRef]
- Schneider, A.; Godin, C.; Boudon, F.; Demotes-Mainard, S.; Sakr, S.; Bertheloot, J. Light regulation of axillary bud outgrowth along plant axes: An overview of the roles of sugars and hormones. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Shimizu-Sato, S.; Tanaka, M.; Mori, H. Auxin–cytokinin interactions in the control of shoot branching. Plant Mol. Biol. 2008, 69, 429. [Google Scholar] [CrossRef] [Green Version]
- Baxter, R.; Brown, S.; England, N.; Ludlow, C.; Taylor, S.; Womack, R. Production of clonal plantlets of tropical pine in tissue culture via axillary shoot activation. Can. J. For. Res. 1989, 19, 1338–1342. [Google Scholar] [CrossRef]
- Jana, S.; Sivanesan, I.; Jeong, B.R. Effect of cytokinins on in vitro multiplication of Sophora tonkinensis. Asian Pac. J. Trop. Biomed. 2013, 3, 549–553. [Google Scholar] [CrossRef] [Green Version]
- Podwyszynska, M. Somaclonal variation in micropropagated tulips based on phenotype observation. J. Fruit Ornam. Plant Res. 2005, 13, 109–122. [Google Scholar]
- Xing, W.; Bao, M.; Qin, H.; Ning, G. Micropropagation of Rosa rugosa through axillary shoot proliferation. Acta Biol. Crac. Ser. Bot. 2010, 52, 69–75. [Google Scholar] [CrossRef]
- Brar, M.; Al-Khayri, J.; Morelock, T.; Anderson, E. Genotypic response of cowpea Vigna unguiculata (L.) to in vitro regeneration from cotyledon explants. In Vitro Cell. Dev. Biol.—Plant 1999, 35, 8–12. [Google Scholar] [CrossRef]
- Ochatt, S.J.; Conreux, C.; Jacas, L. In vitro production of sweet peas (Lathyrus odoratus L.) via axillary shoots. In Protocols for In Vitro Propagation of Ornamental Plants; Springer: Berlin/Heidelberg, Germany, 2010; Volume 589, pp. 293–301. [Google Scholar] [CrossRef]
- Catapan, E.; Otuki, M.F.; Viana, A. Cultura in vitro de Phyllanthus stipulatus (Euphorbiaceae). Braz. J. Bot. 2001, 24, 25–34. [Google Scholar] [CrossRef] [Green Version]
- Dias, M.C.; Almeida, R.; Romano, A. Rapid clonal multiplication of Lavandula viridis L’Hér through in vitro axillary shoot proliferation. Plant Cell Tissue Organ Cult. 2002, 68, 99–102. [Google Scholar] [CrossRef]
- Khan, N.; Ahmed, M.; Hafiz, I.; Abbasi, N.; Ejaz, S.; Anjum, M. Optimizing the concentrations of plant growth regulators for in vitro shoot cultures, callus induction and shoot regeneration from calluses of grapes. OENO One 2015, 49, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Rangsayatorn, N. Micropropagation of Dendrobium draconis Rchb. f. from thin cross-section culture. Sci. Hortic. 2009, 122, 662–665. [Google Scholar] [CrossRef]
- De Klerk, G.-J. Rooting of microcuttings: Theory and practice. In Vitro Cell. Dev. Biol.—Plant 2002, 38, 415–422. [Google Scholar] [CrossRef]
- Nookaraju, A.; Agrawal, D.C. Genetic homogeneity of in vitro raised plants of grapevine cv. Crimson Seedless revealed by ISSR and microsatellite markers. South Afr. J. Bot. 2012, 78, 302–306. [Google Scholar] [CrossRef] [Green Version]
- Agnihotri, R.K.; Mishra, J.; Nandi, S.K. Improved in vitro shoot multiplication and rooting of Dendrocalamus hamiltonii Nees et Arn. Ex Munro: Production of genetically uniform plants and field evaluation. Acta Physiol. Plant. 2009, 31, 961–967. [Google Scholar] [CrossRef]
- Joshi, P.; Dhawan, V. Assessment of genetic fidelity of micropropagated Swertia chirayita plantlets by ISSR marker assay. Biol. Plant. 2007, 51, 22–26. [Google Scholar] [CrossRef]
- Phulwaria, M.; Rai, M.K.; Shekhawat, N.S. An improved micropropagation of Arnebia hispidissima (Lehm.) DC. and assessment of genetic fidelity of micropropagated plants using DNA-based molecular markers. Appl. Biochem. Biotechnol. 2013, 170, 1163–1173. [Google Scholar] [CrossRef] [PubMed]
- Rathore, M.S.; Chikara, J.; Mastan, S.G.; Rahman, H.; Anand, K.-V.; Shekhawat, N.S. Assessment of genetic stability and instability of tissue culture-propagated plantlets of Aloe vera L. by RAPD and ISSR markers. Appl. Biochem. Biotechnol. 2011, 165, 1356–1365. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.R.; Dalal, S.; Singh, R.; Dhawan, A.K.; Kalia, R.K. Evaluation of genetic fidelity of in vitro raised plants of Dendrocalamus asper (Schult. & Schult. F.) Backer ex K. Heyne using DNA-based markers. Acta Physiol. Plant. 2013, 35, 419–430. [Google Scholar] [CrossRef]
- Swain, D.; Lenka, S.; Hota, T.; Rout, G.R. Micro-propagation of Hypericum gaitii Haines, an endangered medicinal plants: Assessment of genetic fidelity. Nucleus 2016, 59, 7–13. [Google Scholar] [CrossRef]
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. Peer J. 2014, 2, e281. [Google Scholar] [CrossRef] [Green Version]
- Bruvo, R.; Michiels, N.K.; D’Souza, T.G.; Schulenburg, H. A simple method for the calculation of microsatellite genotype distances irrespective of ploidy level. Mol. Ecol. 2004, 13, 2101–2106. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Pashley, C.H.; Ellis, J.R.; McCauley, D.E.; Burke, J.M. EST databases as a source for molecular markers: Lessons from Helianthus. J. Hered. 2006, 97, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Edwards, T.; Trigiano, R.N.; Ownley, B.; Windham, A.; Wyman, C.; Wadl, P.; Hadziabdic, D. Genetic diversity and conservation status of Helianthus verticillatus, an endangered sunflower of the southern United States. Front. Genet. 2020, 11, 236. [Google Scholar] [CrossRef]
- Ellis, J.; Pashley, C.; Burke, J.; McCauley, D. High genetic diversity in a rare and endangered sunflower as compared to a common congener. Mol. Ecol. 2006, 15, 2345–2355. [Google Scholar] [CrossRef]
- Wang, X.; Rinehart, T.A.; Wadl, P.A.; Spiers, J.M.; Hadziabdic, D.; Windham, M.T.; Trigiano, R.N. A new electrophoresis technique to separate microsatellite alleles. Afr. J. Biotechnol. 2009, 8, 2432–2436. Available online: https://www.ajol.info/index.php/ajb/article/view/60628 (accessed on 23 February 2020).
- Amos, W.; Hoffman, J.I.; Frodsham, A.; Zhang, L.; Best, S.; Hill, A.-S. Automated binning of microsatellite alleles: Problems and solutions. Mol. Ecol. Notes 2007, 7, 10–14. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage: Thousand Oaks, CA, USA, 2019; Available online: https://socialsciences.mcmaster.ca/jfox/Books/Companion/ (accessed on 23 February 2020).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| The BAP Concentration (µM) | Plants | The Stem Length (cm) 1 | The Root Length (cm) 1 | The Number of Roots 1 | The Number of Leaves 2 |
|---|---|---|---|---|---|
| 0.00 | HV02 | 7.4 ± 1.6 a | 9.1 ± 2.9 ab | 4.6 ± 1.5 ab | 6.5 ± 1.1 |
| HV04 | 4.9 ± 0.4 def | 1.6 ± 0.4 e–j | 1.8 ± 0.8 b–e | 7.3 ± 1.5 | |
| HV05 | 6.9 ± 1.4 b–f | 9.6 ± 2.9 a–e | 5.3 ± 1.3 abc | 7.2 ± 1.0 | |
| HV10 | 6.4 ± 1.4 b–f | 8.4 ± 2.2 a–g | 2.9 ± 0.9 b–f | 7.8 ± 0.6 | |
| HV13 | 4.0 ± 1.2 b–f | 5.1 ± 1.8 hij | 2.4 ± 0.9 def | 8.9 ± 1.4 | |
| HV18 | 6.7 ± 1.1 b–f | 7.0 ± 2.0 j | 3.8 ± 1.3 f | 6.8 ± 1.0 | |
| Mean | 6.1 | 6.8 | 3.5 | 7.4 A | |
| 0.88 | HV02 | 5.0 ± 2.2. c–f | 6.0 ± 4.8 f–j | 2.9 ± 0.8 def | 8.9 ± 2.8 |
| HV04 | 4.6 ± 0.6 b–f | 1.4 ± 0.8 hij | 1.8 ± 0.8 def | 8.4 ± 0.8 | |
| HV05 | 3.2 ± 0.9 ab | 2.3 ± 2.1 a | 1.9 ± 0.9 a | 9.2 ± 1.7 | |
| HV10 | 5.9 ± 2.4 def | 6.9 ± 2.4 d–j | 2.0 ± 0.8 b–f | 10.3 ± 1.3 | |
| HV13 | 4.1 ± 1.3 ef | 5.6 ± 1.5 f–j | 2.1 ± 0.9 c–f | 8.9 ± 1.0 | |
| HV18 | 5.7 ± 1.2 f | 3.1 ± 1.9 f–j | 1.5 ± 1.0 c–f | 8.7 ± 1.1 | |
| Mean | 4.7 | 4.2 | 2.0 | 9.1 B | |
| 2.22 | HV02 | 4.6 ± 1.4 a–d | 6.5 ± 2.0 d–j | 3.3 ± 1.4 bcd | 7.4 ± 1.4 |
| HV04 | 4.4 ± 0.5 a–f | 6.2 ± 1.5 abc | 2.3 ± 0.6 c–f | 8.4 ± 0.7 | |
| HV05 | 3.6 ± 0.5 a | 3.1 ± 2.6 a–f | 2.4 ± 1.3 b–e | 9.4 ± 1.9 | |
| HV10 | 7.4 ± 2.4 a–f | 6.8 ± 3.4 a–d | 3.4 ± 1.1 c–f | 10.3 ± 1.8 | |
| HV13 | 4.0 ± 0.5 d–f | 5.0 ± 1.4 c–i | 1.3 ± 0.7 c–f | 8.0 ± 0.0 | |
| HV18 | 5.8 ± 2.1 b–f | 2.4 ± 2.3 c–j | 2.0 ± 1.2 c–f | 8.2 ± 1.5 | |
| Mean | 5.0 | 5.0 | 2.5 | 8.6 B | |
| 4.44 | HV02 | 4.1 ± 1.0 def | 2.6 ± 1.7 c–j | 3.0 ± 1.4 c–f | 8.5 ± 2.4 |
| HV04 | 4.7 ± 1.3 def | 1.3 ± 1.1 b–h | 2.0 ± 1.4 c–f | 8.0 ± 0.7 | |
| HV05 | 4.1 ± 1.1 abc | 3.5 ± 3.1 c–j | 3.1 ± 1.5 c–f | 8.4 ± 0.7 | |
| HV10 | 5.6 ± 1.4 a–e | 3.4 ± 3.9 i–j | 2.5 ± 0.8 e–f | 9.0 ± 2.2 | |
| HV13 | 4.7 ± 1.6 a–e | 4.9 ± 1.6 hij | 2.4 ± 1.1 def | 9.6 ± 1.8 | |
| HV18 | 5.9 ± 1.0 a–e | 1.1 ± 0.4 g–j | 4.3 ± 1.5 e–f | 8.1 ± 0.3 | |
| Mean | 4.8 | 2.8 | 2.9 | 8.6 B |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nowakowska, M.; Pavlović, Ž.; Nowicki, M.; Boggess, S.L.; Trigiano, R.N. In Vitro Propagation of an Endangered Helianthus verticillatus by Axillary Bud Proliferation. Plants 2020, 9, 712. https://doi.org/10.3390/plants9060712
Nowakowska M, Pavlović Ž, Nowicki M, Boggess SL, Trigiano RN. In Vitro Propagation of an Endangered Helianthus verticillatus by Axillary Bud Proliferation. Plants. 2020; 9(6):712. https://doi.org/10.3390/plants9060712
Chicago/Turabian StyleNowakowska, Marzena, Žaklina Pavlović, Marcin Nowicki, Sarah L. Boggess, and Robert N. Trigiano. 2020. "In Vitro Propagation of an Endangered Helianthus verticillatus by Axillary Bud Proliferation" Plants 9, no. 6: 712. https://doi.org/10.3390/plants9060712