Management of Verticillium Wilt of Avocado Using Tolerant Rootstocks

 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site
2.2. Orchard Inoculation
2.3. Plant Material
2.4. VW Symptoms Evaluation
2.5. Verticillium Dahliae Isolation and Identification
2.6. Productivity Assessment
2.7. Data Analysis
3. Results and Discussion
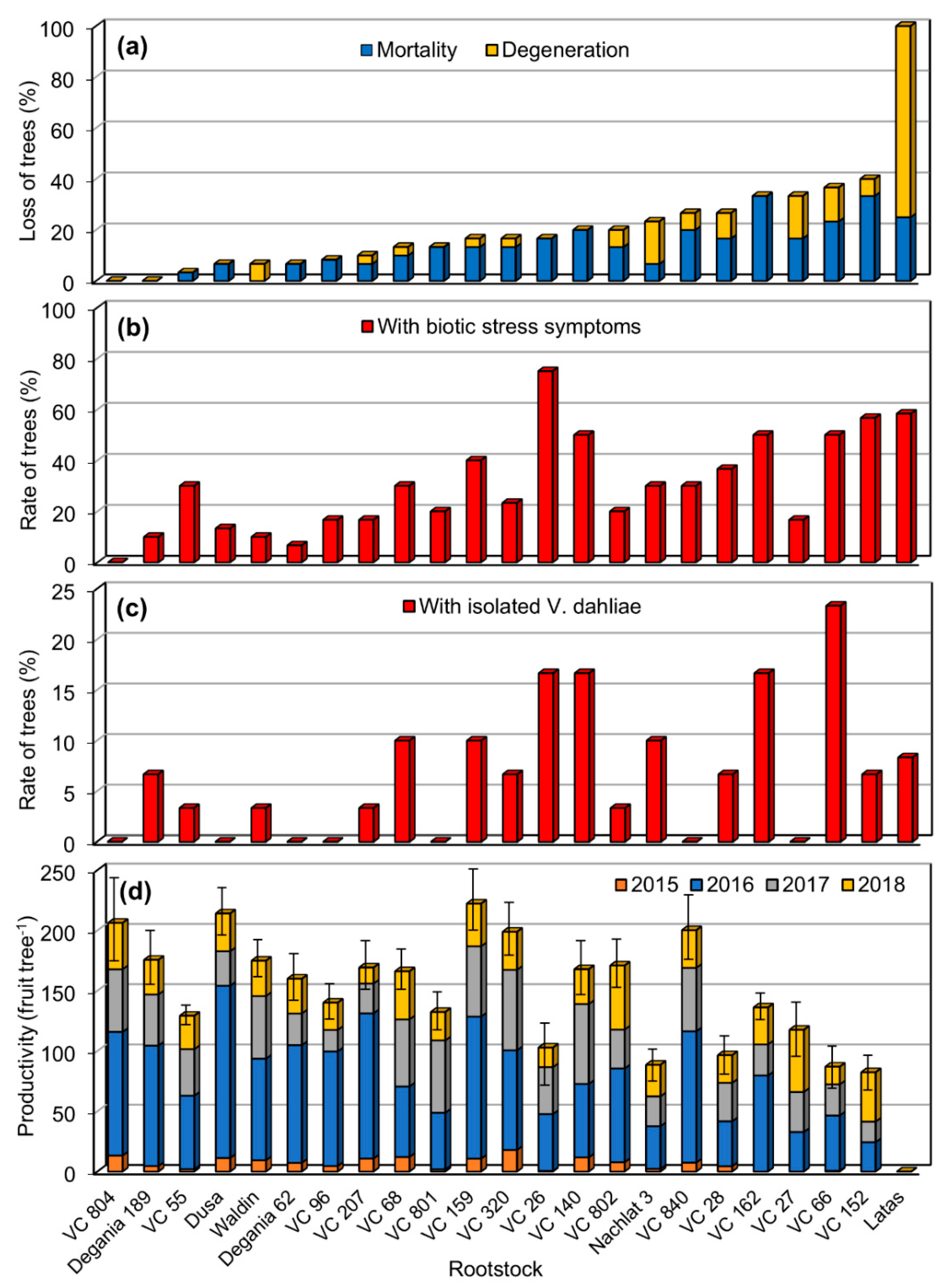
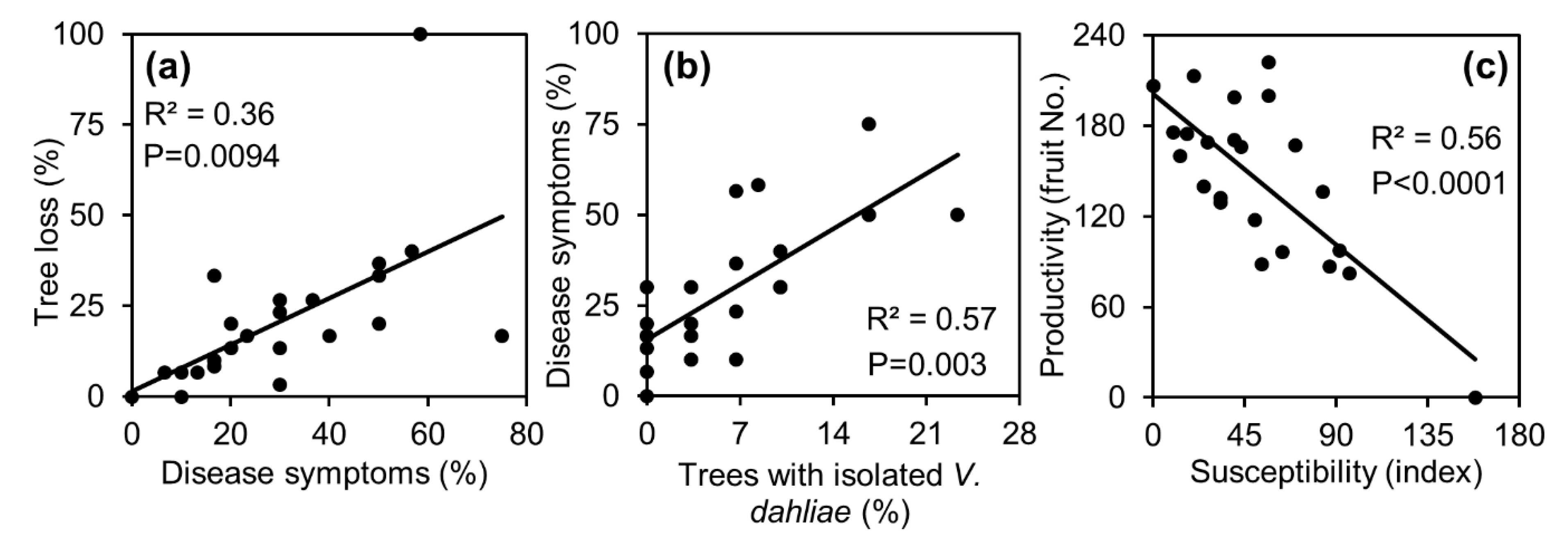
3.1. Loss of Trees
3.2. Verticillium Wilt
3.3. Productivity
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Agrios, G. Vascular wilts caused by ascomycetes and deuteromycetes (mitosporic fungi). In Plant Pathology, 5th ed.; Elsevier Academic Press: New York, NY, USA, 2005; pp. 522–530. [Google Scholar]
- Isaac, I.; Harrison, J.A.C. The symptoms and causal agents of early-dying disease (Verticillium wilt) of potatoes. Ann. Appl. Biol. 1969, 61, 231–244. [Google Scholar] [CrossRef]
- Thomson, R.; McLeod, A.G. Verticillium wilt of tobacco: I. A new disease of tobacco caused by Verticillium dahliae Kleb. N. Z. J. Agric. Res. 1959, 2, 785–791. [Google Scholar] [CrossRef] [Green Version]
- El-Zik, K.M. Integrated control of Verticillium wilt of cotton. Plant Dis. 1985, 69, 1025–1032. [Google Scholar] [CrossRef]
- Levin, A.G.; Erlich, O.; Lebiush, S.; Hazanovsky, M.; Tsror, L. First report of Verticillium wilt caused by Verticillium dahliae on mango in Israel. New Dis. Rep. 2014, 29, 380. [Google Scholar] [CrossRef] [Green Version]
- Tsror, L. Review: Epidemiology and control of Verticillium wilt on olive. Isr. J. Plant Sci. 2011, 59, 59–69. [Google Scholar]
- Schnathorst, W.C. Life cycle and epidemiology of Verticillium. In Fungal wilt Diseases of Plants; Academic Press: New York, NY, USA, 1981; Volume 82. [Google Scholar]
- Wilhelm, S. Longevity of the Verticillium wilt fungus in the laboratory and field. Phytopathology 1955, 45, 180–181. [Google Scholar]
- Markakis, E.A.; Ligoxigakis, E.K.; Avramidou, E.V.; Tzanidakis, N. Survival, persistence, and infection efficiency of Verticillium dahliae passed through the digestive system of sheep. Plant Dis. 2014, 98, 1235–1240. [Google Scholar] [CrossRef] [Green Version]
- Keykhasaber, M.; Thomma, B.P.H.J.; Hiemstra, J.A. Verticillium wilt caused by Verticillium dahliae in woody plants with emphasis on olive and shade trees. Eur. J. Plant Pathol. 2018, 150, 21–37. [Google Scholar] [CrossRef] [Green Version]
- Prieto, P.; Navarro-Raya, C.; Valverde-Corredor, A.; Amyotte, S.G.; Dobinson, K.F.; Mercado-Blanco, J. Colonization process of olive tissues by Verticillium dahliae and its in planta interaction with the biocontrol root endophyte Pseudomonas fluorescens PICF7. Microb. Biotechnol. 2009, 2, 499–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fradin, E.F.; Thomma, B.P.H.J. Physiology and molecular aspects of Verticillium wilt diseases caused by V. dahliae and V. albo-atrum. Mol. Plant Pathol. 2006, 7, 71–86. [Google Scholar] [CrossRef]
- Hiemstra, J.A.; Harris, D.C. A Compendium of Verticillium wilts in Tree Species; Ponsen and Looijen: Wageningen, The Netherlands, 1998; ISBN 9073771250. [Google Scholar]
- Klosterman, S.J.; Atallah, Z.K.; Vallad, G.E.; Subbarao, K.V. Diversity, pathogenicity, and management of Verticillium species. Annu. Rev. Phytopathol. 2009, 47, 39–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zentmyer, G.A. Verticillium wilt of avocado. Phytopathology 1949, 39, 677–682. [Google Scholar]
- Dann, E.K.; Ploetz, R.C.; Coates, L.M.; Pegg, K.G. Foliar, fruit and soilborne diseases. In The Avocado: Botany, Production and Uses; Schaffer, B., Wolstenholme, B.N., Whiley, A.W., Eds.; CABI: Wallingford, UK, 2013; pp. 380–422. [Google Scholar]
- Malcolm, G.M.; Kuldau, G.A.; Gugino, B.K.; Jiménez-Gasco, M.D.M. Hidden host plant associations of soilborne fungal pathogens: An ecological perspective. Phytopathology 2013, 103, 538–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karajeh, M.R.; Masoud, S.A. Molecular detection of Verticillium dahliae Kleb. in asymptomatic olive trees. J. Phytopathol. 2006, 154, 496–499. [Google Scholar] [CrossRef]
- Levin, A.G.; Lavee, S.; Tsror, L. Epidemiology and effects of verticillium wilt on yield of olive trees (cvs. Barnea and Souri) irrigated with saline water in Israel. Phytoparasitica 2003, 31, 333–343. [Google Scholar] [CrossRef]
- Levin, A.G.; Lavee, S.; Tsror, L. Epidemiology of Verticillium dahliae on olive (cv. Picual) and its effect on yield under saline conditions. Plant Pathol. 2003, 52, 212–218. [Google Scholar] [CrossRef]
- Bost, J.B.; Smith, N.J.H.; Crane, J.H. History, distribution and uses. In The Avocado: Botany, Production and Uses; Schaffer, B., Wolstenholme, B.N., Whiley, A.W., Eds.; CABI: Wallingford, UK, 2013; pp. 10–30. [Google Scholar]
- Ben-Ya’acov, A.; Michelson, E. Avocado rootstocks. Hortic. Rev. (Am. Soc. Hortic. Sci). 1995, 17, 381–429. [Google Scholar]
- Crane, J.H.; Douhan, G.; Faber, B.A.; Arpaia, M.L.; Bender, G.S.; Balerdi, C.F.; Barrientos-Priego, A.F. Cultivars and rootstocks. In The avocado: Botany, Production and Uses, 2nd ed.; Schaffer, B., Wolstenholme, B.N., Whiley, A.W., Eds.; CABI: Boston, MA, USA, 2013; pp. 200–233. [Google Scholar]
- Ambrozek, C.; Saitone, T.L.; Sexton, R.J. Five-Year Evaluation of the Hass Avocado Board’s Promotion Programs: 2013–2017; Hass Avocado Board: Mission Viejo, CA, USA, 2018. [Google Scholar]
- Stout, A.B.; Savage, E.M. The flower behavior of avocados with special reference to interplanting. Proc. Fla. State Hort. Soc. 1925, 38, 80–91. [Google Scholar]
- Zentmyer, G.A. Soil-borne pathogens of avocado. In Proceedings of the First International Tropical Fruit Short Course: The Avocado; University of Florida: Gainesville, FL, USA, 1976. [Google Scholar]
- Smith, L.A.; Dann, E.K.; Pegg, K.G.; Whiley, A.W.; Giblin, F.R.; Doogan, V.; Kopittke, R. Field assessment of avocado rootstock selections for resistance to phytophthora root rot. Aust. Plant Pathol. 2011, 40, 39–47. [Google Scholar] [CrossRef]
- COFFEY, M. Phytophthora root rot of avocado: An integrated approach to control in California. Plant Dis. 1987, 71, 1046–1052. [Google Scholar]
- Douhan, G.W. Current and future perspectives regarding avocado rootstock breeding at UCR. Calif. Avocado Soc. Yearb. 2009, 92, 87–111. [Google Scholar]
- Schieber, G.; Zentmyer, E. The search for resistance to Phytophthora root rot in Latin America. S. Afr. Avocado Growers Assoc. Yearb. 1987, 10, 109–110. [Google Scholar]
- Tsror, L.; Nachmias, A. Significance of the root system in Verticillium wilt tolerance in potato and resistance in tomato. Isr. J. Plant Sci. 1995, 43, 315–323. [Google Scholar]
- Paplomatas, E.J.; Elena, K.; Tsagkarakou, A.; Perdikaris, A. Control of verticillium wilt of tomato and cucurbits through grafting of commercial varieties on resistant rootstocks. Acta Hortic. 2002, 579, 445–449. [Google Scholar] [CrossRef]
- Bletsos, F.; Thanassoulopoulos, C.; Roupakias, D. Effect of grafting on growth, yield, and verticillium wilt of eggplant. HortScience 2003, 38, 183–186. [Google Scholar] [CrossRef]
- Temperini, O.; Calabrese, N.; Temperini, A.; Rouphael, Y.; Tesi, R.; Lenzi, A.; Carito, A.; Colla, G. Grafting artichoke onto cardoon rootstocks: Graft compatibility, yield and Verticillium wilt incidence. Sci. Hortic. (Amsterdam) 2013, 149, 22–27. [Google Scholar] [CrossRef]
- López-Escudero, F.J.; Mercado-Blanco, J. Verticillium wilt of olive: A case study to implement an integrated strategy to control a soil-borne pathogen. Plant Soil 2011, 344, 1–50. [Google Scholar] [CrossRef] [Green Version]
- Bubici, G.; Cirulli, M. Control of Verticillium wilt of olive by resistant rootstocks. Plant Soil 2012, 352, 363–376. [Google Scholar] [CrossRef]
- Lahav, E.; Lavi, U. Genetics and breeding. In The Avocado: Botany, Production and Uses; Schaffer, B., Wolstenholme, B.N., Whiley, A.W., Eds.; CABI: Oxfordshire, UK, 2013; pp. 51–85. [Google Scholar]
- Ben-Ya’acov, A.; Frenkel, M. Sensitivity of avocado rootstocks to Verticillium wilt. Calif. Avocado Soc. Yearb. 1974, 57, 111–113. [Google Scholar]
- Halma, F.F.; Zentmyer, G.A.; Wilhelm, S. Susceptibility of avocado rootstocks to Verticillium wilt. Calif. Avocado Soc. Yearb. 1954, 38, 153–155. [Google Scholar]
- Lazare, S.; Haberman, A.; Yermiyahu, U.; Erel, R.; Simenski, E.; Dag, A. Avocado rootstock influences scion leaf mineral content. Arch. Agron. Soil Sci. 2019, 1–11. [Google Scholar] [CrossRef]
- Warton, D.I.; Hui, F.K.C. The arcsine is asinine: The analysis of proportions in ecology. Ecology 2011, 92, 3–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levin, A.G.; Lavee, S.; Tsror, L. The influence of salinity on Verticillium dahliae in stem cuttings of five olive cultivars. J. Phytopathol. 2007, 155, 587–592. [Google Scholar] [CrossRef]
- Saadatmand, A.R.; Banihashemi, Z.; Sepaskhah, A.R.; Maftoun, M. Soil salinity and water stress and their effect on susceptibility to verticillium wilt disease, ion composition and growth of pistachio. J. Phytopathol. 2008, 156, 287–292. [Google Scholar] [CrossRef]
- Hillel, D. Salinity Management for Sustainable Irrigation: Integrating Science, Environment, and Economics; The World Bank: Wmhington, DC, USA, 2000; ISBN 082134773X. [Google Scholar]
- Salazar-García, S.; Garner, L.C.; Lovatt, C.J. Reproductive biology. In The Avocado, Botany, Production and Uses, 2nd ed.; Schaffer, B., Wolstenholme, B.N., Whiley, A.W., Eds.; CABI: Oxfordshire, UK, 2013; pp. 118–167. [Google Scholar]
- Robb, J. Verticillium tolerance: Resistance, susceptibility, or mutualism? Can. J. Bot. 2007, 85, 903–910. [Google Scholar] [CrossRef]
- Norris, R.F. Integrated pest management. In Encyclopedia of Biological Invasions; Simberloff, D., Rejmanek, M., Eds.; University of California Press: Berkeley, CA, USA, 2011. [Google Scholar]
- Webber, H.J. The avocado stock problem. In Annual Reports 1925 and 1926; California Avocado Association: Irvine, CA, USA, 1926; pp. 37–38. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Rootstock | Race (Putative) a | Propagation Method | Origin |
|---|---|---|---|
| VC26 | W.I. | Clonal | A. Ben-Ya’acov collection; Israel |
| VC27 | W.I. | Clonal | A. Ben-Ya’acov collection; Israel |
| VC28 | W.I. | Clonal | A. Ben-Ya’acov collection; Israel |
| VC55 | W.I. | Clonal | A. Ben-Ya’acov collection; Israel |
| VC66 | W.I. | Clonal | A. Ben-Ya’acov collection; Israel |
| VC68 | W.I. | Clonal | A. Ben-Ya’acov collection; Israel |
| VC96 | W.I. | Clonal | A. Ben-Ya’acov collection; Israel |
| VC140 | W.I. | Clonal | A. Ben-Ya’acov collection; Israel |
| VC152 | W.I. | Clonal | A. Ben-Ya’acov collection; Israel |
| VC159 | W.I. | Clonal | A. Ben-Ya’acov collection; Israel |
| VC162 | W.I. | Clonal | A. Ben-Ya’acov collection; Israel |
| VC207 | W.I.×Mex. | Clonal | Day; U.S. |
| VC320 | W.I. | Clonal | Kaiima Bio Agritech; Israel |
| VC801 | W.I. | Clonal | A. Ben-Ya’acov collection; Israel |
| VC802 | W.I. | Clonal | A. Ben-Ya’acov collection; Israel |
| VC804 | W.I. | Clonal | A. Ben-Ya’acov collection; Israel |
| VC840 | Mex. | Clonal | A. Ben-Ya’acov collection; Israel |
| Latas | Mex.×Gu. | Clonal | Westfalia Fruit; South Africa |
| Dusa | Mex.×Gu. | Clonal | Westfalia Fruit; South Africa |
| Waldin | W.I. | Seed | U.S. |
| Degania 62 | W.I. | Seed | A. Ben-Ya’acov collection; Israel |
| Degania 189 | W.I. | Seed | A. Ben-Ya’acov collection; Israel |
| Nachlat 3 | W.I. | Seed | A. Ben-Ya’acov collection; Israel |
| Rootstock | Number of Evaluated Trees | Productivity (Fruit Tree−1) a | Susceptibility (Index) b | Performance Score c |
|---|---|---|---|---|
| VC804 | 30 | 206 a | 0 | 206 |
| Dusa | 30 | 213 a | 20 | 193 |
| Degania 189 | 30 | 176 abc | 10 | 166 |
| VC159 | 30 | 222 a | 57 | 165 |
| VC320 | 30 | 199 ab | 40 | 159 |
| Waldin | 30 | 175 abc | 17 | 158 |
| Degania 62 | 30 | 160 abc | 13 | 147 |
| VC840 | 30 | 200 ab | 57 | 143 |
| VC207 | 30 | 169 abc | 27 | 142 |
| VC802 | 30 | 171 abc | 40 | 131 |
| VC68 | 30 | 166 abc | 43 | 123 |
| VC96 | 12 | 140 abc | 25 | 115 |
| VC801 | 30 | 132 abc | 33 | 99 |
| VC140 | 30 | 167 abc | 70 | 97 |
| VC55 | 30 | 129 abc | 33 | 96 |
| VC27 | 18 | 118 abc | 50 | 68 |
| VC162 | 6 | 136 abc | 83 | 53 |
| Nachlat 3 | 30 | 89 c | 53 | 36 |
| VC28 | 30 | 96 bc | 63 | 33 |
| VC26 | 26 | 98 abc | 92 | 6 |
| VC66 | 30 | 87 c | 87 | 0 |
| VC152 | 30 | 82 c | 97 | −15 |
| Latas | 12 | 0 | 158 | −158 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haberman, A.; Tsror, L.; Lazare, S.; Hazanovsky, M.; Lebiush, S.; Zipori, I.; Busatn, A.; Simenski, E.; Dag, A. Management of Verticillium Wilt of Avocado Using Tolerant Rootstocks. Plants 2020, 9, 531. https://doi.org/10.3390/plants9040531
Haberman A, Tsror L, Lazare S, Hazanovsky M, Lebiush S, Zipori I, Busatn A, Simenski E, Dag A. Management of Verticillium Wilt of Avocado Using Tolerant Rootstocks. Plants. 2020; 9(4):531. https://doi.org/10.3390/plants9040531
Chicago/Turabian StyleHaberman, Amnon, Leah Tsror (Lahkim), Silit Lazare, Marina Hazanovsky, Sara Lebiush, Isaac Zipori, Amnon Busatn, Eli Simenski, and Arnon Dag. 2020. "Management of Verticillium Wilt of Avocado Using Tolerant Rootstocks" Plants 9, no. 4: 531. https://doi.org/10.3390/plants9040531