Tolerance of Douglas Fir Somatic Plantlets to Aluminum Stress: Biological, Cytological, and Mineral Studies


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of Al on the Growth of Douglas Fir Somatic Plantlets
2.2. SEM Observations of Root Tip Morphology
2.3. Distribution of Al and Minerals in Different Organs of Douglas Fir Somatic Plantlets
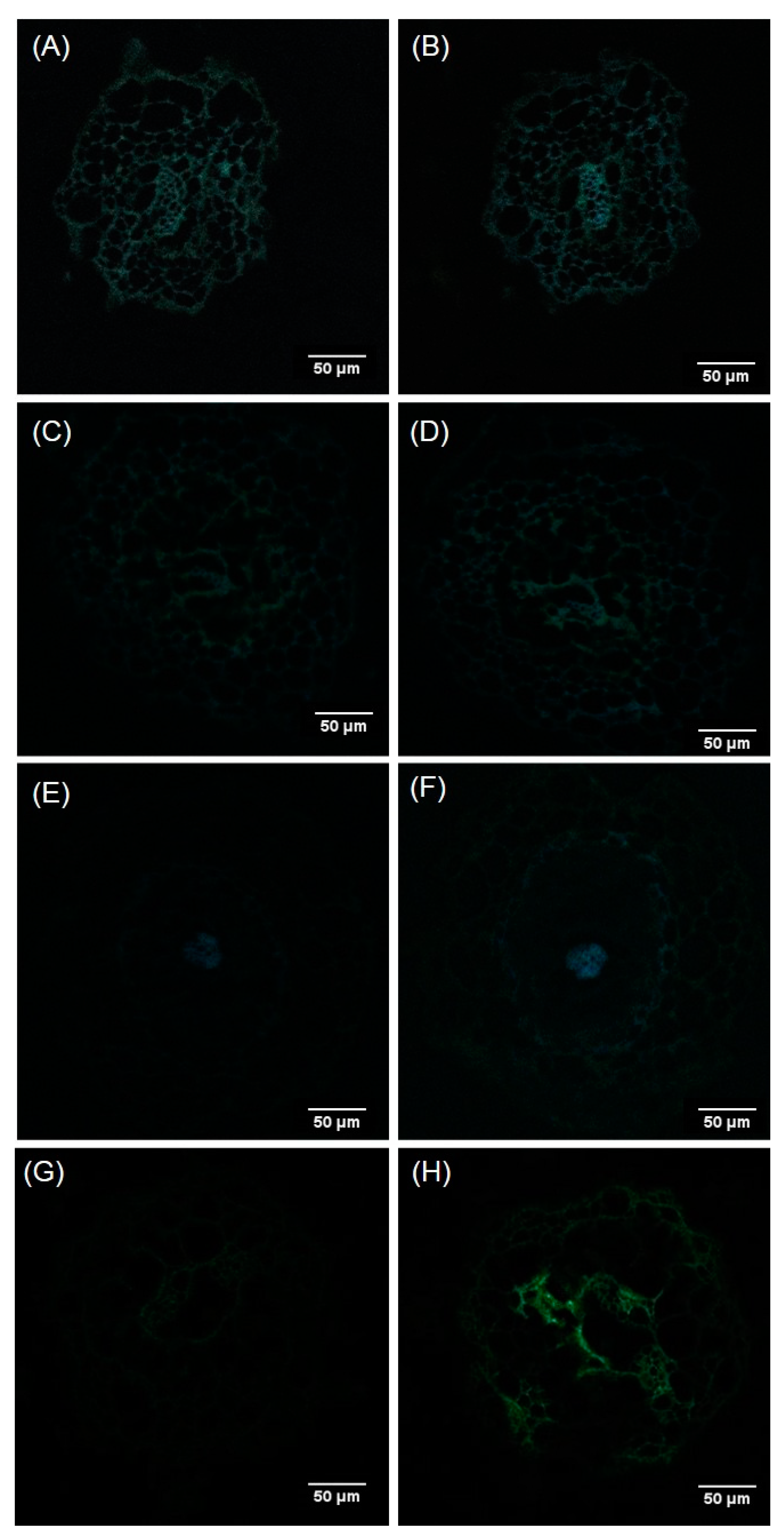
2.4. Localization of Al in the Roots of Douglas Fir Somatic Plantlets
2.5. Impact of Al on Mineral Content of Douglas Fir Somatic Plantlets
3. Discussion
4. Materials and methods
4.1. Plant Material and Growth Conditions
4.2. Aluminum Treatment
4.3. Micromorphology Analysis of Root Tips by Scanning Electron Microscopy
4.4. Microscopic Examination of Roots
4.4.1. Calcofluor White Staining
4.4.2. Morin Staining
4.5. Mineral Content and Al Quantification
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Eckhart, T.; Pötzelsberger, E.; Koeck, R.; Thom, D.; Lair, G.J.; van Loo, M.; Hasenauer, H. Forest stand productivity derived from site conditions: An assessment of old Douglas-fir stands (Pseudotsuga menziesii (Mirb.) Franco var. menziesii) in Central Europe. Ann. For. Sci. 2019, 76, 19. [Google Scholar] [CrossRef] [Green Version]
- Bojórquez-Quintal, E.; Escalante-Magaña, C.; Echevarría-Machado, I.; Martínez-Estévez, M. Aluminum, a friend or foe of higher plants in acid soils. Front. Plant Sci. 2017, 8, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Batista, M.F.; Moscheta, I.S.; Bonato, C.M.; Batista, M.A.; Almeida, O.J.G.d.; Inoue, T.T. Aluminum in corn plants: Influence on growth and morpho-anatomy of root and leaf. Rev. Bras. Ciênc. Solo 2013, 37, 177–187. [Google Scholar] [CrossRef]
- Fung, K.F.; Carr, H.P.; Zhang, J.; Wong, M.H. Growth and nutrient uptake of tea under different aluminium concentrations. J. Sci. Food Agric. 2008, 88, 1582–1591. [Google Scholar] [CrossRef]
- Simon, L.; Smalley, T.J.; Benton Jones, J.; Lasseigne, F.T. Aluminum toxicity in tomato. Part 1. growth and mineral nutrition. J. Plant Nutr. 1994, 17, 293–306. [Google Scholar] [CrossRef]
- Zhang, H.; Jiang, Z.; Qin, R.; Zhang, H.; Zou, J.; Jiang, W.; Liu, D. Accumulation and cellular toxicity of aluminum in seedling of Pinus massoniana. BMC Plant Biol. 2014, 14, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delhaize, E.; Ryan, P.R. Aluminum toxicity and tolerance in plants. Plant Physiol. 1995, 107, 315–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, D.L.; Blancaflor, E.B.; Kochian, L.V.; Gilroy, S. Spatial coordination of aluminium uptake, production of reactive oxygen species, callose production and wall rigidification in maize roots. Plant Cell Environ. 2006, 29, 1309–1318. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Liu, P.; Yang, Y.S.; Xu, G.D. Effect of Al in soil on photosynthesis and related morphological and physiological characteristics of two soybean genotypes. Bot. Stud. 2007, 48, 435–444. [Google Scholar]
- Bose, J.; Babourina, O.; Rengel, Z. Role of magnesium in alleviation of aluminium toxicity in plants. J. Exp. Bot. 2011, 62, 2251–2264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panda, S.K.; Baluska, F.; Matsumoto, H. Aluminum stress signaling in plants. Plant Signal. Behav. 2009, 4, 592–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bose, J.; Babourina, O.; Shabala, S.; Rengel, Z. Aluminum-dependent dynamics of ion transport in Arabidopsis: Specificity of low pH and aluminum responses. Physiol. Plant. 2010, 139, 401–412. [Google Scholar] [CrossRef] [PubMed]
- Brunner, I.; Sperisen, C. Aluminum exclusion and aluminum tolerance in woody plants. Front. Plant Sci. 2013, 4, 172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imadi, S.R.; Waseem, S.; Kazi, A.G.; Azooz, M.M.; Ahmad, P. Aluminum toxicity in plants: An overview. In Plant Metal Interaction; Ahmad, P., Ed.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 1–20. [Google Scholar]
- Bryant, K.N.; Fowlkes, A.J.; Mustafa, S.F.; O’neil, B.J.; Osterman, A.C.; Smith, T.M.; Shepard, M.R.; Woosley, R.S.; Butcher, D.J. Determination of aluminum, calcium, and magnesium in fraser fir, balsam fir, and red spruce foliage and soil from the southern and middle appalachians. Microchem. J. 1997, 56, 382–392. [Google Scholar] [CrossRef]
- Oulehle, F.; Hruska, J. Tree species (Picea abies and Fagus sylvatica) effects on soil water acidification and aluminium chemistry at sites subjected to long-term acidification in the Ore Mts., Czech Republic. J. Inorg. Biochem. 2005, 99, 1822–1829. [Google Scholar] [CrossRef] [PubMed]
- Bintz, W.W.; Butcher, D.J. Characterization of the health of southern Appalachian red spruce (Piceae rubens) through determination of calcium, magnesium, and aluminum concentrations in foliage and soil. Microchem. J. 2007, 87, 170–174. [Google Scholar] [CrossRef]
- Perry, C.H.; Amacher, M.C. Patterns of soil calcium and aluminum across the conterminous United States. In Forest Health Monitoring: 2008 National Technical Report. GTR-SRS-158; Potter, K.M., Conkling, B.L., Eds.; US Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2012; pp. 119–130. [Google Scholar]
- Keltjens, W.G.; van Loenen, E. Effects of aluminium and mineral nutrition on growth and chemical composition of hydroponically grown seedlings of five different forest tree species. Plant Soil 1989, 119, 39–50. [Google Scholar] [CrossRef]
- Keltjens, W.G. Effects of aluminum on growth and nutrient status of Douglas-fir seedlings grown in culture solution. Tree Physiol. 1990, 6, 165–175. [Google Scholar] [CrossRef]
- Hirano, Y.; Frey, B.; Brunner, I. Contrasting reactions of roots of two coniferous tree species to aluminum stress. Environ. Exp. Bot. 2012, 77, 12–18. [Google Scholar] [CrossRef]
- Ryder, M.; Gérard, F.; Evans, D.E.; Hodson, M.J. The use of root growth and modelling data to investigate amelioration of aluminium toxicity by silicon in Picea abies seedlings. J. Inorg. Biochem. 2003, 97, 52–58. [Google Scholar] [CrossRef]
- Prabagar, S.; Hodson, M.J.; Evans, D.E. Silicon amelioration of aluminium toxicity and cell death in suspension cultures of Norway spruce (Picea abies (L.) Karst.). Environ. Exp. Bot. 2011, 70, 266–276. [Google Scholar] [CrossRef]
- Raynal, D.J.; Joslin, J.D.; Thornton, F.C.; Schaedle, M.; Henderson, G.S. Sensitivity of tree seedlings to aluminum: iii. red spruce and loblolly pine. J. Environ. Qual. 1990, 19, 180–187. [Google Scholar] [CrossRef]
- Lelu-Walter, M.A.; Gautier, F.; Eliášová, K.; Sanchez, L.; Teyssier, C.; Lomenech, A.M.; Le Metté, C.; Hargreaves, C.; Trontin, J.-F.; Reeves, C.; et al. High gellan gum concentration and secondary somatic embryogenesis: Two key factors to improve somatic embryo development in Pseudotsuga menziesii [Mirb.]. Plant Cell Tissue Organ Cult. 2018, 132, 137–155. [Google Scholar] [CrossRef]
- Zhao, S.J.; Wang, J.H.; Liang, Y.L.; Xu, L.X. Somatic Embryogenesis and plantlet regeneration from hairy roots transformed by agrobacterium rhizogenes in panax quinquefolium L. Int. J. Biomed. Pharm. Sci. 2012, 6, 97–100. [Google Scholar]
- Lelu, M.A.; Bastien, C.; Klimaszewska, K.; Ward, C.; Charest, P.J. An improved method for somatic plantlet production in hybrid larch (Larix x leptoeuropaea): Part 1. Somatic embryo maturation. Plant Cell Tissue Organ Cult. 1994, 36, 107–115. [Google Scholar] [CrossRef]
- Lelu, M.A.; Bastien, C.; Klimaszewska, K.; Charest, P.J. An improved method for somatic plantlet production in hybrid larch (Larix x leptoeuropaea): Part 2. Control of germination and plantlet development. Plant Cell Tissue Organ Cult. 1994, 36, 117–127. [Google Scholar] [CrossRef]
- Tirajoh, A.; Kyung, T.S.; Punja, Z.K. Somatic embryogenesis and plantlet regeneration in American ginseng (Panax quinquefolium L.). In Vitro. Cell. Dev. Bio. Plant 1998, 34, 203–211. [Google Scholar] [CrossRef]
- Eticha, D.; Stass, A.; Horst, W.J. Cell-wall pectin and its degree of methylation in the maize root-apex: Significance for genotypic differences in aluminium resistance. Plant Cell Environ. 2005, 28, 1410–1420. [Google Scholar] [CrossRef]
- Zhou, G.F.; Delhaize, E.; Zhou, M.X.; Ryan, P.R. Biotechnological solutions for enhancing the aluminium resistance of crop plants. In Abiotic Stress in Plants – Mechanisms and Adaptations; Shanker, A., Venkateswarlu, B., Eds.; IntechOpen: London, UK, 2011; pp. 119–142. [Google Scholar] [CrossRef] [Green Version]
- DeWald, L.E.; Sucoff, E.I.; Ohno, T.O.; Buschena, C.A. Response of northern red oak (Quercus rubra) seedlings to soil solution aluminum. Can. J. For. Res. 1990, 20, 331–336. [Google Scholar] [CrossRef]
- Arroyave, C.; Barceló, J.; Poschenrieder, C.; Tolrà, R. Aluminium-induced changes in root epidermal cell patterning, a distinctive feature of hyperresistance to Al in Brachiaria decumbens. J. Inorg. Biochem. 2011, 1477–1483. [Google Scholar] [CrossRef]
- Wagatsuma, T.; Kaneko, M.; Hayasaka, Y. Destruction process of plant root cells by aluminum. Soil Sci. Plant Nutr. 1987, 33, 161–175. [Google Scholar] [CrossRef] [Green Version]
- Souza, L.T.D.; Cambraia, J.; Ribeiro, C.; de Oliveira, J.A.; da Silva, L.C. Effects of aluminum on the elongation and external morphology of root tips in two maize genotypes. Bragantia 2016, 75, 19–25. [Google Scholar] [CrossRef]
- Blamey, F.P.C.; Nishizawa, N.K.; Yoshimura, E. Timing, magnitude, and location of initial soluble aluminum injuries to mungbean roots. Soil Sci. Plant Nutr. 2004, 50, 67–76. [Google Scholar] [CrossRef]
- Kopittke, P.M.; Blamey, F.P.C.; Menzies, N.W. Toxicities of soluble Al, Cu, and La include ruptures to rhizodermal and root cortical cells of cowpea. Plant Soil 2008, 303, 217–227. [Google Scholar] [CrossRef]
- Motoda, H.; Kano, Y.; Hiragami, F.; Kawamura, K.; Matsumoto, H. Morphological changes in the apex of pea roots during and after recovery from aluminium treatment. Plant Soil 2010, 333, 49–58. [Google Scholar] [CrossRef]
- Lăpădătescu, S.; Petolescu, C.; Lazăr, A.; Bălă, M. Influence of growth regulators on morphogenetic processes under in vitro condition. J. Hortic. For. Biotechol. 2012, 16, 197–202. [Google Scholar]
- Lebedev, V.; Arkaev, M.; Dremova, M.; Pozdniakov, I.; Shestibratov, K. Effects ofgrowth regulators and gelling agents on ex vitro rooting of raspberry. Plants 2018, 8, 3. [Google Scholar] [CrossRef] [Green Version]
- van den Dries, N.; Giannì, S.; Czerednik, A.; Krens, F.A.; de Klerk, G.J.M. Flooding of the apoplast is a key factor in the development of hyperhydricity. J. Exp. Bot. 2013, 64, 5221–5230. [Google Scholar] [CrossRef]
- Kevers, C.; Franck, T.; Strasser, R.J.; Dommes, J.; Gaspar, T. Hyperhydricity of micropropagated shoots: A typically stress-induced change of physiological state. Plant Cell Tissue Organ Cult. 2004, 77, 181–191. [Google Scholar] [CrossRef]
- Dantas, A.K.; Majada, J.P.; Fernández, B.; Cañal, M.J. Mineral nutrition in carnation tissue cultures under different ventilation conditions. Plant Growth Regul. 2001, 33, 237–243. [Google Scholar] [CrossRef]
- Huang, J.W.; Grunes, D.L.; Kochian, L.V. Aluminum effects on the kinetics of calcium uptake into cells of the wheat root apex. Planta 1992, 188, 414–421. [Google Scholar] [CrossRef] [PubMed]
- Pongrac, P.; Tolrà, R.; Hajiboland, R.; Vogel-Mikuš, K.; Kelemen, M.; Vavpetič, P.; Pelicon, P.; Barceló, J.; Regvar, M.; Poschenrieder, C.; et al. Contrasting allocation of magnesium, calcium and manganese in leaves of tea (Camellia sinensis (L.) Kuntze) plants may explain their different extraction efficiency into tea. Food Chem. Toxicol. 2020, 135, 110974. [Google Scholar] [CrossRef] [PubMed]
- van Praag, H.J.; Weissen, F.; Dreze, P.; Cogneau, M. Effects of aluminium on calcium and magnesium uptake and translocation by root segments of whole seedlings of Norway spruce (Picea abies Karst). Plant Soil 1997, 189, 267–273. [Google Scholar] [CrossRef]
- Ribeiro, M.A.Q.; de Almeida, A.A.F.; Mielke, M.S.; Gomes, F.P.; Pires, M.V.; Baligar, V.C. Aluminum effects on growth, photosynthesis, and mineral nutrition of cacao genotypes. J. Plant Nutr. 2013, 36, 1161–1179. [Google Scholar] [CrossRef]
- Abdalla, M.M. Physiological aspects of aluminium toxicity on some metabolic and hormonal contents of Hordeum Vulgare seedlings. Aust. J. Basic Appl. Sci. 2008, 2, 549–560. [Google Scholar]
- Maathuis, F.J.M. Sodium in plants: Perception, signalling, and regulation of sodium fluxes. J. Exp. Bot. 2014, 65, 849–858. [Google Scholar] [CrossRef]
- Singha, S.; Townsend, E.C.; Oberly, G.H. Relationship between calcium and agar on vitrification and shoot-tip necrosis of quince (Cydonia oblonga Mill.) shoots in vitro. Plant Cell Tissue Organ Cult. 1990, 23, 135–142. [Google Scholar] [CrossRef]
- Pasqualetto, P.L.; Zimmerman, R.H.; Fordham, I. The influence of cation and gelling agent concentrations on vitrification of apple cultivars in vitro. Plant Cell Tissue Organ Cult. 1988, 14, 31–40. [Google Scholar] [CrossRef]
- Ryan, P.R.; Delhaize, E. The convergent evolution of aluminium resistance in plants exploits a convenient currency. Funct. Plant Biol. 2010, 37, 275–284. [Google Scholar] [CrossRef] [Green Version]
- Cuenca, G.; Herrera, R.; Medina, E.E. Aluminium tolerance in trees of a tropical cloud forest. Plant Soil 1990, 175, 169–175. [Google Scholar] [CrossRef]
- Kariya, K.; Demiral, T.; Sasaki, T.; Tsuchiya, Y.; Turkan, I.; Sano, T.; Hasezawa, S.; Yamamoto, Y. A novel mechanism of aluminum-induced cell death involving vacuolar processing enzyme and vacuolar collapse in tobacco cell line BY-2. J. Inorg. Biochem. 2013, 128, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Dong, Y.; Wang, H.; Chang, E.; Zhao, Z.; Wang, R.; Xu, R.; Jiang, J. Alleviation of aluminum phytotoxicity by canola straw biochars varied with their cultivating soils through an investigation of wheat seedling root elongation. Chemosphere 2019, 218, 907–914. [Google Scholar] [CrossRef] [PubMed]
- Reeves, C.; Hargreaves, C.; Trontin, J.F.; Lelu-Walter, M.A. Simple and efficient protocols for the initiation and proliferation of embryogenic tissue of Douglas-fir. Trees 2018, 32, 175–190. [Google Scholar] [CrossRef] [Green Version]
- Eticha, D.; Staß, A.; Horst, W.J. Localization of aluminium in the maize root apex: Can morin detect cell wall-bound aluminium? J. Exp. Bot. 2005, 56, 1351–1357. [Google Scholar] [CrossRef] [PubMed]
- Astier, C.; Gloaguen, V.; Faugeron, C. Phytoremediation of cadmium-contaminated soils by young Douglas fir trees: Effects of cadmium exposure on cell wall composition. Int. J. Phytoremediation 2014, 16, 790–803. [Google Scholar] [CrossRef]










© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amara, H.; Lelu-Walter, M.-A.; Gloaguen, V.; Faugeron-Girard, C. Tolerance of Douglas Fir Somatic Plantlets to Aluminum Stress: Biological, Cytological, and Mineral Studies. Plants 2020, 9, 536. https://doi.org/10.3390/plants9040536
Amara H, Lelu-Walter M-A, Gloaguen V, Faugeron-Girard C. Tolerance of Douglas Fir Somatic Plantlets to Aluminum Stress: Biological, Cytological, and Mineral Studies. Plants. 2020; 9(4):536. https://doi.org/10.3390/plants9040536
Chicago/Turabian StyleAmara, Holm, Marie-Anne Lelu-Walter, Vincent Gloaguen, and Céline Faugeron-Girard. 2020. "Tolerance of Douglas Fir Somatic Plantlets to Aluminum Stress: Biological, Cytological, and Mineral Studies" Plants 9, no. 4: 536. https://doi.org/10.3390/plants9040536