Eco-Physiological Traits Related to Recovery from Complete Submergence in the Model Legume Lotus japonicus

 , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Plant Growth during Submergence
2.2. Plant Recovery from Submergence
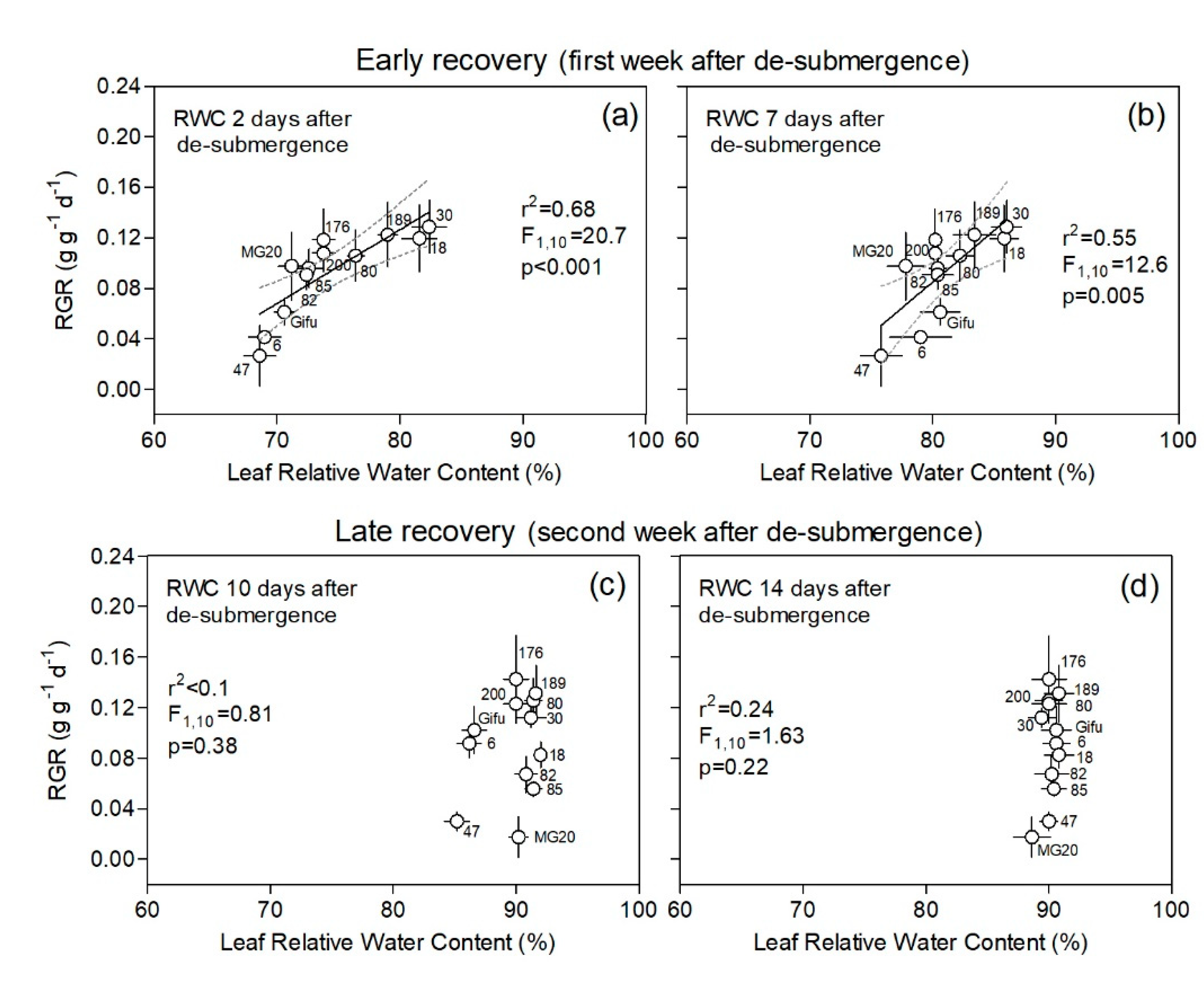
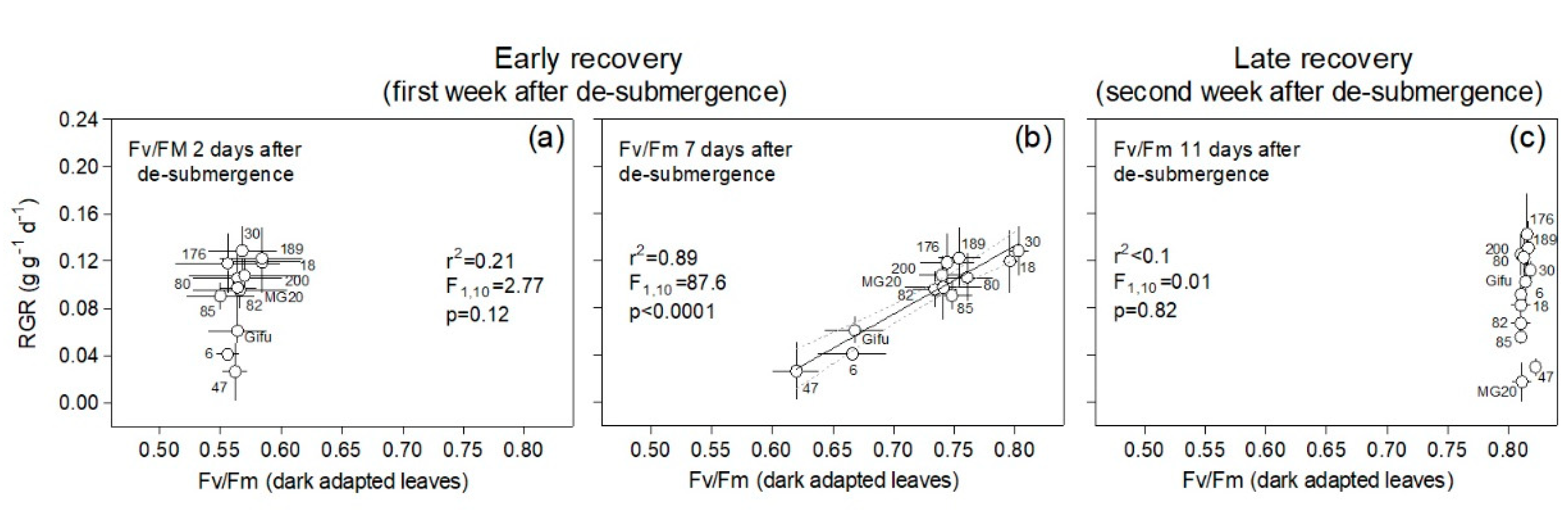
2.3. Leaf Physiological Variables Related to Plant Recovery
2.4. Correlations among Traits Aiding Plant Recovery following Submergence
3. Discussion
4. Materials and Methods
4.1. Species Description
4.2. Plant Material and Growing Conditions
4.3. Experimental Design
4.4. Dry Mass and Relative Growth Rate
4.5. Leaf Physiological Responses: Relative Water Content, Stomatal Conductance, Chlorophyll Fluorescence, and Greenness
4.6. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IPCC. 2014. Climate Change 2014: Synthesis Report. Contribution of Working Groups I. Available online: https://www.ipcc.ch/ (accessed on 20 March 2020).
- Voesenek, L.A.C.J.; Colmer, T.D.; Pierik, R.; Millenaar, F.F.; Peeters, A.J.M. How plants cope with complete submergence. New Phytol. 2006, 170, 213–226. [Google Scholar] [CrossRef] [Green Version]
- Colmer, T.D.; Voesenek, L.A.C.J. Flooding tolerance: Suites of plant traits in variable environments. Funct. Plant Biol. 2009, 36, 665–681. [Google Scholar] [CrossRef]
- Bailey-Serres, J.; Voesenek, L.A.C.J. Flooding stress: Acclimations and genetic diversity. Annu. Rev. Plant Biol. 2008, 59, 313–339. [Google Scholar] [CrossRef] [Green Version]
- Bailey-Serres, J.; Voesenek, L.A.C.J. Life in the balance: A signaling network controlling survival of flooding. Curr. Opin. Plant Biol. 2010, 13, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Striker, G.G.; Izaguirre, R.F.; Manzur, M.E.; Grimoldi, A.A. Different strategies of Lotus japonicus, L. corniculatus and L. tenuis to deal with complete submergence at seedling stage. Plant Biol. 2012, 14, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Voesenek, L.A.C.J.; Bailey-Serres, J. Flooding tolerance: O2 sensing and survival strategies. Curr. Opin. Plant Biol. 2013, 16, 647–653. [Google Scholar] [CrossRef]
- Striker, G.G. Time is on our side: The importance of considering a recovery period when assessing flooding tolerance in plants. Ecol. Res. 2012, 27, 983–987. [Google Scholar] [CrossRef]
- Fukao, T.; Xu, K.; Ronald, P.C.; Bailey-Serres, J. A variable cluster of ethylene response factor–like genes regulates metabolic and developmental acclimation responses to submergence in rice. Plant Cell 2006, 18, 2021–2034. [Google Scholar] [CrossRef] [Green Version]
- Hattori, Y.; Nagai, K.; Furukawa, S.; Song, X.J.; Kawano, R.; Sakakibara, H.; Wu, J.; Matsumoto, T.; Yoshimura, A.; Kitano, H.; et al. The ethylene response factors SNORKEL1 and SNORKEL2 allow rice to adapt to deep water. Nature 2009, 460, 1026–1030. [Google Scholar] [CrossRef]
- Nagai, K.; Hattori, Y.; Ashikari, M. Stunt or elongate? Two opposite strategies by which rice adapts to floods. J. Plant Res. 2010, 123, 303–309. [Google Scholar] [CrossRef]
- Fukao, T.; Barrera-Figueroa, B.E.; Juntawong, P.; Peña-Castro, J.M. Submergence and waterlogging stress in plants: A review highlighting research opportunities and understudied aspects. Front. Plant Sci. 2019, 10, 340. [Google Scholar] [CrossRef] [PubMed]
- Yeung, E.; van Veen, H.; Vashisht, D.; Paiva, A.L.S.; Hummel, M.; Rankenberg, T.; Steffens, B.; Steffen-Heins, A.; Sauter, M.; de Vries, M.; et al. A stress recovery signaling network for enhanced flooding tolerance in Arabidopsis thaliana. PNAS 2018, 115, E6085–E6094. [Google Scholar] [CrossRef] [Green Version]
- Yeung, E.; Bailey-Serres, J.; Sasidharan, R. After the deluge: Plant revival post-flooding. Trends Plant Sci. 2019, 24, 443–454. [Google Scholar] [CrossRef] [PubMed]
- Pajuelo, E.; Stougaard, J. Lotus japonicus as a model system. In Lotus japonicus handbook; Stougaard, J., Márquez, A.J., Eds.; Springer: Dordrecht, The Netherlands, 2005; pp. 3–24. [Google Scholar]
- Escaray, F.J.; Menendez, A.B.; Gárriz, A.; Pieckenstain, F.L.; Estrella, M.J.; Castagno, L.N.; Carrasco, P.; Sanjuan, J.; Ruiz, O.A. Ecological and agronomic importance of the plant genus Lotus. Its application in grassland sustainability and the amelioration of constrained and contaminated soils. Plant Sci. 2012, 182, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Striker, G.G.; Colmer, T.D. Flooding tolerance of forage legumes. J. Exp. Bot. 2017, 68, 1851–1872. [Google Scholar] [CrossRef] [Green Version]
- James, E.K.; Crawford, R.M.M. Effect of oxygen availability on nitrogen fixation by two Lotus species under flooded conditions. J. Exp. Bot. 1998, 49, 599–609. [Google Scholar] [CrossRef]
- Striker, G.G.; Insausti, P.; Grimoldi, A.A.; Ploschuk, E.L.; Vasellati, V. Physiological and anatomical basis of differential tolerance to soil flooding of Lotus corniculatus, L. and Lotus glaber Mill. Plant Soil 2005, 276, 301–311. [Google Scholar] [CrossRef]
- Young, N.D.; Cannon, S.B.; Sato, S.; Kim, D.; Cook, D.R.; Town, C.D.; Roe, B.A.; Tabata, S. Sequencing the genespaces of Medicago truncatula and Lotus japonicus. Plant Physiol. 2005, 137, 1174–1181. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Q.; Gresshoff, P.M. Classical and molecular genetics of the model legume Lotus japonicus. Mol. Plant Microbe Interact. 1997, 10, 59–68. [Google Scholar] [CrossRef] [Green Version]
- Mun, T.; Bachmann, A.; Gupta, V.; Stougaard, J.; Andersen, S.U. Lotus Base: An integrated information portal for the model legume Lotus japonicus. Sci. Rep. 2016, 6, 39447. [Google Scholar] [CrossRef] [Green Version]
- Striker, G.G.; Casas, C.; Manzur, M.E.; Ploschuk, R.A.; Casal, J.J. Phenomic networks reveal largely independent root and shoot adjustment in waterlogged plants of Lotus japonicus. Plant Cell Environ. 2014, 37, 2278–2293. [Google Scholar] [PubMed]
- Striker, G.G.; Casas, C.; Kuang, X.; Grimoldi, A.A. No escape? Costs and benefits of leaf de-submergence in the pasture grass Chloris gayana under different flooding regimes. Funct. Plant Biol. 2017, 44, 899–906. [Google Scholar] [CrossRef]
- Setter, T.L.; Bhekasut, P.; Greenway, H. Desiccation of leaves after de-submergence is one cause for intolerance to complete submergence of the rice cultivar IR 42. Funct. Plant Biol. 2010, 37, 1096–1104. [Google Scholar] [CrossRef]
- Luo, F.L.; Nagel, K.A.; Zeng, B.; Schurr, U.; Matsubara, S. Photosynthetic acclimation is important for post-submergence recovery of photosynthesis and growth in two riparian species. Ann. Bot. 2009, 104, 1435–1444. [Google Scholar] [CrossRef] [PubMed]
- Luo, F.L.; Nagel, K.A.; Scharr, H.; Zeng, B.; Schurr, U.; Matsubara, S. Recovery dynamics of growth, photosynthesis and carbohydrate accumulation after de-submergence: A comparison between two wetland plants showing escape and quiescence strategies. Ann. Bot. 2011, 107, 49–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Setter, T.L.; Laureles, E.V. The beneficial effect of reduced elongation growth on submergence tolerance of rice. J. Exp. Bot. 1996, 47, 1551–1559. [Google Scholar] [CrossRef] [Green Version]
- Hattori, Y.; Nagai, K.; Ashikari, M. Rice growth adapting to deepwater. Curr. Opin. Plant Biol. 2011, 14, 100–105. [Google Scholar] [CrossRef]
- Zaman, M.S.U.; Malik, A.I.; Erskine, W.; Kaur, P. Changes in gene expression during germination reveal pea genotypes with either “quiescence” or “escape” mechanisms of waterlogging tolerance. Plant Cell Environ. 2019, 42, 245–258. [Google Scholar] [CrossRef] [Green Version]
- Manzur, M.E.; Grimoldi, A.A.; Insausti, P.; Striker, G.G. Escape from water or remain quiescent? Lotus tenuis changes its strategy depending on depth of submergence. Ann. Bot. 2009, 104, 1163–1169. [Google Scholar]
- Tanguilig, V.C.; Yambao, E.B.; O’toole, J.C.; De Datta, S.K. Water stress effects on leaf elongation, leaf water potential, transpiration, and nutrient uptake of rice, maize, and soybean. Plant Soil 1987, 103, 155–168. [Google Scholar] [CrossRef]
- Aroca, R.; Porcel, R.; Ruiz-Lozano, J.M. Regulation of root water uptake under abiotic stress conditions. J. Exp. Bot. 2012, 63, 43–57. [Google Scholar]
- Herzog, M.; Pedersen, O. Partial versus complete submergence: Snorkelling aids root aeration in Rumex palustris but not in R. acetosa. Plant Cell Environ. 2014, 37, 2381–2390. [Google Scholar]
- Pan, Y.; Cieraad, E.; Clarkson, B.R.; Colmer, T.D.; Pedersen, O.; Visser, E.J.W.; Voesenek, L.A.C.J.; van Bodegom, P.M. Drivers of plant traits that allow survival in wetlands. Funct. Ecol. 2020. [Google Scholar] [CrossRef]
- Cakmak, I.; Hengeler, C.; Marschner, H. Partitioning of shoot and root dry matter and carbohydrates in bean plants suffering from phosphorus, potassium and magnesium deficiency. J. Exp. Bot. 1994, 45, 1245–1250. [Google Scholar] [CrossRef]
- Andrews, M.; Sprent, J.I.; Raven, J.A.; Eady, P.E. Relationships between shoot to root ratio, growth and leaf soluble protein concentration of Pisum sativum, Phaseolus vulgaris and Triticum aestivum under different nutrient deficiencies. Plant Cell Environ. 1999, 22, 949–958. [Google Scholar] [CrossRef]
- Striker, G.G.; Ploschuk, R.A. Recovery from short-term complete submergence in temperate pasture grasses. Crop Pasture Sci. 2018, 69, 745–753. [Google Scholar] [CrossRef]
- Mielke, M.S.; de Almeida, A.A.F.; Gomes, F.P.; Aguilar, M.A.G.; Mangabeira, P.A.O. Leaf gas exchange, chlorophyll fluorescence and growth responses of Genipa americana seedlings to soil flooding. Environ. Exp. Bot. 2003, 50, 221–231. [Google Scholar] [CrossRef]
- Ploschuk, R.A.; Grimoldi, A.A.; Ploschuk, E.L.; Striker, G.G. Growth during recovery evidences the waterlogging tolerance of forage grasses. Crop Pasture Sci. 2017, 68, 574–582. [Google Scholar] [CrossRef]
- Blokhina, O.; Virolainen, E.; Fagerstedt, K.V. Antioxidants, oxidative damage and oxygen deprivation stress: A review. Ann. Bot. 2003, 91, 179–194. [Google Scholar] [CrossRef] [Green Version]
- Vriet, C.; Smith, A.M.; Wang, T.L. Root starch reserves are necessary for vigorous re-growth following cutting back in Lotus japonicus. PLoS ONE 2014, 9, e87333. [Google Scholar] [CrossRef]
- Swanson, E.B.; Somers, D.A.; Tomes, D.T. Birdsfoot trefoil (Lotus corniculatus L.). In Legumes and Oilseed Crops I; Springer: Berlin/Heidelberg, Germany, 1990; pp. 323–340. [Google Scholar]
- Kawaguchi, M.; Motomura, T.; Imaizumi-Anraku, H.; Akao, S.; Kawasaki, S. Providing the basis for genomics in Lotus japonicus: The accessions Miyakojima and Gifu are appropriate crossing partners for genetic analyses. Mol Gen. Genom. 2001, 266, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Striker, G.G.; Kotula, L.; Colmer, T.D. Tolerance to partial and complete submergence in the forage legume Melilotus siculus: An evaluation of 15 accessions for petiole hyponastic response and gas-filled spaces, leaf hydrophobicity and gas films, and root phellem. Ann. Bot. 2019, 123, 169–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colmer, T.D.; Pedersen, O. Underwater photosynthesis and respiration in leaves of submerged wetland plants: Gas films improve CO2 and O2 exchange. New Phytol. 2008, 177, 918–926. [Google Scholar] [CrossRef] [PubMed]
- Hunt, R. Plant growth curves. In The Functional Approach to Plant Growth Analysis; Edward Arnold Ltd.: London, UK, 1982. [Google Scholar]
- Čatský, J. Determination of water deficit in disks cut out from leaf blades. Biol. Plant. 1960, 2, 76. [Google Scholar] [CrossRef]
- Maxwell, K.; Johnson, G.N. Chlorophyll fluorescence—A practical guide. J. Exp. Bot. 2000, 51, 659–668. [Google Scholar] [CrossRef] [PubMed]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; González, L.; Tablada, M.; Robledo, Y.C. InfoStat versión 2011. Grupo InfoStat, FCA, Universidad Nacional de Córdoba, Argentina. Available online: http://www.infostat.com.ar (accessed on 20 March 2020).








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Controls ► Recovering from sub. ▼ | S:R | LRWC 2-d Recovery | LRWC 7-d Recovery | gs 2-d Recovery | gs 7-d Recovery | Fv/Fm 7-d Recovery | Greenness Basal Leaves 2-d Recovery | Greenness Apical Leaves 2-d Recovery | Greenness Basal Leaves 7-d Recovery | Greenness Apical Leaves 7-d Recovery |
|---|---|---|---|---|---|---|---|---|---|---|
| S:R | 0.15 ns | 0.22 ns | 0.18 ns | −0.02 ns | 0.50 ns | −0.04 ns | −0.18 ns | −0.21 ns | 0.27 ns | |
| LRWC 2-d recovery | −0.69 * | 0.31 ns | 0.22 ns | 0.16 ns | −0.22 ns | 0.38 ns | −0.17 ns | −0.20 ns | −0.14 ns | |
| LRWC 7-d recovery | −0.78 ** | 0.94 ** | 0.07 ns | 0.26 ns | −0.29 ns | 0.13 ns | 0.33 ns | −0.26 ns | 0.25 ns | |
| gs 2-d recovery | −0.62 * | 0.91 ** | 0.85 ** | 0.54 ns | 0.25 ns | 0.31 ns | −0.21 ns | −0.28 ns | 0.58 * | |
| gs 7-d recovery | −0.51 ns | 0.75 * | 0.73 * | 0.85 ** | −0.15 ns | 0.45 ns | 0.30 ns | 0.04 ns | 0.43 ns | |
| Fv/Fm 7-d recovery | −0.54 ns | 0.85 ** | 0.80 ** | 0.93 ** | 0.95 ** | −0.03 ns | −0.09 ns | −0.24 ns | 0.31 ns | |
| Greenness basal leaves 2-d recovery | −0.51 ns | 0.86 ** | 0.76 * | 0.95 ** | 0.88 ** | 0.92 ** | −0.52 ns | 0.22 ns | −0.08 ns | |
| Greenness apical leaves 2-d recovery | −0.44 ns | 0.68 * | 0.60 * | 0.83 ** | 0.92 ** | 0.93 ** | 0.87 ** | 0.40 ns | −0.72 * | |
| Greenness basal leaves 7-d recovery | −0.58 * | 0.92 ** | 0.85 ** | 0.88 ** | 0.79 * | 0.86 ** | 0.87 ** | 0.75 * | 0.26 ns | |
| Greenness apical leaves 7-d recovery | −0.46 ns | 0.80 ** | 0.68 * | 0.90 ** | 0.88 ** | 0.95 ** | 0.91 ** | 0.94 ** | 0.84 ** |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Buraschi, F.B.; Mollard, F.P.O.; Grimoldi, A.A.; Striker, G.G. Eco-Physiological Traits Related to Recovery from Complete Submergence in the Model Legume Lotus japonicus. Plants 2020, 9, 538. https://doi.org/10.3390/plants9040538
Buraschi FB, Mollard FPO, Grimoldi AA, Striker GG. Eco-Physiological Traits Related to Recovery from Complete Submergence in the Model Legume Lotus japonicus. Plants. 2020; 9(4):538. https://doi.org/10.3390/plants9040538
Chicago/Turabian StyleBuraschi, Florencia B., Federico P.O. Mollard, Agustín A. Grimoldi, and Gustavo G. Striker. 2020. "Eco-Physiological Traits Related to Recovery from Complete Submergence in the Model Legume Lotus japonicus" Plants 9, no. 4: 538. https://doi.org/10.3390/plants9040538