The Spread and Transmission of Sweet Potato Virus Disease (SPVD) and Its Effect on the Gene Expression Profile in Sweet Potato

 ,
,
Abstract
:1. Introduction
2. Results
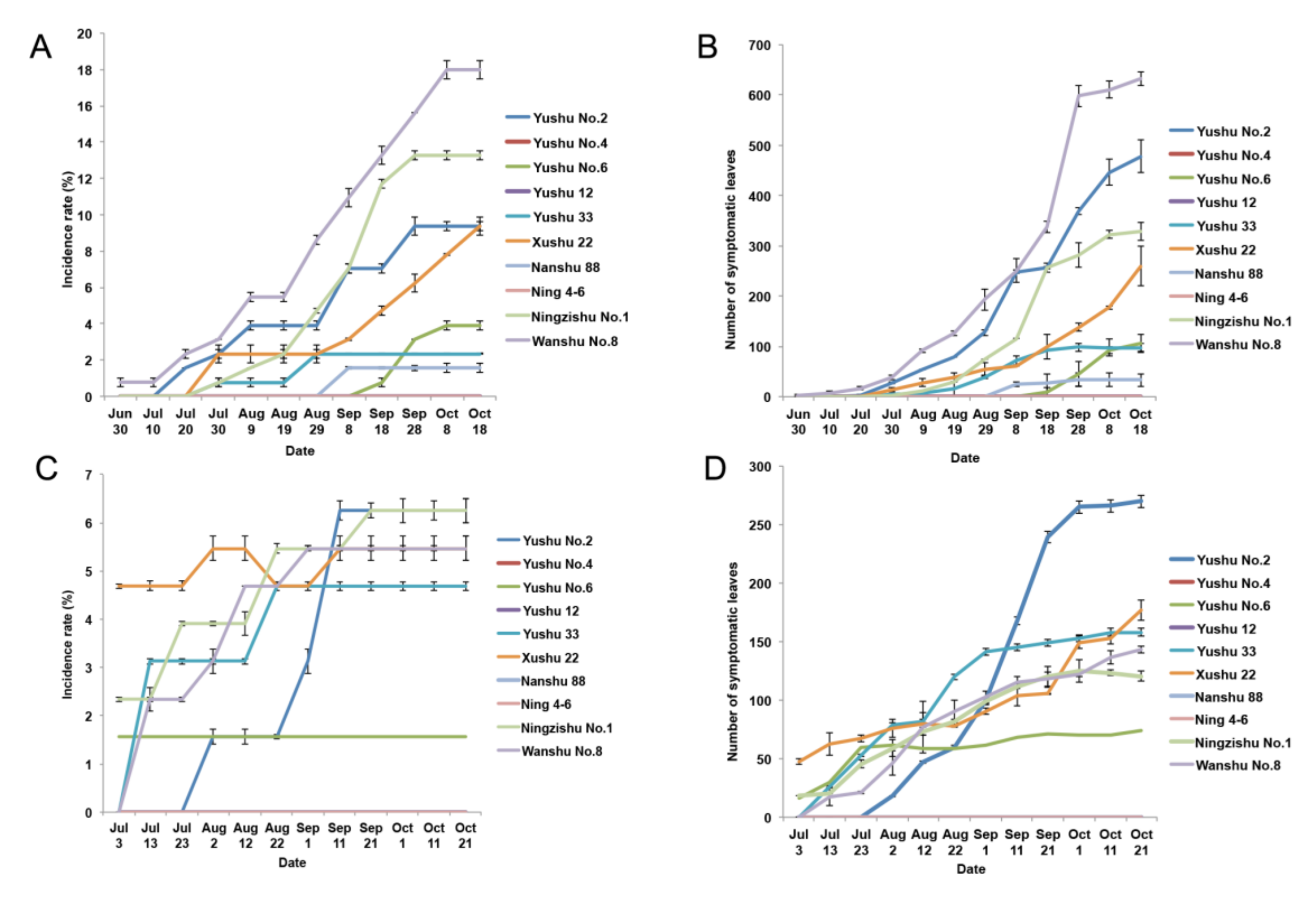
2.1. Characteristics of SPVD Spread in the Field
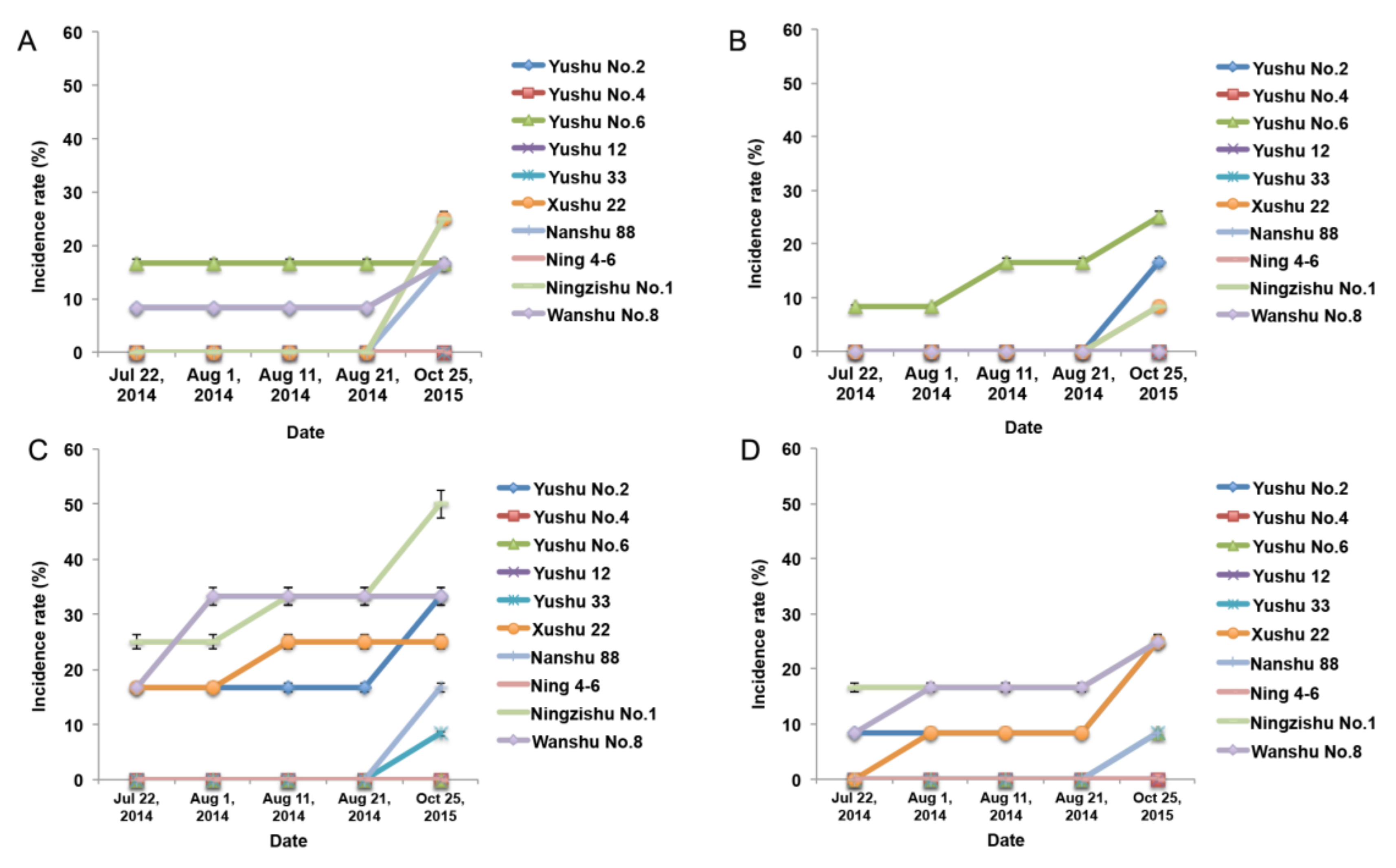
2.2. SPVD Transmission through Artificial Friction Inoculation
2.3. Grafting was Effective for SPVD Transmission
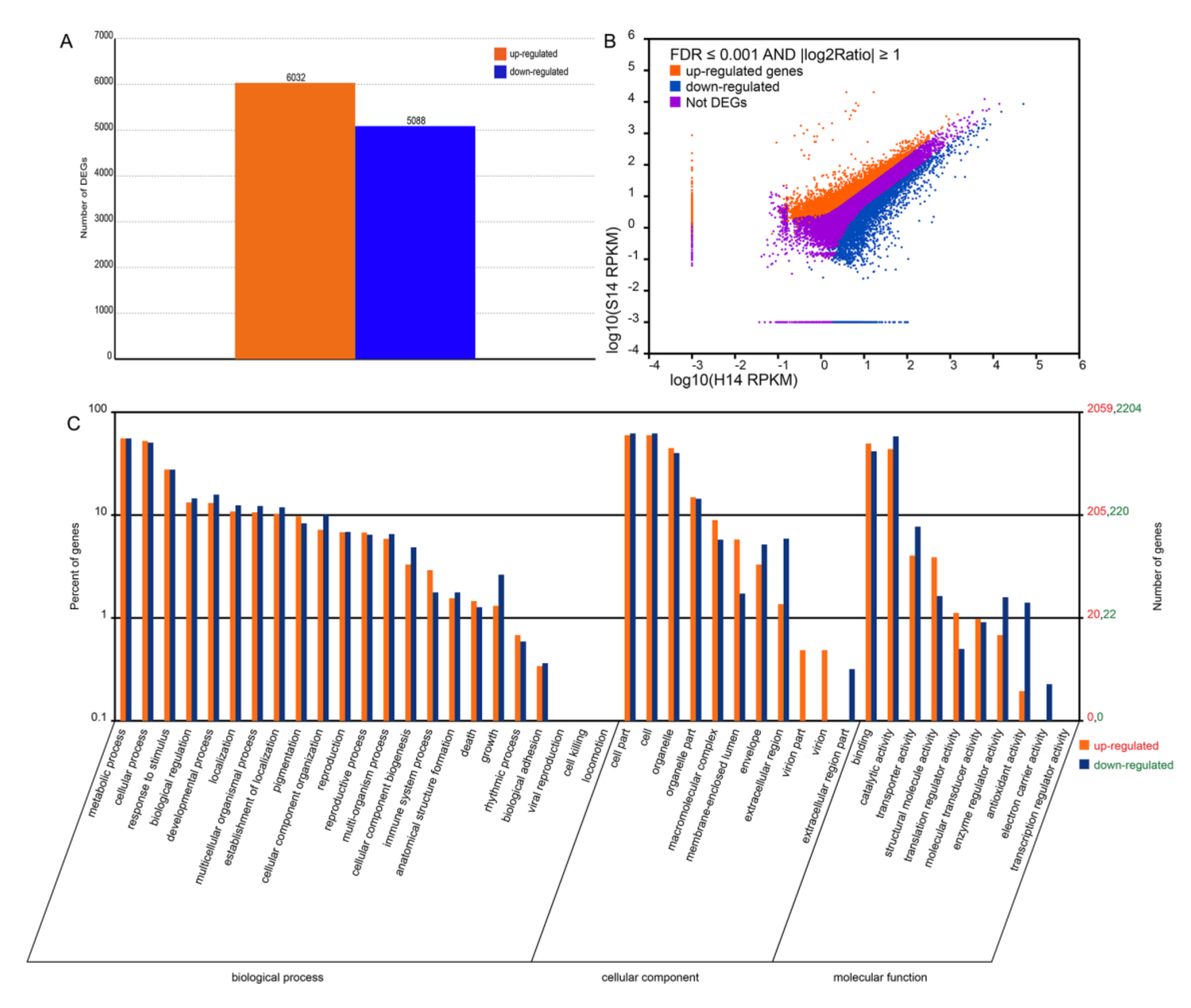
2.4. Comparison of the Transcriptomes of Non-Infected and SPFMV and SPCSV-Infected Sweet Potato
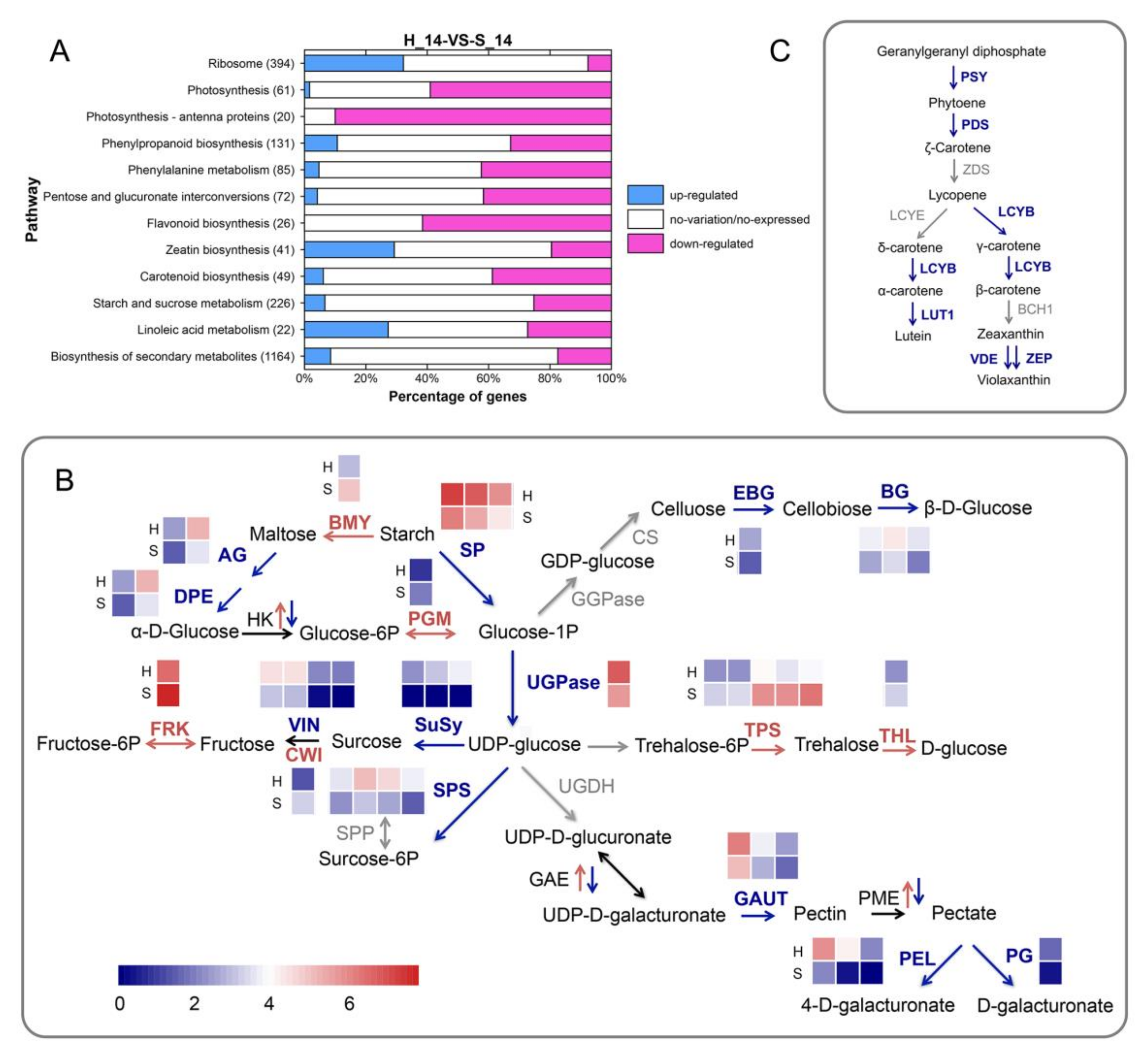
2.5. Gene Ontology and Metabolic Pathway Analysis of Differentially Expressed Genes
2.6. Genes Involved in Photosynthesis are Downregulated in SPFMV and SPCSV-Infected Plants when Compared with Non-Infected Plants
2.7. SPVD Affects the Expression Levels of Genes Involved in Carbohydrate Metabolism
2.8. Expression of Phenylpropanoid Metabolic Pathway Genes is Altered in the Infected Plants
2.9. SPVD Alters the Expression of Genes Involved in Carotenoid, Zeatin, and Linoleic Acid Metabolism
2.10. DEGs that Participate in the Response to SPVD
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Field Experiments
4.3. Friction Inoculation Experiments
4.4. Graft Inoculation
4.5. Plant Materials used for Transcriptome Analysis
4.6. RNA Extraction
4.7. Illumina Sequencing, Data Filtering, and De Novo Assembly
4.8. Unigene Expression Analysis and Differentially Expressed Gene (DEG) Identification
4.9. Functional Annotation
4.10. GO Term and KEGG Pathway Enrichment
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Tigabu, B.; Tilahun, B. Performance evaluation of improved sweet potato (Ipomoea batatas L.) varieties at Gedeo Zone, Southern Ethiopia. Int. J. Sci. Res. 2015, 4, 116–119. [Google Scholar]
- Zhang, K.; Wu, Z.D.; Tang, D.B.; Lv, C.W.; Luo, K.; Zhao, Y.; Liu, X.; Huang, Y.X.; Wang, J.C. Development and identification of SSR markers associated with starch properties and β-carotene content in the storage root of sweet potato (Ipomoea batatas L.). Front. Plant Sci. 2016, 7, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schafleitner, R.; Tincopa, L.R.; Palomino, O.; Rossel, G.; Robles, R.F.; Alagon, R.; Rivera, C.; Quispe, C.; Rojas, L.; Pacheco, J.A.; et al. A sweetpotato gene index established by de novo assembly of pyrosequencing and Sanger sequences and mining for gene-based microsatellite markers. BMC Genom. 2010, 11, 604. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barkessa, M.K.E. A review on sweet potato (Ipomea batatas) viruses and associated diseases. Int. J. Res. Agric. For. 2018, 5, 1–10. [Google Scholar]
- Gibson, R.W.; Aritua, V. The perspective of sweetpotato chlorotic stunt virus in sweetpotato production in Africa: A review. Afr. Crop Sci. J. 2002, 10, 281–310. [Google Scholar] [CrossRef]
- Adikini, S.; Mukasa, S.B.; Mwanga, R.O.M.; Gibson, R.W. Effects of Sweet potato feathery mottle virus and Sweet potato chlorotic stunt virus on the yield of sweetpotato in Uganda. J. Phytopathol. 2016, 164, 242–254. [Google Scholar] [CrossRef]
- Tairo, F.; Mukasa, S.B.; Jones, R.A.C.; Kullaya, A.; Rubaihayo, P.R.; Valkonen, J.P.T. Unravelling the genetic diversity of the three main viruses involved in Sweet Potato Virus Disease (SPVD), and its practical implications. Mol. Plant Pathol. 2005, 6, 199–211. [Google Scholar] [CrossRef]
- Cuellar, W.J.; Kreuze, J.F.; Rajamäki, M.L.; Cruzado, K.R.; Untiveros, M.; Valkonen, J.P.T. Elimination of antiviral defense by viral RNase III. PNAS 2009, 106, 10354–10358. [Google Scholar] [CrossRef] [Green Version]
- Untiveros, M.; Quispe, D.; Kreuze, J. Analysis of complete genomic sequences of isolates of the sweet potato feathery mottle virus strains C and EA: Molecular evidence for two distinct potyvirus species and two P1 protein domains. Arch. Virol. 2010, 155, 2059–2063. [Google Scholar] [CrossRef]
- Kwak, H.R.; Kim, J.; Kim, M.K.; Seo, J.K.; Jung, M.N.; Kim, J.S.; Lee, S.; Choi, H.S. Molecular characterization of five potyviruses infecting Korean sweet potatoes based on analyses of complete genome sequences. Plant Pathol. J. 2015, 31, 388–401. [Google Scholar] [CrossRef] [Green Version]
- Karyeija, R.F.; Kreuze, J.F.; Gibson, R.W.; Valkonen, J.P.T. Synergistic interactions of a Potyvirus and a phloem-limited Crinivirus in sweet potato plants. Virology 2000, 269, 26–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kokkinos, C.D.; Clark, C.A.; McGregor, C.E.; LaBonte, D.R. The effect of sweet potato virus disease and its viral components on gene expression levels in sweetpotato. J. Am. Soc. Hortic. Sci. 2006, 131, 657–666. [Google Scholar] [CrossRef] [Green Version]
- Pipan, B.; Žnidarčič, D.; Kunstelj, N.; Meglič, V. Genetic evaluation of sweetpotato accessions introduced to the Central European area. J. Agric. Sci. Technol. 2017, 19, 1139–1150. [Google Scholar]
- Pipan, B.; Žnidarčič, D.; Meglič, V. Evaluation of genetic diversity of sweet potato [Ipomoea batatas (L.) Lam.] on different ploidy levels applying two capillary platforms. J. Hortic. Sci. Biotechnol. 2017, 92, 192–198. [Google Scholar] [CrossRef]
- Yada, B.; Alajo, A.; Ssemakula, G.N.; Mwanga, R.O.M.; Brown-Guedira, G.; Yencho, G.C. Selection of simple sequence repeat markers associated with inheritance of sweetpotato virus disease resistance in sweetpotato. Crop Sci. 2017, 57, 1421–1430. [Google Scholar] [CrossRef] [Green Version]
- Ngailo, S.; Shimelis, H.; Sibiya, J.; Mtunda, K. Screening of Tanzanian sweet potato germplasm for yield and related traits and resistance to sweet potato virus disease. Acta Agric. Scand. Sec. B Soil Plant Sci. 2016, 66, 52–66. [Google Scholar] [CrossRef]
- Ngailo, S.; Shimelis, H.; Sibiya, J.; Mtunda, K.; Mashilo, J. Genotype-by-environment interaction of newly-developed sweet potato genotypes for storage root yield, yield-related traits and resistance to sweet potato virus disease. Heliyon 2019, 5, e01448. [Google Scholar] [CrossRef] [Green Version]
- Ngailo, S.; Shimelis, H.; Sibiya, J.; Mtunda, K.; Mashilo, J. Combining ability and heterosis of selected sweetpotato (Ipomoea batatas L.) clones for storage root yield, yield-related traits and resistance to sweetpotato virus disease. Euphytica 2019, 215, 87. [Google Scholar] [CrossRef]
- Wang, L.T.; Wang, A.Y.; Hsieh, C.W.; Chen, C.Y.U.; Sung, H.Y. Vacuolar invertases in sweet potato: Molecular cloning, characterization, and analysis of gene expression. J. Agric. Food Chem. 2005, 53, 3672–3678. [Google Scholar] [CrossRef]
- Zhang, K.; Wu, Z.D.; Tang, D.B.; Luo, K.; Lu, H.X.; Liu, Y.Y.; Dong, J.; Wang, X.; Lv, C.W.; Wang, J.C.; et al. Comparative transcriptome analysis reveals critical function of sucrose metabolism related-enzymes in starch accumulation in the storage root of sweet potato. Front. Plant Sci. 2017, 8, 914. [Google Scholar] [CrossRef]
- Cai, L.; Zhang, L.; Fu, Q.; Xu, Z.F. Identification and expression analysis of cytokinin metabolic genes IPTs, CYP735A and CKXs in the biofuel plant Jatropha curcas. PeerJ 2018, 6, e4812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, J.L.; Yan, C.S.; Roston, R.; Shanklin, J.; Xu, C.C. Arabidopsis lipins, PDAT1 ACYLTRANSFERASE, and SDP1 triacylglycerol lipase synergistically direct fatty acids toward β-oxidation, thereby maintaining membrane lipid homeostasis. Plant Cell 2014, 26, 4119–4134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.S.; Li, Z.Y.; Chen, Y.; Chen, M.; Li, L.C.; Ma, Y.Z. Heat shock protein 90 in plants: Molecular mechanisms and roles in stress responses. Int. J. Mol. Sci. 2012, 13, 15706–15723. [Google Scholar] [CrossRef]
- Moshe, A.; Gorovits, R.; Liu, Y.; Czosnek, H. Tomato plant cell death induced by inhibition of HSP90 is alleviated by Tomato yellow leaf curl virus infection. Mol. Plant Pathol. 2016, 17, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Gorovits, R.; Moshe, A.; Amrani, L.; Kleinberger, R.; Anfoka, G.; Czosnek, H. The six Tomato yellow leaf curl virus genes expressed individually in tomato induce different levels of plant stress response attenuation. Cell Stress Chaperones 2017, 22, 345–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, T.J.; Busta, L.; Zhang, Q.; Ding, P.T.; Jetter, R.; Zhang, Y.L. TGACG-BINDING FACTOR 1 (TGA1) and TGA4 regulate salicylic acid and pipecolic acid biosynthesis by modulating the expression of systemic acquired resistance deficient 1 (SARD1) and calmodulin-binding protein 60g (CBP60g). New Phytol. 2018, 217, 344–354. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Liu, S.T.; Qiao, Q.; Zhang, D.S.; Qin, Y.H.; Zhang, Z.C. Methodology for identification of disease resistance of sweet potato cultivars to sweet potato virus disease and yield loss estimation. Acta Phytophyl. Sin. 2014, 41, 176–181. [Google Scholar]
- Gibson, R.W.; Kreuze, J.F. Degeneration in sweetpotato due to viruses, virus-cleaned planting material and reversion: A review. Plant Pathol. 2015, 64, 1–15. [Google Scholar] [CrossRef]
- Rwegasira, G.M. Prevalence of sweetpotato virus vectors in Lake Victoria Basin of Tanzania: A challenge to disease management techniques. J. Agric. Sci. Technol. A 2011, 1, 979–986. [Google Scholar] [CrossRef]
- Wau, W.; Komolong, B. Evaluation of sweet potato virus occurrence and distribution in sweet potato farmer’s fields in Papua New Guinea. Acta Sci. Agric. 2019, 3, 62–67. [Google Scholar]
- Lu, H.X.; Lv, C.W.; Wu, Z.D.; Luo, K.; Yin, W.; Yang, H.; Wang, J.C.; Zhang, K. Development of detection method for sweet potato feathery mottle virus (SPFMV) and sweet potato chlorotic stunt virus (SPCSV) through fluorescence quantitative RT-PCR. Sci. Agric. Sin. 2016, 49, 90–102. [Google Scholar]
- Garcia-Ruiz, H. Susceptibility genes to plant viruses. Viruses 2018, 10, 484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gutierrez, D.L.; Fuentes, S.; Salazar, L.F. Sweetpotato virus disease (SPVD): Distribution, incidence, and effect on sweetpotato yield in Peru. Plant Dis. 2003, 87, 297–302. [Google Scholar] [CrossRef] [PubMed]
- Yan, S.L.; Lehrer, A.T.; Hajirezaei, M.R.; Springer, A.; Komor, E. Modulation of carbohydrate metabolism and chloroplast structure in sugarcane leaves which were infected by Sugarcane Yellow Leaf Virus (SCYLV). Physiol. Mol. Plant Pathol. 2008, 73, 78–87. [Google Scholar] [CrossRef]
- De Haro, L.A.; Arellano, S.M.; Novák, O.; Feil, R.; Dumón, A.D.; Mattio, M.F.; Tarkowská, D.; Llauger, G.; Strnad, M.; Lunn, J.E.; et al. Mal de Río Cuarto virus infection causes hormone imbalance and sugar accumulation in wheat leaves. BMC Plant Biol. 2019, 19, 112. [Google Scholar] [CrossRef] [Green Version]
- Hipper, C.; Brault, V.; Ziegler-Graff, V.; Revers, F. Viral and cellular factors involved in phloem transport of plant viruses. Front. Plant Sci. 2013, 4, 154. [Google Scholar] [CrossRef] [Green Version]
- Malinovsky, F.G.; Fangel, J.U.; Willats, W.G.T. The role of the cell wall in plant immunity. Front. Plant Sci. 2014, 5, 178. [Google Scholar] [CrossRef] [Green Version]
- Bacete, L.; Mélida, H.; Miedes, E.; Molina, A. Plant cell wall-mediated immunity: Cell wall changes trigger disease resistance responses. Plant J. 2018, 93, 614–636. [Google Scholar] [CrossRef]
- Kogovšek, P.; Pompe-Novak, M.; Petek, M.; Fragner, L.; Weckwerth, W.; Gruden, K. Primary metabolism, phenylpropanoids and antioxidant pathways are regulated in potato as a response to potato virus y infection. PLoS ONE 2016, 11, e0146135. [Google Scholar] [CrossRef]
- Shi, J.; Zhao, L.; Yan, B.Y.; Zhu, Y.Q.; Ma, H.S.; Chen, W.Y.; Ruan, S.L. Comparative transcriptome analysis reveals the transcriptional alterations in growth-and development-related genes in sweet potato plants infected and non-infected by SPFMV, SPV2, and SPVG. Int. J. Mol. Sci. 2019, 20, 1012. [Google Scholar] [CrossRef] [Green Version]
- Leiss, K.A.; Choi, Y.H.; Verpoorte, R.; Klinkhamer, P.G.L. An overview of NMR-based metabolomics to identify secondary plant compounds involved in host plant resistance. Phytochem. Rev. 2011, 10, 205–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, C.Z.; Zhang, Y.Y.; Zhong, Y.; Yang, J.W.; Yan, S.T. Gene expression changes in leaves of Citrus sinensis (L.) Osbeck infected by Citrus tristeza virus. J. Hortic. Sci. Biotechnol. 2016, 91, 466–475. [Google Scholar] [CrossRef]
- Hossain, M.Z.; Ishiga, Y.; Yamanaka, N.; Ogiso-Tanaka, E.; Yamaoka, Y. Soybean leaves transcriptomic data dissects the phenylpropanoid pathway genes as a defence response against Phakopsora pachyrhizi. Plant Physiol. Biochem. 2018, 132, 424–433. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ahammed, G.J.; Li, X.; Wei, J.P.; Li, Y.; Yan, P.; Zhang, L.P.; Han, W.Y. Exogenous brassinosteroid enhances plant defense against Colletotrichum gloeosporioides by activating phenylpropanoid pathway in Camellia sinensis L. J. Plant Growth Regul. 2018, 37, 1235–1243. [Google Scholar] [CrossRef]
- Dixon, R.A.; Achnine, L.; Kota, P.; Liu, C.J.; Reddy, M.S.S.; Wang, L.J. The phenylpropanoid pathway and plant defence-A genomics perspective. Mol. Plant Pathol. 2002, 3, 271–390. [Google Scholar] [CrossRef] [PubMed]
- Chezem, W.R.; Memon, A.; Li, F.S.; Weng, J.K.; Clay, N.K. SG2-type R2R3-MYB transcription factor MYB15 controls defense-induced lignification and basal immunity in Arabidopsis. Plant Cell 2017, 29, 1907–1926. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.X.; Lu, S.F. Biosynthesis and regulation of phenylpropanoids in plants. Crit. Rev. Plant Sci. 2017, 36, 257–290. [Google Scholar] [CrossRef]
- Jabeen, A.; Kiran, T.V.; Subrahmanyam, D.; Lakshmi, D.L.; Bhagyanarayana, G.; Krishnaveni, D. Variations in chlorophyll and carotenoid contents in Tungro infected rice plants. J. Res. Dev. 2017, 5, 1–7. [Google Scholar]
- Kapinga, R.; Ndunguru, J.; Mulokozi, G.; Tumwegamire, S. Impact of common sweetpotato viruses on total carotenoids and root yields of an orange-fleshed sweetpotato in Tanzania. Sci. Hortic. Amst. 2009, 122, 1–5. [Google Scholar] [CrossRef]
- Tian, L. Recent advances in understanding carotenoid-derived signaling molecules in regulating plant growth and development. Front. Plant Sci. 2015, 6, 790. [Google Scholar] [CrossRef] [Green Version]
- Park, S.W.; Liu, P.P.; Forouhar, F.; Vlot, A.C.; Tong, L.; Tietjen, K.; Klessig, D.F. Use of a synthetic salicylic acid analog to investigate the roles of methyl salicylate and its esterases in plant disease resistance. J. Biol. Chem. 2009, 284, 7307–7317. [Google Scholar] [CrossRef] [Green Version]
- Belkhadir, Y.; Nimchuk, Z.; Hubert, D.A.; Mackey, D.; Dangl, J.L. Arabidopsis RIN4 negatively regulates disease resistance mediated by RPS2 and RPM1 downstream or independent of the NDR1 signal modulator and is not required for the virulence functions of bacterial type III effectors AvrRpt2 or AvrRpm1. Plant Cell 2004, 16, 2822–2835. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.M.; Song, H.R.; Han, S.K.; Han, M.; Kim, C.Y.; Park, J.; Lee, Y.H.; Jeon, J.S.; Noh, Y.S.; Noh, B. HDA19 is required for the repression of salicylic acid biosynthesis and salicylic acid-mediated defense responses in Arabidopsis. Plant J. 2012, 71, 135–146. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Yuan, X.; Zhou, T.; Fan, Y.; Zhou, Y. Arabidopsis is susceptible to rice stripe virus infections. J. Phytopathol. 2011, 159, 767–772. [Google Scholar] [CrossRef]
- Wu, S.W.; Wang, H.W.; Yang, Z.D.; Kong, L.R. Expression comparisons of pathogenesis-related (PR) genes in wheat in response to infection/infestation by Fusarium, Yellow dwarf virus (YDV) aphid-transmitted and Hessian Fly. J. Integr. Agric. 2014, 13, 926–936. [Google Scholar] [CrossRef]
- Menke, F.L.H.; Kang, H.G.; Chen, Z.; Jeong, M.P.; Kumar, D.; Klessig, D.F. Tobacco transcription factor WRKY1 is phosphorylated by the MAP kinase SIPK and mediates HR-like cell death in tobacco. Mol. Plant Microbe Interact. 2005, 18, 1027–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, C.; Hwang, S.H.; Fang, I.R.; Kwon, S., II; Park, S.R.; Ahn, I.; Kim, J.B.; Hwang, D.J. Molecular characterization of Oryza sativa WRKY6, which binds to W-box-like element 1 of the Oryza sativa pathogenesis-related (PR) 10a promoter and confers reduced susceptibility to pathogens. New Phytol. 2015, 208, 846–859. [Google Scholar] [CrossRef]
- Birkenbihl, R.P.; Liu, S.; Somssich, I.E. Transcriptional events defining plant immune responses. Curr. Opin. Plant Biol. 2017, 38, 1–9. [Google Scholar] [CrossRef]
- Collum, T.D.; Stone, A.L.; Sherman, D.J.; Rogers, E.E.; Dardick, C.; Culver, J.N. Translatome profiling of Plum pox virus–infected leaves in European plum reveals temporal and spatial coordination of defense responses in phloem tissues. Mol. Plant Microbe Interact. 2020, 33, 66–77. [Google Scholar] [CrossRef]
- Li, S.J.; Castillo-González, C.; Yu, B.; Zhang, X.R. The functions of plant small RNAs in development and in stress responses. Plant J. 2017, 90, 654–670. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.W.; Luo, L.X.; Li, J.Q.; Liu, P.F.; Chen, X.Y.; Hao, J.J. Pollen and seed transmission of Cucumber green mottle mosaic virus in cucumber. Plant Pathol. 2014, 63, 72–77. [Google Scholar] [CrossRef]
- Conesa, A.; Götz, S.; García-Gómez, J.M.; Terol, J.; Talón, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, Z.; Zhou, X.; Ling, Y.; Zhang, Z.; Su, Z. AgriGO: A GO analysis toolkit for the agricultural community. Nucleic Acids Res. 2010, 38, 64–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variety | Disease Index Calculated at 15 DAG | Disease Index Calculated at 30 DAG |
|---|---|---|
| Yushu No.2 | 12.14 ± 1.01abAB | 17.86 ± 1.01cB |
| Yushu No.4 | 5.71 ± 0.00abcABC | 10.00 ± 4.04cBC |
| Yushu No.6 | 2.86 ± 0.00bcdABCD | 7.86 ± 3.03cdBC |
| Yushu 12 | 2.86 ± 0.00bcdABCD | 5.71 ± 0.00cdBCD |
| Xushu 22 | 12.14 ± 1.01abAB | 26.43 ± 3.03bB |
| Ning 4-6 | 35.00 ± 1.01aA | 63.57 ± 11.11bA |
| Ningzishu No.1 | 23.57 ± 1.01aAB | 75.00 ± 13.13aA |
| Unigene | Enzyme | log2 Ratio | Up/Downregulation |
|---|---|---|---|
| Unigene0031486 | phenylalanine ammonia lyase (PAL, EC 4.3.1.24) | −6.31 | Down |
| Unigene0032806 | cinnamate 4-hydroxylase (C4H, EC 1.14.13.11) | −2.14 | Down |
| Unigene0033816 | chalcone synthase (CHS,EC 2.3.1.74) | −16.48 | Down |
| Unigene0017175 | chalcone isomerase (CHI, EC 5.5.1.6) | −2.10 | Down |
| Unigene0033967 | flavonoid 3’-hydroxylase (F3′H, EC 1.14.13.21) | −13.36 | Down |
| Unigene0028067 | flavanone 3-hydroxyrase (F3H, EC 1.14.11.9) | −16.43 | Down |
| Unigene0036071 | dihydroflavonol 4-reductase (DFR, EC 1.1.1.219) | −15.59 | Down |
| Unigene0035228 | anthocyanidin synthase (ANS, EC 1.14.11.19) | −16.52 | Down |
| Unigene0005028 | flavonol synthase (FLS, EC 1.14.11.23) | −11.39 | Down |
| Unigene0014446 | hydroxycinnamoyl-CoA: shikimate O- hydroxycinnamoyltransferase (HCT, EC 2.3.1.133) | −1.89 | Down |
| Unigene0033185 | caffeoyl-CoA 3-O-methyltransferase 5 (CCoAOMT, EC 2.1.1.104) | −1.56 | Down |
| Unigene0029746 | serine carboxypeptidase (EC 2.3.1.91) | −1.01 | Down |
| Unigene0036646 | β-glucosidase 40 (EC 3.2.1.21) | −1.87 | Down |
| Unigene0015200 | 4-coumarate-COA ligase (4CL, EC 6.2.1.12) | 1.53 | Up |
| Unigene0001094 | Ferulate-5-hydroxylase (F5H, EC 1.14.13) | 11.81 | Up |
| Unigene0016523 | cinnamoyl-CoA reductase (CCR, EC 1.2.1.44) | 1.24 | Up |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, K.; Lu, H.; Wan, C.; Tang, D.; Zhao, Y.; Luo, K.; Li, S.; Wang, J. The Spread and Transmission of Sweet Potato Virus Disease (SPVD) and Its Effect on the Gene Expression Profile in Sweet Potato. Plants 2020, 9, 492. https://doi.org/10.3390/plants9040492
Zhang K, Lu H, Wan C, Tang D, Zhao Y, Luo K, Li S, Wang J. The Spread and Transmission of Sweet Potato Virus Disease (SPVD) and Its Effect on the Gene Expression Profile in Sweet Potato. Plants. 2020; 9(4):492. https://doi.org/10.3390/plants9040492
Chicago/Turabian StyleZhang, Kai, Huixiang Lu, Chuanfang Wan, Daobin Tang, Yong Zhao, Kai Luo, Shixi Li, and Jichun Wang. 2020. "The Spread and Transmission of Sweet Potato Virus Disease (SPVD) and Its Effect on the Gene Expression Profile in Sweet Potato" Plants 9, no. 4: 492. https://doi.org/10.3390/plants9040492