Solanales Stem Biomechanical Properties Are Primarily Determined by Morphology Rather Than Internal Structural Anatomy and Cell Wall Composition


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
Sampling
2.2. Biomechanical Measurements
2.3. Anatomy
2.4. Image Analysis
2.5. Immunohistochemistry
2.6. Fractionation of Cell Wall Polymers into Broad Classes
2.6.1. Preparation of Alcohol-Insoluble Residue
2.6.2. Separation of Polysaccharides
2.7. Statistical Analysis
3. Results
3.1. Structural Differences between the Stem Types
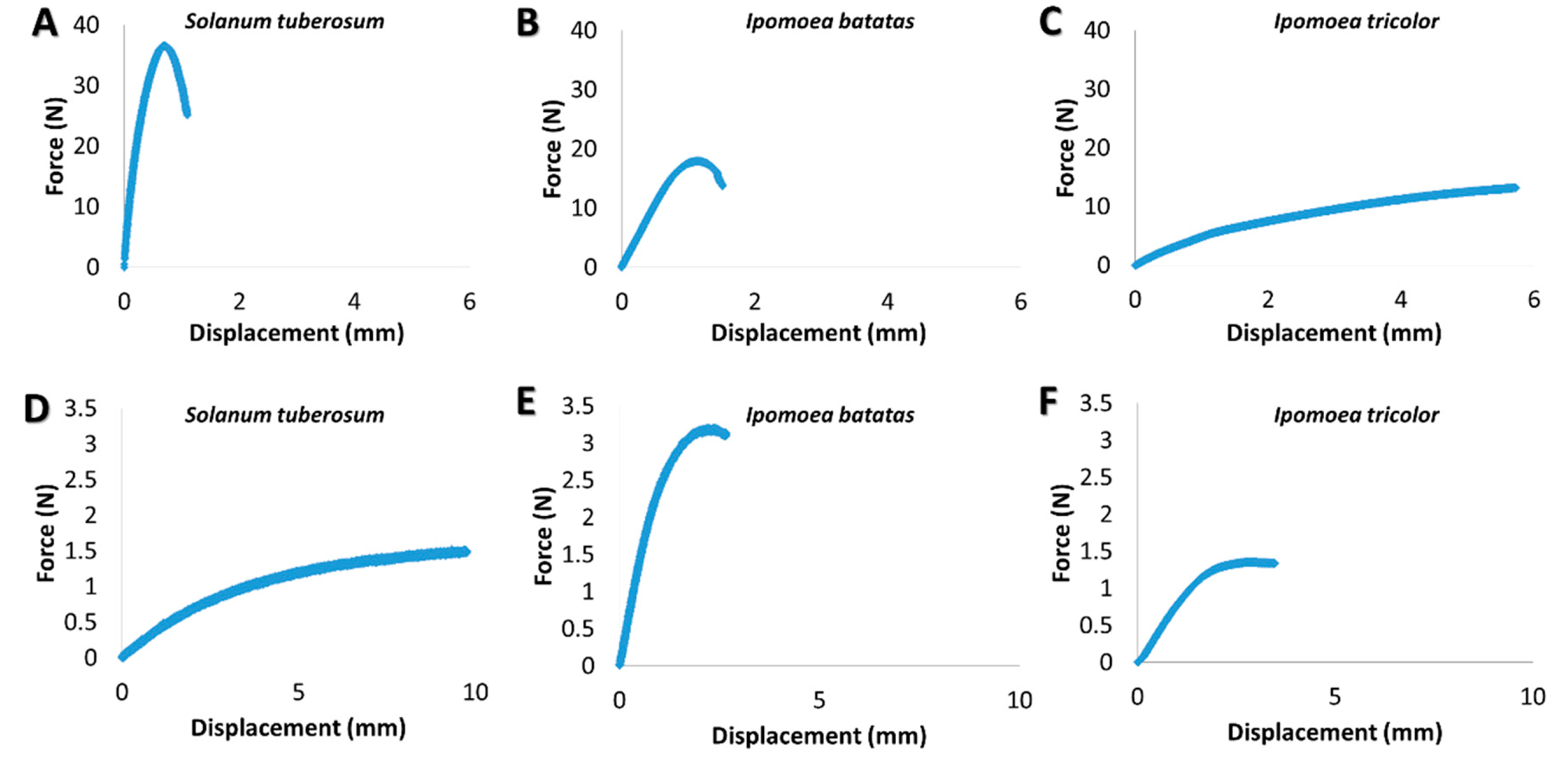
3.2. Biomechanical Stem Parameters
3.3. Cell wall Composition
3.4. Immunolocalisation of Specific Cell Wall Epitopes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Darwin, C. The Movements and Habits of Climbing Plants; John Murray: London, UK, 1882. [Google Scholar]
- Desalegn, W.; Beierkuhnlein, C. Plant species and growth form richness along altitudinal gradients in the southwest Ethiopian highlands. J. Veg. Sci. 2010, 21, 617–626. [Google Scholar] [CrossRef]
- Kooyman, R.M.; Zanne, A.E.; Gallagher, R.V.; Cornwell, W.K.; Rossetto, M.; O’Connor, P.; Parkes, E.A.; Catterall, C.F.; Laffan, S.W.; Lusk, C.H. Effects of Growth Form and Functional Traits on Response of Woody Plants to Clearing and Fragmentation of Subtropical Rainforest. Conserv. Biol. 2013, 27, 1468–1477. [Google Scholar] [CrossRef] [PubMed]
- Couvreur, T.L.P.; Kissling, W.D.; Condamine, F.L.; Svenning, J.-C.; Rowe, N.P.; Baker, W.J. Global diversification of a tropical plant growth form: Environmental correlates and historical contingencies in climbing palms. Front. Genet. 2015, 5, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, G.C. Lianas as structural parasites: The Bursera Simaruba Example. Ecology 1987, 68, 77–81. [Google Scholar] [CrossRef]
- Rowe, N.; Isnard, S.; Speck, T. Diversity of mechanical architectures in climbing plants: An evolutionary perspective. J. Plant Growth Regul. 2004, 23, 108–128. [Google Scholar] [CrossRef]
- Carlquist, S. Observations on functional wood histology of vines and lianas. Aliso 1985, 11, 139–157. [Google Scholar] [CrossRef] [Green Version]
- Ewers, F.W.; Fisher, J.B.; Chiu, S.-T. A survey of vessel dimensions in stems of tropical lianas and other growth forms. Oecologia 1990, 84, 544–552. [Google Scholar] [CrossRef]
- Carlquist, S.; Hanson, M. Wood and stem anatomy of Convolvulaceae. Aliso 1991, 13, 51–94. [Google Scholar] [CrossRef] [Green Version]
- Gartner, B.L. Stem hydraulic properties of vines vs. shrubs of western poison oak, Toxicodendron diversilobum. Oecologia 1991, 87, 180–189. [Google Scholar] [CrossRef]
- Shtein, I.; Hayat, Y.; Munitz, S.; Harcavi, E.; Akerman, M.; Drori, E.; Schwartz, A.; Netzer, Y. From structural constraints to hydraulic function in three Vitis rootstocks. Trees 2016, 31, 851–861. [Google Scholar] [CrossRef]
- Wyka, T.P.; Oleksyn, J.; Karolewski, P.; Schnitzer, S.A. Phenotypic correlates of the lianescent growth form: A review. Ann. Bot. 2013, 112, 1667–1681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speck, T. A biomechanical method to distinguish between self-supporting and non self-supporting fossil plants. Rev. Palaeobot. Palynol. 1994, 81, 65–82. [Google Scholar] [CrossRef]
- Rowe, N.P.; Speck, T. Biomechanical characteristics of the ontogeny and growth habit of the tropical liana Condylocarpon guianense (Apocynaceae). Int. J. Plant Sci. 1996, 157, 406–417. [Google Scholar] [CrossRef]
- Isnard, S.; Speck, T.; Rowe, N.P. Mechanical architecture and development in Clematis: Implications for canalised evolution of growth forms. New Phytol. 2003, 158, 543–559. [Google Scholar] [CrossRef] [Green Version]
- Brett, C.; Waldron, K. Physiology and Biochemistry of Plant Cell Walls. In Physiology and Biochemistry of Plant Cell Walls; Springer Science and Business Media LLC: Berlin, Germany, 1990. [Google Scholar]
- Shtein, I.; Bar-On, B.; Popper, Z. Plant and algal structure: From cell walls to biomechanical function. Physiol. Plant. 2018, 164, 56–66. [Google Scholar] [CrossRef]
- Zykwinska, A.W.; Ralet, M.-C.; Garnier, C.D.; Thibault, J.-F.J. Evidence for In Vitro Binding of Pectin Side Chains to Cellulose1. Plant Physiol. 2005, 139, 397–407. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.B.; Cosgrove, D.J. Changes in cell wall biomechanical properties in the xyloglucan-deficient xxt1/xxt2 mutant of Arabidopsis. Plant Physiol. 2012, 158, 465–475. [Google Scholar] [CrossRef] [Green Version]
- Nguema-Ona, E.; Moore, J.; Fagerström, A.D.; Fangel, J.U.; Willats, W.; Hugo, A.; Vivier, M. Overexpression of the grapevine PGIP1 in tobacco results in compositional changes in the leaf arabinoxyloglucan network in the absence of fungal infection. BMC Plant Biol. 2013, 13, 46. [Google Scholar] [CrossRef]
- Gu, J.; Catchmark, J.M. Roles of xyloglucan and pectin on the mechanical properties of bacterial cellulose composite films. Cellulose 2013, 21, 275–289. [Google Scholar] [CrossRef]
- Harris, P.J.; Kelderman, M.R.; Kendon, M.F.; McKenzie, R.J. Monosaccharide compositions of unlignified cell walls of monocotyledons in relation to the occurrence of wall-bound ferulic acid. Biochem. Syst. Ecol. 1997, 25, 167–179. [Google Scholar] [CrossRef]
- Smith, B.G.; Harris, P.J. The polysaccharide composition of Poales cell walls. Biochem. Syst. Ecol. 1999, 27, 33–53. [Google Scholar] [CrossRef]
- Carnachan, S.M.; Harris, P.J. Polysaccharide compositions of primary cell walls of the palms Phoenix canariensis and Rhopalostylis sapida. Plant Physiol. Biochem. 2000, 38, 699–708. [Google Scholar] [CrossRef]
- Popper, Z.; Fry, S.C. Primary cell wall composition of bryophytes and charophytes. Ann. Bot. 2003, 91, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popper, Z.; Fry, S.C. Primary cell wall composition of pteridophytes and spermatophytes. New Phytol. 2004, 164, 165–174. [Google Scholar] [CrossRef]
- Popper, Z.; Michel, G.; Herve, C.; Domozych, D.S.; Willats, W.; Tuohy, M.G.; Kloareg, B.; Stengel, D.B. Evolution and Diversity of Plant Cell Walls: From Algae to Flowering Plants. Annu. Rev. Plant Biol. 2011, 62, 567–590. [Google Scholar] [CrossRef] [Green Version]
- Bowling, A.J.; Vaughn, K.C. Gelatinous fibers are widespread in coiling tendrils and twining vines. Am. J. Bot. 2009, 96, 719–727. [Google Scholar] [CrossRef]
- Bowling, A.J.; Vaughn, K.C. Immunocytochemical characterization of tension wood: Gelatinous fibers contain more than just cellulose. Am. J. Bot. 2008, 95, 655–663. [Google Scholar] [CrossRef]
- Esau, K. Anatomy of Seed Plants. Soil Sci. 1960, 90, 149. [Google Scholar] [CrossRef]
- Hoffmann, B.; Chabbert, B.; Monties, B.; Speck, T. Mechanical, chemical and X-ray analysis of wood in the two tropical lianas Bauhinia guianensis and Condylocarpon guianense: Variations during ontogeny. Planta 2003, 217, 32–40. [Google Scholar] [CrossRef]
- Sanders, E. Switzerland plans futuristic train link. Phys. World 2000, 13, 9. [Google Scholar] [CrossRef]
- Eich, E. Solanaceae and Convolvulaceae: Secondary Metabolites; Springer: Berlin/Heidelberg, Germany, 2008. [Google Scholar]
- Available online: http://www.moag.gov.il/en/Ministrys%20Units/Training%20and%20Profession%20Service/Pages/default.aspx (accessed on 26 May 2020).
- Selle, H.K.; Bar-On, B.; Marom, G.; Wagner, H.D. Gelatin yarns inspired by tendons—Structural and mechanical perspectives. Mater. Sci. Eng. C 2015, 47, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Shtein, I.; Koyfman, A.; Eshel, A.; Bar-On, B. Autotomy in plants: Organ sacrifice in Oxalis leaves. J. R. Soc. Interface 2019, 16, 20180737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spatz, H.C.; O’Leary, E.J.; Vincent, J.F.V. Young’s moduli and shear moduli in cortical bone. Proc. R. Soc. B Biol. Sci. 1996, 263, 287–294. [Google Scholar] [CrossRef]
- Timoshenko, S. Strength of Materials—Part 1, 2nd ed.; D. Van Nostrand Company: New York, NY, USA, 1940. [Google Scholar]
- Roark, R.; Young, W. Formulas for Stress and Strain; McGraw-Hill: New York, NY, USA, 1975. [Google Scholar]
- Paiva, E.; Pinho, S.; Oliveira, D. Large plant samples: How to process for GMA embedding? In Methods in Molecular Biology; Chiarin-Garcia, H., Melo, R., Eds.; Springer/Humana Press: New York, NY, USA, 2011; pp. 1–45. [Google Scholar]
- Igersheim, A.; Cichocki, O. A simple method for microtome sectioning of prehistoric charcoal specimens, embedded in 2-hydroxyethyl methacrylate (HEMA). Rev. Palaeobot. Palynol. 1996, 92, 389–393. [Google Scholar] [CrossRef]
- O’Brien, T.P.; Feder, N.; McCully, M.E. Polychromatic staining of plant cell walls by toluidine blue O. Protoplasma 1964, 59, 368–373. [Google Scholar] [CrossRef]
- Rasband, W.S. US National Institutes of Health, Bethesda, Maryland, USA. 2011. Available online: http://imagej.nih.gov/ij/ (accessed on 1 May 2020).
- Herve, C.; Marcus, S.E.; Knox, J.P. Monoclonal antibodies, Carbohydrate-Binding Modules, and the detection of polysaccharides in plant cell walls. In Advanced Structural Safety Studies; Springer Science and Business Media LLC: Berlin, Germany, 2010; Volume 715, pp. 103–113. [Google Scholar]
- Leroux, O.; Sørensen, I.; Marcus, S.E.; Viane, R.L.; Willats, W.; Knox, J.P. Antibody-based screening of cell wall matrix glycans in ferns reveals taxon, tissue and cell-type specific distribution patterns. BMC Plant Biol. 2015, 15, 56. [Google Scholar] [CrossRef] [Green Version]
- Knox, J.P.; Linstead, P.J.; King, J.; Cooper, C.; Roberts, K. Pectin esterification is spatially regulated both within cell walls and between developing tissues of root apices. Planta 1990, 181, 512–521. [Google Scholar] [CrossRef]
- Steffan, W.; Kovác, P.; Albersheim, P.; Darvill, A.G.; Hahn, M.G. Characterization of a monoclonal antibody that recognizes an arabinosylated (1,6)-beta-d-galactan epitope in plant complex carbohydrates. Carbohydr Res. 1995, 275, 295–307. [Google Scholar] [CrossRef]
- Willats, W.G.; Marcus, S.E.; Knox, J.P. Generation of monoclonal antibody specific to (1-->5)-alpha-L-arabinan. Carbohydr Res. 1996, 308, 149–152. [Google Scholar] [CrossRef]
- Marcus, S.E.; Blake, A.W.; Benians, T.A.S.; Lee, K.J.D.; Poyser, C.; Donaldson, L.; Leroux, O.; Rogowski, A.; Petersen, H.L.; Boraston, A.; et al. Restricted access of proteins to mannan polysaccharides in intact plant cell walls. Plant J. 2010, 64, 191–203. [Google Scholar] [CrossRef]
- Pedersen, H.L.; Fangel, J.U.; McCleary, B.; Ruzanski, C.; Rydahl, M.G.; Ralet, M.-C.; Farkas, V.; von Schantz, L.; Marcus, S.E.; Andersen, M.C.F.; et al. Versatile high resolution oligosaccharide microarrays for plant glycobiology and cell wall research. J. Biol. Chem. 2012, 287, 39429–39438. [Google Scholar] [CrossRef] [Green Version]
- Pattathil, S.; Avci, U.; Baldwin, D.; Swennes, A.G.; McGill, J.A.; Popper, Z.; Bootten, T.; Albert, A.; Davis, R.H.; Chennareddy, C.; et al. A comprehensive toolkit of plant cell wall glycan-directed monoclonal antibodies. Plant Physiol. 2010, 153, 514–525. [Google Scholar] [PubMed] [Green Version]
- Pennell, R.I.; Janniche, L.; Kjellbom, P.; Scofield, G.N.; Peart, J.M.; Roberts, K. Developmental regulation of a plasma membrane arabinogalactan protein epitope in oilseed rape flowers. Plant Cell 1991, 3, 1317–1326. [Google Scholar] [PubMed]
- Smallwood, M.; Martin, H.; Knox, J.P. An epitope of rice threonine- and hydroxyproline-rich glycoprotein is common to cell wall and hydrophobic plasma-membrane glycoproteins. Planta 1995, 196, 510–522. [Google Scholar] [PubMed]
- O’Rourke, C.; Gregson, T.; Murray, L.; Sadler, I.H.; Fry, S.C. Sugar composition of the pectic polysaccharides of charophytes, the closest algal relatives of land-plants: Presence of 3-O-methyl-d-galactose residues. Ann. Bot. 2015, 116, 225–236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Usherwood, J.; Ball, D.; Ennos, A. Mechanical and anatomical adaptations in terrestrial and aquatic buttercups to their respective environments. J. Exp. Bot. 1997, 48, 1469–1475. [Google Scholar] [CrossRef] [Green Version]
- Spatz, H.-C.; Bruechert, F. Basic biomechanics of self-supporting plants: Wind loads and gravitational loads on a Norway spruce tree. For. Ecol. Manag. 2000, 135, 33–44. [Google Scholar] [CrossRef]
- Marcus, S.E.; Verhertbruggen, Y.; Herve, C.; Ordaz-Ortiz, J.J.; Farkas, V.; Pedersen, H.L.; Willats, W.; Knox, J.P. Pectic homogalacturonan masks abundant sets of xyloglucan epitopes in plant cell walls. BMC Plant Biol. 2008, 8, 60. [Google Scholar] [CrossRef] [Green Version]
- Ewers, F.W.; Fisher, J.B. Why vines have narrow stems: Histological trends in Bauhinia (Fabaceae). Oecologia 1991, 88, 233–237. [Google Scholar] [CrossRef]
- Silk, W.K.; Hubbard, M. Axial forces and normal distributed loads in twining stems of morning glory. J. Biomech. 1991, 24, 599–606. [Google Scholar] [CrossRef]
- Silk, W.K.; Holbrook, N.M. The importance of frictional interactions in maintaining the stability of the twining habit. Am. J. Bot. 2005, 92, 1820–1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isnard, S.; Silk, W.K. Moving with climbing plants from Charles Darwin’s time into the 21st century. Am. J. Bot. 2009, 96, 1205–1221. [Google Scholar] [CrossRef] [PubMed]
- Silva, W.B.; Vicente, M.; Robledo, J.M.; Reartes, D.; Ferrari, R.C.; Bianchetti, R.; Araujo, W.; Freschi, L.; Peres, L.E.; Zsögön, A. SELF-PRUNING Acts Synergistically with DIAGEOTROPICA to Guide Auxin Responses and Proper Growth Form. Plant Physiol. 2018, 176, 2904–2916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Popper, Z. Evolution and diversity of green plant cell walls. Curr. Opin. Plant Biol. 2008, 11, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, M.; Jia, Z.; Peña, M.J.; Cash, M.; Harper, A.; Blackburn, A.R.; Darvill, A.; York, W.S. Structural analysis of xyloglucans in the primary cell walls of plants in the subclass Asteridae. Carbohydr. Res. 2005, 340, 1826–1840. [Google Scholar] [CrossRef] [PubMed]
- Thimm, J.C.; Burritt, D.J.; Sims, I.M.; Newman, R.H.; Ducker, W.A.; Melton, L.D. Celery (Apium graveolens) parenchyma cell walls: Cell walls with minimal xyloglucan. Physiol. Plant. 2002, 116, 164–171. [Google Scholar] [CrossRef]
- Dick-Pérez, M.; Zhang, Y.; Hayes, J.; Salazar, A.; Zabotina, O.A.; Hong, M. Structure and interactions of plant cell-wall polysaccharides by Two- and Three-Dimensional Magic-Angle-Spinning Solid-State NMR. Biochemistry 2011, 50, 989–1000. [Google Scholar] [CrossRef]
- Cosgrove, D.J.; Jarvis, M.C. Comparative structure and biomechanics of plant primary and secondary cell walls. Front. Plant Sci. 2012, 3, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Spicer, R.A.; Niklas, K.J. Plant biomechanics: An engineering approach to plant form and function. J. Ecol. 1993, 81, 592. [Google Scholar] [CrossRef] [Green Version]
- Niklas, K.J. A mechanical perspective on foliage leaf form and function. New Phytol. 1999, 143, 19–31. [Google Scholar] [CrossRef]
- Mattheck, C. Design in nature. In Trees as Instructors; Rombach Verlag: Freiburg im Breisgau, Germany, 1997. [Google Scholar]
- Mattheck, G. Trees. In The Mechanical Design; Springer Science & Business Media: Berlin, Germany, 2012. [Google Scholar]
- Bar-On, B.; Sui, X.; Livanov, K.; Achrai, B.; Kalfon-Cohen, E.; Wiesel, E.; Wagner, H.D. Structural origins of morphing in plant tissues. Appl. Phys. Lett. 2014, 105, 33703. [Google Scholar] [CrossRef]
- Shtein, I.; Elbaum, R.; Bar-On, B. The hygroscopic opening of sesame fruits is induced by a functionally graded pericarp architecture. Front. Plant Sci. 2016, 7, 584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shtein, I.; Shelef, Y.; Marom, Z.; Zelinger, E.; Schwartz, A.; Popper, Z.; Bar-On, B.; Harpaz-Saad, S. Stomatal cell wall composition: Distinctive structural patterns associated with different phylogenetic groups. Ann. Bot. 2017, 119, 1021–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marom, Z.; Shtein, I.; Bar-On, B. Stomatal opening: The role of cell-wall mechanical anisotropy and its analytical relations to the bio-composite characteristics. Front. Plant Sci. 2017, 8, 2061. [Google Scholar] [CrossRef] [Green Version]
- Speck, T.; Burgert, I. Plant stems: Functional design and mechanics. Annu. Rev. Mater. Res. 2011, 41, 169–193. [Google Scholar] [CrossRef]
- Wilson, B.F. Shrub stems: Form and function. In Plant Stems; Gartner, B.L., Ed.; Elsevier Inc.: Amsterdam, The Netherlands, 1995; pp. 91–102. [Google Scholar]
- Mattheck, C.; Bethge, K. The Structural Optimization of Trees. Naturwissenschaften 1998, 85, 1–10. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Wall Components | mAb | Secondary Antibody | Supplier | Epitope Specificity |
|---|---|---|---|---|
| Pectins | JIM5 | rat | Plantprobes | Predominantly non-methyl-esterified homogalacturonans [46] |
| JIM7 | rat | Plantprobes | Methyl-esterified homogalacturonans [46] | |
| CCRC-M7 | mouse | Carbosource | Rhamnogalacturonan I (specifically -trimer of beta-(1,6)-Gal) [47] | |
| LM6 | rat | Plantprobes | (1→5)-β-Arabinan [48] | |
| Hemicelluloses | LM21 | rat | Plantprobes | Mannans, glucomannans, galactomannans (β-(1→4)-manno-oligosaccharides) [49] |
| LM25 | rat | Plantprobes | Xyloglucan (XLLG, XXLG and XXXG oligosaccharides of xyloglucan) [50] | |
| CCRC-M138 | mouse | Carbosource | Xylan-6 [51] | |
| AGPs | JIM8 | rat | Plantprobes | Arabinogalactan-proteins (AGPs) [52] |
| Extensins | LM1 | rat | Plantprobes | Angiosperm extensins (HRGP) [53] |
| mAb | Solanum Tuberosum | Ipomoea Batatas | Ipomoea Tricolor | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Epidermis | Collenchyma | Xylem | Phloem | Pith | Epidermis | Collenchyma | Xylem | Phloem | Pith | Epidermis | Collenchyma | Xylem | Phloem | Pith | ||
| Pectin | JIM5 | ++ | +++ | - | +++ | ++ 1 | ++ | +++ | - 2 | +++ | ++ | ++ | +++ | - 2 | +++ | ++ 1 |
| JIM7 | +++ | +++ | ++ | +++ | ++ | +++ | +++ | ++ | +++ | ++ | +++ | +++ | +- | +++ | ++ | |
| CCRC-M7 | + | +- 1 | - 3 | +++ | - | ++ | ++ 1,4 | + | +++ | + | ++ | ++ 4 | + | +++ | + | |
| LM6 | ++ | +++ | - | ++ | ++ | ++ 5 | +++ | ++ 6 | ++ | ++ | ++ 5 | +++ | ++ 6 | ++ | ++ | |
| Hemicellulose | CCRC-M138 | - 3 | - 3 | +++ 7,8 | +++ 9,10 | - | - | + | +++ 2,7,8 | +++ 9 | - | - | +- | +++ 2,7,8 | +++ 9 | - |
| LM25 unmasked | ++ | +++ | - | - | - | ++ | +++ | ++ | ++ | ++ | ++ | +++ | ++ | ++ | ++ | |
| LM21 unmasked | +- | + | - | - | - | +- | +- | + 7 | - | - | +- | - | - | - | - | |
| AGP | JIM8 | - | - | ++ 8 | - | - | + | - | ++ 8 | - | - | + | - | + 8 | - | - 3 |
| Extensins | LM1 | - 3 | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shtein, I.; Koyfman, A.; Schwartz, A.; Popper, Z.A.; Bar-On, B. Solanales Stem Biomechanical Properties Are Primarily Determined by Morphology Rather Than Internal Structural Anatomy and Cell Wall Composition. Plants 2020, 9, 678. https://doi.org/10.3390/plants9060678
Shtein I, Koyfman A, Schwartz A, Popper ZA, Bar-On B. Solanales Stem Biomechanical Properties Are Primarily Determined by Morphology Rather Than Internal Structural Anatomy and Cell Wall Composition. Plants. 2020; 9(6):678. https://doi.org/10.3390/plants9060678
Chicago/Turabian StyleShtein, Ilana, Alex Koyfman, Amnon Schwartz, Zoë A. Popper, and Benny Bar-On. 2020. "Solanales Stem Biomechanical Properties Are Primarily Determined by Morphology Rather Than Internal Structural Anatomy and Cell Wall Composition" Plants 9, no. 6: 678. https://doi.org/10.3390/plants9060678