Putative Silicon Transporters and Effect of Temperature Stresses and Silicon Supplementation on Their Expressions and Tissue Silicon Content in Poinsettia




Abstract
:1. Introduction
2. Results
2.1. Identification of Putative Genes Involved in Si Transport
2.2. Characteristics of the Puatative Si Transporters in Poinsettia
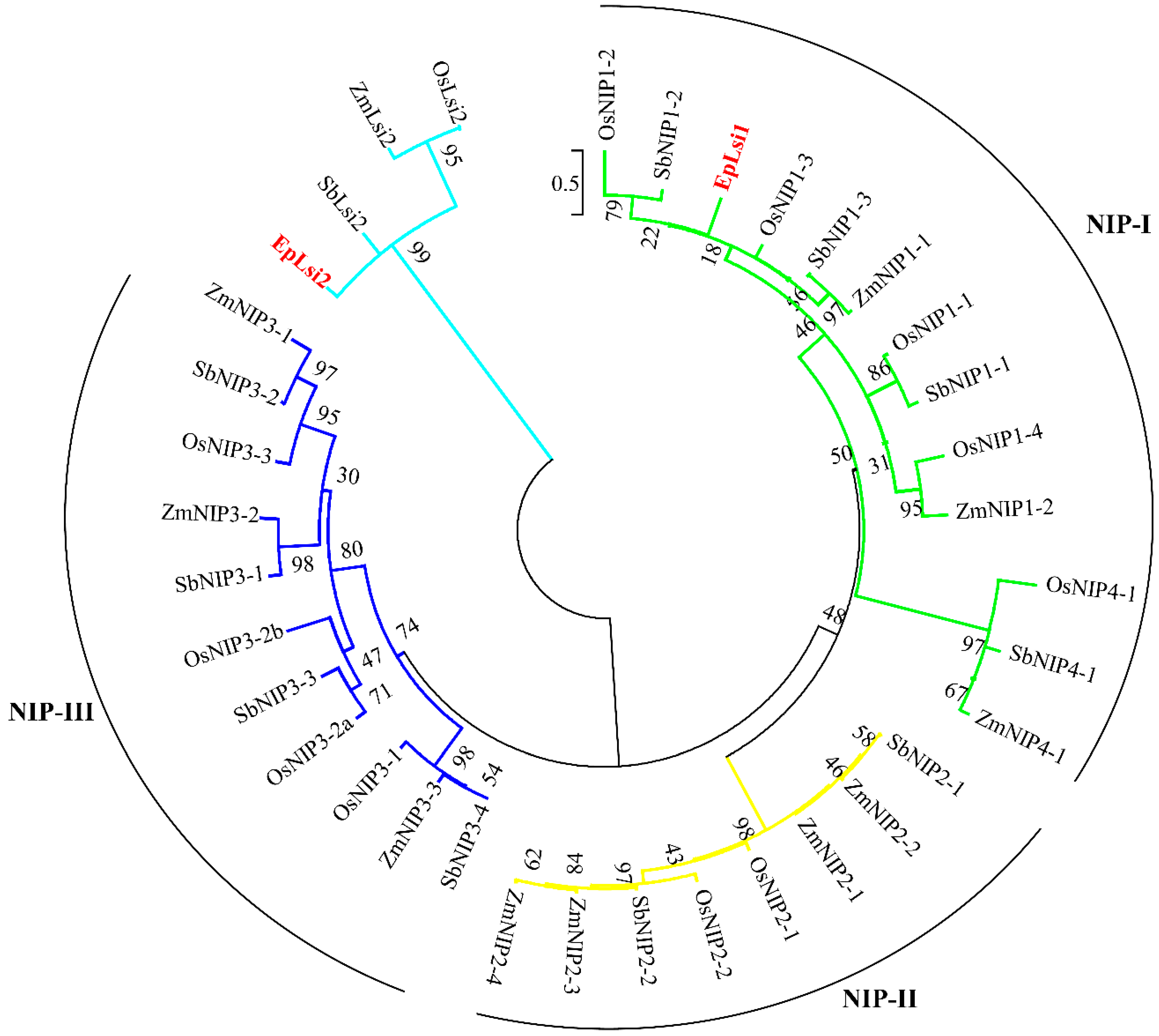
2.3. Phylogenetic Analysis of the Putative Si Transporters
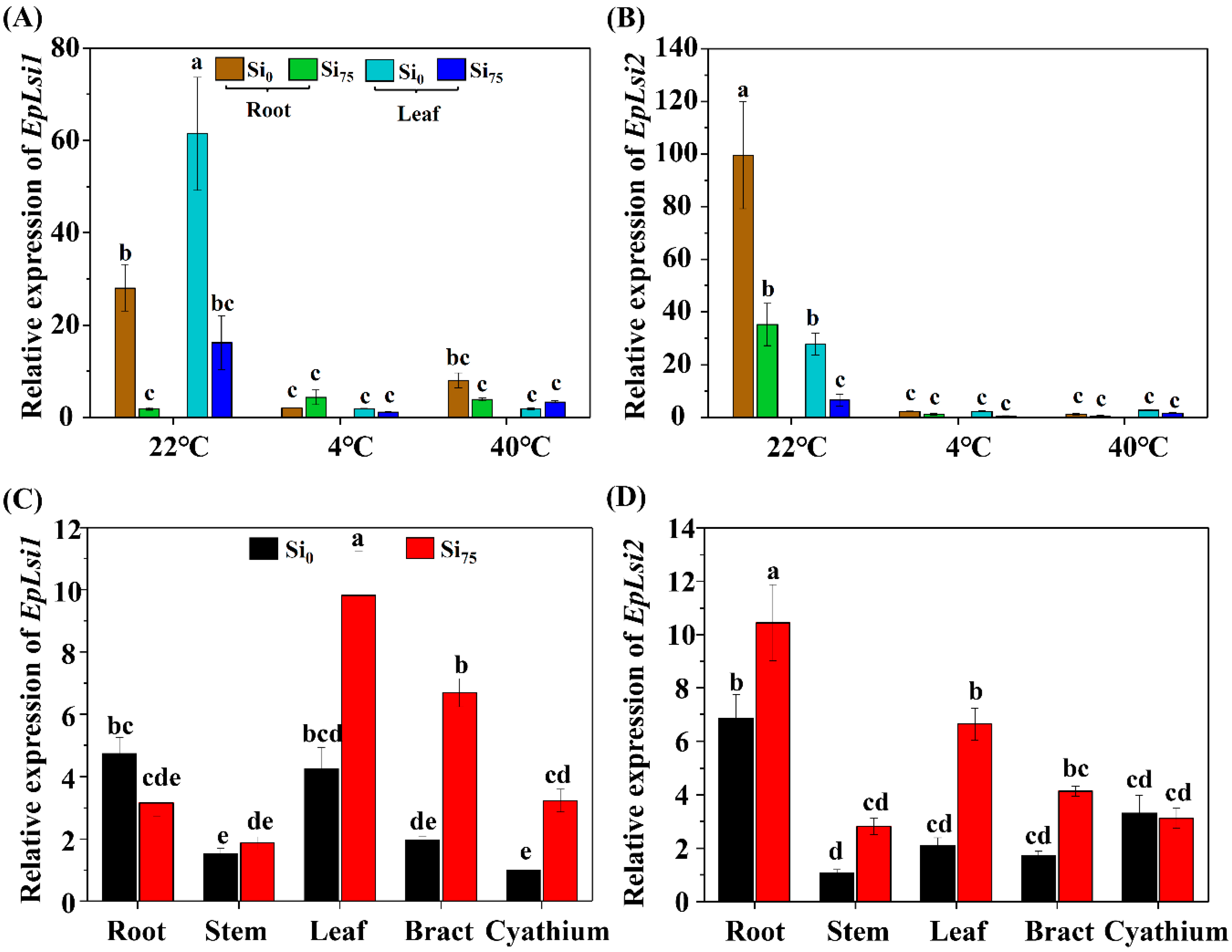
2.4. The Effects of Temperature Stresses and Si Supplementation on the Expressions of EpLsi1 and EpLsi2
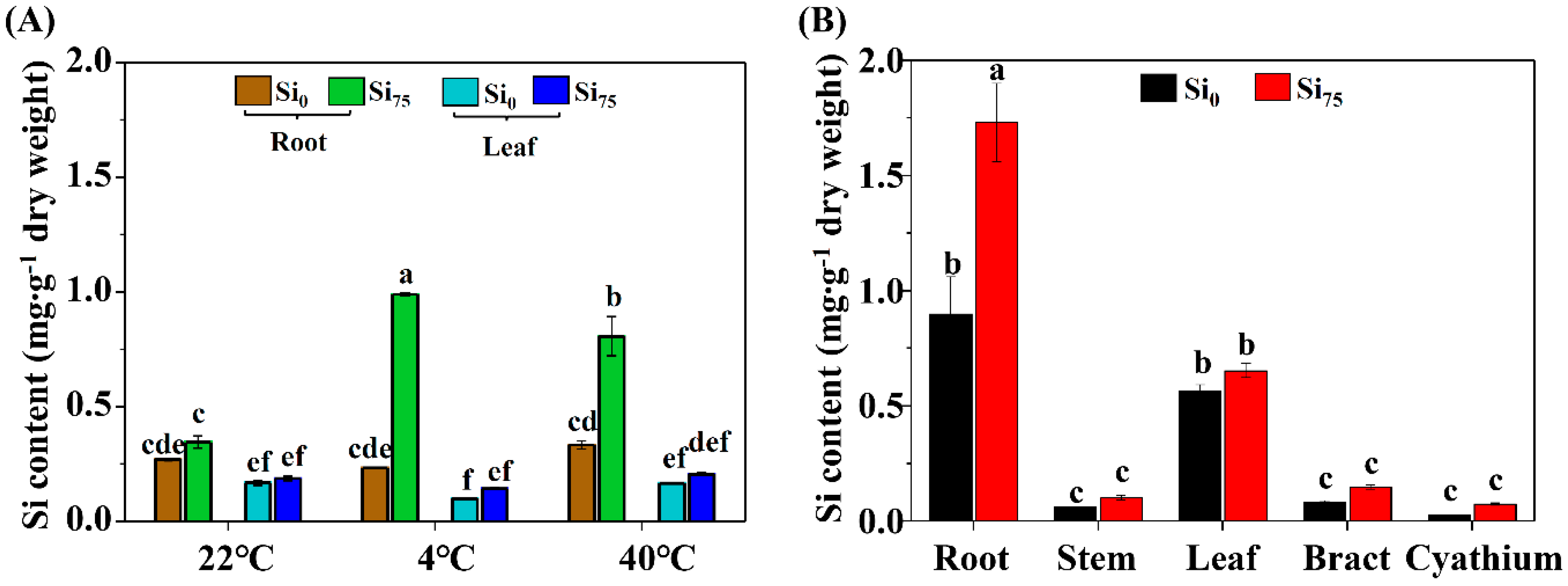
2.5. The Effects of Temperature Stresses and Si Supplementation on the Tissue Si Content
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. De novo Assembly and Identification of Putative Si Transporters in Poinsettia
4.3. Sequence Analysis of Si Transporters
4.4. Quantitative Real-Time PCR Analysis
4.5. Determination of the Si Content
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mattson, N.S.; Leatherwood, W.R. Potassium silicate drenches increase leaf silicon content and affect morphological traits of several floriculture crops grown in a peat-based substrate. HortScience 2010, 45, 43–47. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Cai, X.; Jeong, B.R. Silicon affects root development, tissue mineral content, and expression of silicon transporter genes in poinsettia (Euphorbia pulcherrima willd.) cultivars. Plants 2019, 8, 180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamenidou, S.; Cavins, T.J.; Marek, S. Silicon supplements affect horticultural traits of greenhouse-produced ornamental sunflowers. HortScience 2008, 43, 236–239. [Google Scholar] [CrossRef] [Green Version]
- Kamenidou, S.; Cavins, T.J.; Marek, S. Silicon supplements affect floricultural quality traits and elemental nutrient concentrations of greenhouse produced gerbera. Sci. Hortic. 2010, 123, 390–394. [Google Scholar] [CrossRef]
- Sivanesan, I.; Son, M.S.; Lee, J.P.; Jeong, B.R. Effects of silicon on growth of Tagetes patula L.‘Boy Orange’ and ‘Yellow Boy’seedlings cultured in an environment controlled chamber. Propag. Ornam. Plants 2010, 10, 136–140. [Google Scholar]
- Ma, J.F.; Tamai, K.; Yamaji, N.; Mitani, N.; Konishi, S.; Katsuhara, M.; Ishiguro, M.; Murata, Y.; Yano, M. A silicon transporter in rice. Nature 2006, 440, 688–691. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N.; Mitani, N.; Tamai, K.; Konishi, S.; Fujiwara, T.; Katsuhara, M.; Yano, M. An efflux transporter of silicon in rice. Nature 2007, 448, 209–212. [Google Scholar] [CrossRef]
- Wallace, I.S.; Roberts, D.M. Homology modeling of representative subfamilies of arabidopsis major intrinsic proteins. Classification based on the aromatic/arginine selectivity filter. Plant Physiol. 2004, 135, 1059–1068. [Google Scholar] [CrossRef] [Green Version]
- Mitani, N.; Yamaji, N.; Ma, J.F. Characterization of substrate specificity of a rice silicon transporter, Lsi1. Pflug. Arch. Eur. J. Phy. 2008, 456, 679–686. [Google Scholar] [CrossRef]
- Deshmukh, R.K.; Vivancos, J.; Ramakrishnan, G.; Guerin, V.; Carpentier, G.; Sonah, H.; Labbe, C.; Isenring, P.; Belzile, F.J.; Belanger, R.R. A precise spacing between the npa domains of aquaporins is essential for silicon permeability in plants. Plant J. 2015, 83, 489–500. [Google Scholar] [CrossRef]
- Mitani, N.; Yamaji, N.; Ago, Y.; Iwasaki, K.; Ma, J.F. Isolation and functional characterization of an influx silicon transporter in two pumpkin cultivars contrasting in silicon accumulation. Plant J. 2011, 66, 231–240. [Google Scholar] [CrossRef]
- Luyckx, M.; Hausman, J.F.; Lutts, S.; Guerriero, G. Silicon and plants: Current knowledge and technological perspectives. Front. Plant Sci. 2017, 8, 411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soundararajan, P.; Sivanesan, I.; Jana, S.; Jeong, B.R. Influence of silicon supplementation on the growth and tolerance to high temperature in Salvia splendens. Hortic. Environ. Biote. 2014, 55, 271–279. [Google Scholar] [CrossRef]
- He, Y.; Xiao, H.; Wang, H.; Chen, Y.; Yu, M. Effect of silicon on chilling-induced changes of solutes, antioxidants, and membrane stability in seashore paspalum turfgrass. Acta Physiol. Plant. 2010, 32, 487–494. [Google Scholar] [CrossRef]
- HABIBI, G. Effect of foliar-applied silicon on photochemistry, antioxidant capacity and growth in maize plants subjected to chilling stress. Acta Agric. Slov. 2016, 107, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.F.; Tamai, K.; Ichii, M.; Wu, G.F. A rice mutant defective in si uptake. Plant Phy. 2002, 130, 2111–2117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Y.; Si, J.; Romheld, V. Silicon uptake and transport is an active process in Cucumis sativus. New Phytol. 2005, 167, 797–804. [Google Scholar] [CrossRef]
- Chiba, Y.; Mitani, N.; Yamaji, N.; Ma, J.F. Hvlsi1 is a silicon influx transporter in barley. Plant J. 2009, 57, 810–818. [Google Scholar] [CrossRef]
- Sun, H.; Guo, J.; Duan, Y.; Zhang, T.; Huo, H.; Gong, H. Isolation and functional characterization of CsLsi1, a silicon transporter gene in Cucumis sativus. Physiol. Plantarum 2017, 159, 201–214. [Google Scholar] [CrossRef]
- Wang, H.-S.; Yu, C.; Fan, P.-P.; Bao, B.-F.; Li, T.; Zhu, Z.-J. Identification of two cucumber putative silicon transporter genes in Cucumis sativus. J. Plant Growth Regul. 2015, 34, 332–338. [Google Scholar] [CrossRef]
- Mitani, N.; Chiba, Y.; Yamaji, N.; Ma, J.F. Identification and characterization of maize and barley Lsi2-like silicon efflux transporters reveals a distinct silicon uptake system from that in rice. Plant Cell 2009, 21, 2133–2142. [Google Scholar] [CrossRef] [Green Version]
- Mitani, N.; Yamaji, N.; Ma, J.F. Identification of maize silicon influx transporters. Plant Cell Physiol. 2009, 50, 5–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bokor, B.; Ondos, S.; Vaculik, M.; Bokorova, S.; Weidinger, M.; Lichtscheidl, I.; Turna, J.; Lux, A. Expression of genes for Si uptake, accumulation, and correlation of Si with other elements in ionome of maize kernel. Front. Plant Sci. 2017, 8, 1063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Melo, S.P.; Monteiro, F.A.; De Bona, F.D. Silicon distribution and accumulation in shoot tissue of the tropical forage grass Brachiaria brizantha. Plant Soil 2010, 336, 241–249. [Google Scholar] [CrossRef]
- Vatansever, R.; Ozyigit, I.I.; Filiz, E.; Gozukirmizi, N. Genome-wide exploration of silicon (Si) transporter genes, Lsi1 and Lsi2 in plants; insights into si-accumulation status/capacity of plants. Biometals 2017, 30, 185–200. [Google Scholar] [CrossRef] [PubMed]
- Wallace, I.S.; Roberts, D.M. Distinct transport selectivity of two structural subclasses of the nodulin-like intrinsic protein family of plant aquaglyceroporin channels. Biochemistry 2005, 44, 16826–16834. [Google Scholar] [CrossRef] [PubMed]
- Epstein, E. The anomaly of silicon in plant biology. Proc. Natl. Acad. Sci. USA 1994, 91, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Montpetit, J.; Vivancos, J.; Mitani-Ueno, N.; Yamaji, N.; Remus-Borel, W.; Belzile, F.; Ma, J.F.; Belanger, R.R. Cloning, functional characterization and heterologous expression of TaLsi1, a wheat silicon transporter gene. Plant Mol. Biol. 2012, 79, 35–46. [Google Scholar] [CrossRef]
- Kuehny, J.; Branch, P.; Adams, P. Stem strength of poinsettia. Acta Hortic. 2000, 515, 257–264. [Google Scholar] [CrossRef]
- Stevens, T.J., 3rd; Kilmer, R.L.; Glenn, S.J. An economic comparison of biological and conventional control strategies for whiteflies (Homoptera: Aleyrodidae) in greenhouse poinsettias. J. Econ. Entomol. 2000, 93, 623–629. [Google Scholar] [CrossRef] [Green Version]
- Isa, M.; Bai, S.; Yokoyama, T.; Ma, J.F.; Ishibashi, Y.; Yuasa, T.; Iwaya-Inoue, M. Silicon enhances growth independent of silica deposition in a low-silica rice mutant, Lsi1. Plant Soil 2010, 331, 361–375. [Google Scholar] [CrossRef]
- Alhousari, F.; Greger, M. Silicon and mechanisms of plant resistance to insect pests. Plants 2018, 7, 33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meharg, C.; Meharg, A.A. Silicon, the silver bullet for mitigating biotic and abiotic stress, and improving grain quality, in rice? Environ. Exp. Bot. 2015, 120, 8–17. [Google Scholar] [CrossRef] [Green Version]
- McAvoy, R.J.; Bible, B.B. Silica sprays reduce the incidence and severity of bract necrosis in poinsettia. HortScience 1996, 31, 1146–1149. [Google Scholar] [CrossRef]
- Laane, H.M. The effects of foliar sprays with different silicon compounds. Plants 2018, 7, 45. [Google Scholar] [CrossRef] [Green Version]
- Bokor, B.; Bokorova, S.; Ondos, S.; Svubova, R.; Lukacoca, Z.; Hyblova, M.; Szemes, T.; Lux, A. Ionome and expression level of Si transporter genes (Lsi1, Lsi2, and Lsi6) affected by Zn and Si interaction in maize. Environ. Sci. Pollut. Res. 2015, 22, 6800–6811. [Google Scholar] [CrossRef]
- Raven, J. Silicon transport at the cell and tissue level. In Studies in Plant Science; Elsevier: Amsterdam, The Netherlands, 2001; Volume 8, pp. 41–55. [Google Scholar]
- Bauer, P.; Elbaum, R.; Weiss, I.M. Calcium and silicon mineralization in land plants: Transport, structure and function. Plant Sci. 2011, 180, 746–756. [Google Scholar] [CrossRef]
- Galaxy. Available online: https://usegalaxy.org/ (accessed on 21 July 2018).
- Phytozome. Available online: https://phytozome.jgi.doe.gov/pz/portal.html (accessed on 16 March 2019).
- Phyre2. Available online: http://www.sbg.bio.ic.ac.uk/phyre2/html/ (accessed on 18 March 2019).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Gene ID | E-Value | Length (bp) | Query | Positives z |
|---|---|---|---|---|---|
| Si influx transporter | DN37620_c0_g2_i1 | 6E-066 | 1006 | LOC_Os02g51110.1 | 149/234 (64%) |
| DN37981_c1_g4_i1 | 1E-043 | 779 | GRMZM2G028325_T01 | 101/153 (66%) | |
| DN25780_c0_g2_i1 | 2E-027 | 243 | LOC_Os02g51110.1 | 60/78 (77%) | |
| DN14261_c0_g1_i1 | 1E-026 | 302 | GRMZM2G081239_T01 | 64/98 (65%) | |
| DN37981_c1_g1_i1 | 4E-017 | 850 | LOC_Os02g51110.1 | 59/104 (57%) | |
| DN37620_c0_g2_i2 | 2E-016 | 525 | LOC_Os02g51110.1 | 40/56 (71%) | |
| DN37981_c1_g2_i1 | 1E-015 | 479 | GRMZM2G028325_T01 | 46/71 (65%) | |
| DN25780_c0_g3_i1 | 5E-010 | 235 | LOC_Os02g51110.1 | 28/34 (82%) | |
| DN25780_c0_g1_i1 | 5E-009 | 251 | Sobic.010G092600.1 | 33/43 (77%) | |
| DN49493_c0_g1_i1 | 1E-004 | 304 | LOC_Os02g51110.1 | 29/53 (55%) | |
| Si efflux transporter | DN44911_c1_g2_i1 | 0 | 1982 | Sobic.004G350000.1 | 402/528 (76%) |
| DN11282_c0_g2_i1 | 4E-034 | 476 | GRMZM2G158682_T01 | 98/116 (84%) |
| Gene ID z | Peptide Length (AA) | ORF Integrity | Protein Domain Family y | Annotation x |
|---|---|---|---|---|
| DN37620_c0_g2_i1 | 273 | Complete | Major intrinsic protein | NIP1-2 |
| DN37981_c1_g4_i1 | 152 | Partial | Major intrinsic protein | NIP5-1 |
| DN25780_c0_g2_i1 | 80 | Partial | Major intrinsic protein | NIP1-2 |
| DN14261_c0_g1_i1 | 100 | Partial | Major intrinsic protein | NIP6-1 |
| DN37981_c1_g1_i1 | 153 | Partial | Major intrinsic protein | NIP5-1 |
| DN37620_c0_g2_i2 | 81 | Partial | Major intrinsic protein | NIP1-2 |
| DN37981_c1_g2_i1 | 111 | Partial | Major intrinsic protein | NIP5-1 |
| DN25780_c0_g3_i1 | 63 | Partial | - | NIP1 |
| DN25780_c0_g1_i1 | 43 | Partial | - | NIP1-2 |
| DN49493_c0_g1_i1 | 57 | Partial | - | NIP2-1 |
| DN44911_c1_g2_i1 | 523 | Complete | Citrate transporter | Si efflux transporter |
| DN11282_c0_g2_i1 | 128 | Partial | Citrate transporter | Si efflux transporter |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Li, Y.; Jeong, B.R. Putative Silicon Transporters and Effect of Temperature Stresses and Silicon Supplementation on Their Expressions and Tissue Silicon Content in Poinsettia. Plants 2020, 9, 569. https://doi.org/10.3390/plants9050569
Hu J, Li Y, Jeong BR. Putative Silicon Transporters and Effect of Temperature Stresses and Silicon Supplementation on Their Expressions and Tissue Silicon Content in Poinsettia. Plants. 2020; 9(5):569. https://doi.org/10.3390/plants9050569
Chicago/Turabian StyleHu, Jiangtao, Yali Li, and Byoung Ryong Jeong. 2020. "Putative Silicon Transporters and Effect of Temperature Stresses and Silicon Supplementation on Their Expressions and Tissue Silicon Content in Poinsettia" Plants 9, no. 5: 569. https://doi.org/10.3390/plants9050569