Constitutive and Induced Expression of Total Phenol and Phenol Oxidases in Wheat Genotypes Ranging in Resistance/Susceptibility to the Root-Lesion Nematode Pratylenchus thornei



Abstract
:1. Introduction
2. Results
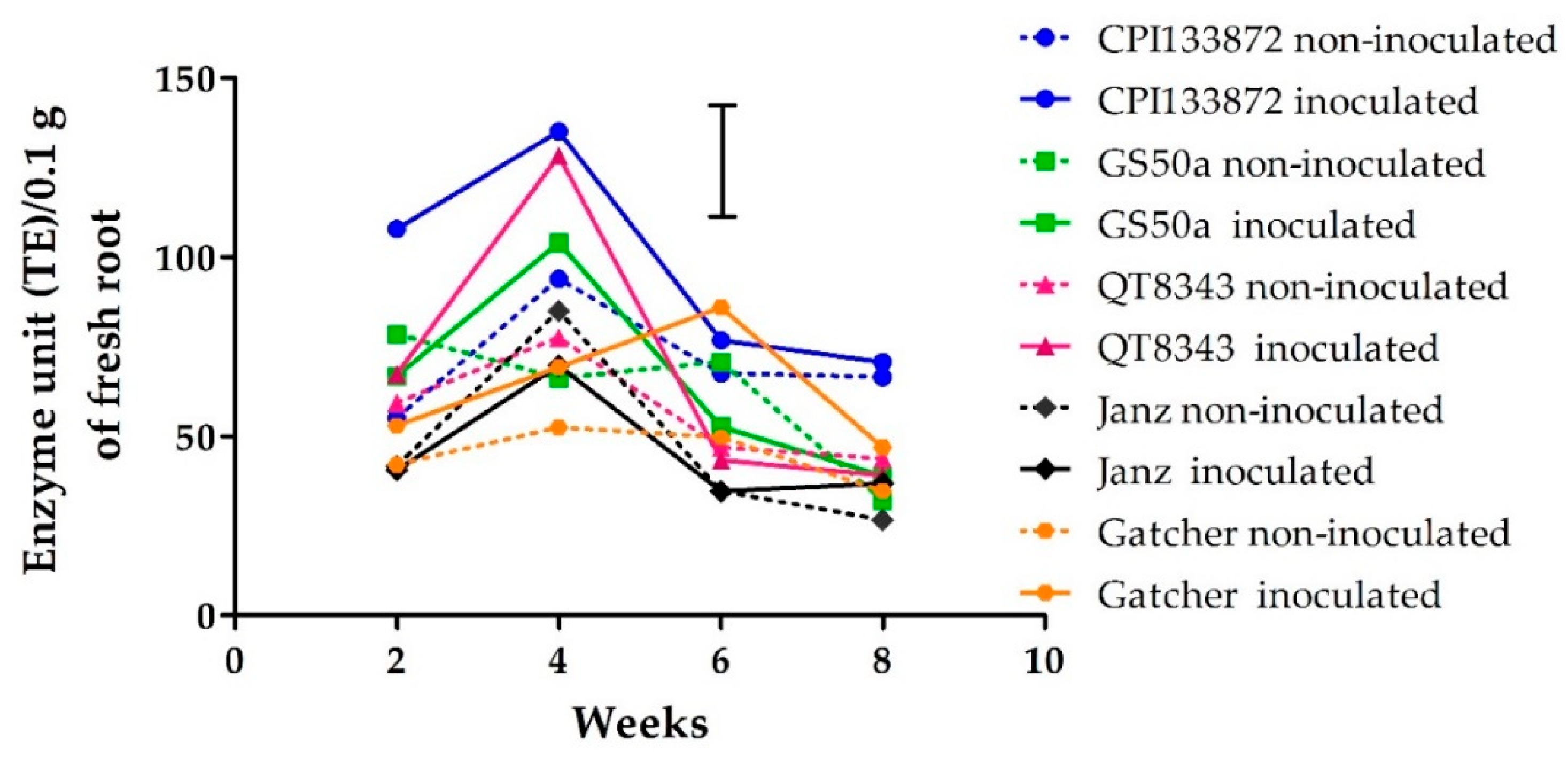
2.1. Experiment 1: Accumulation of Total Phenols Over Time
2.2. Experiment 2: Constitutive Levels of Total Phenols in Gnotobiotic Conditions
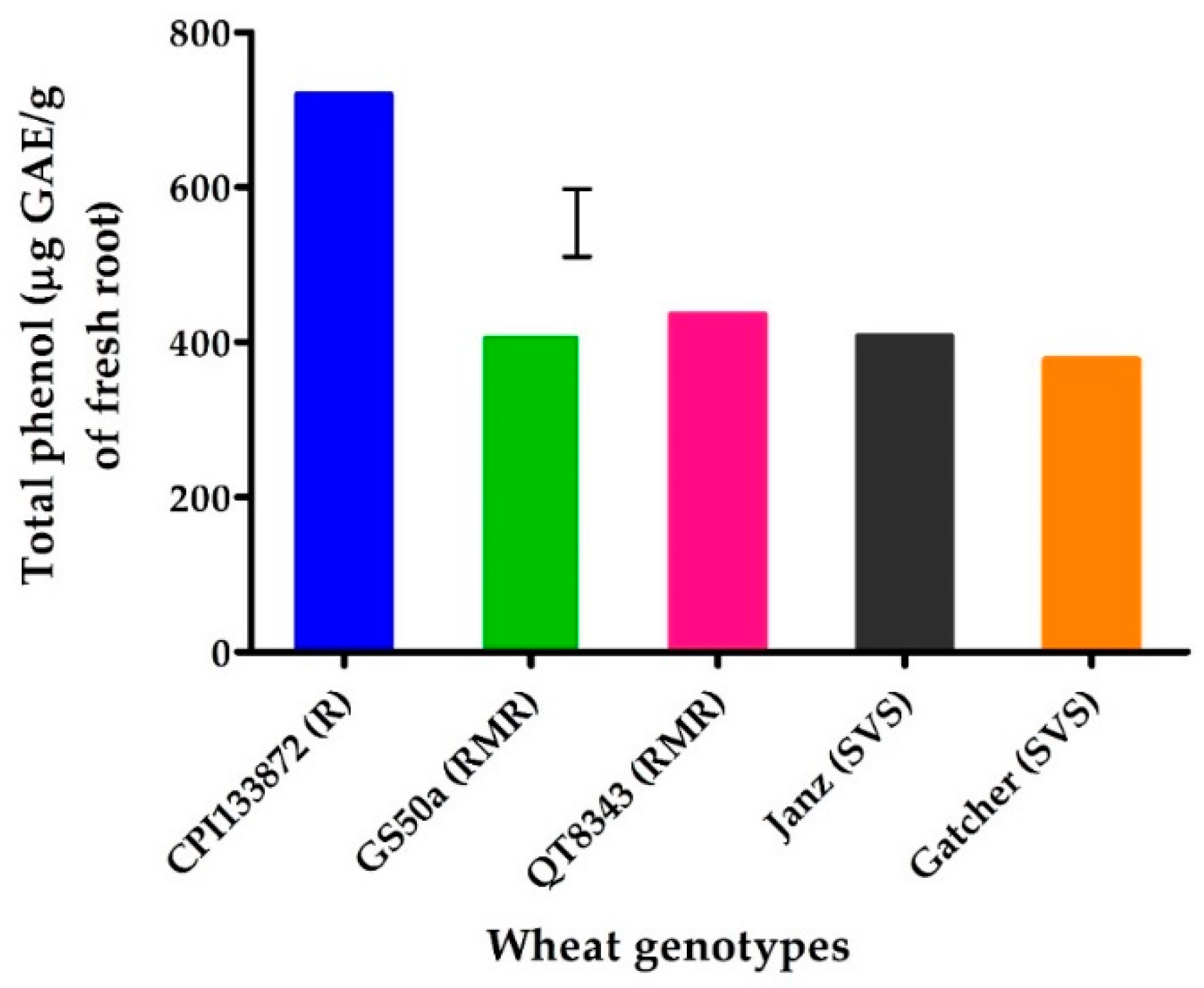
2.3. Experiment 3: Total Phenols in Wheat Genotypes Ranging in Resistance/Susceptibility to P. thornei
2.4. Polyphenol Oxidase Enzyme Activity
2.5. Peroxidase Enzyme Activity
2.6. Nematode Quantification
2.7. Determination of Dry Shoot Weight and Fresh Root Weight
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.1.1. Experiment 1: The Accumulation of Total Phenols and Oxidative Enzymes (PPO and POD) Over Time
4.1.2. Experiment 2: Constitutive Levels of Total Phenols in Wheat Roots under Gnotobiotic Conditions
4.1.3. Experiment 3: Total Phenols in Wheat Genotypes Ranging in Resistance/Susceptibility to P. thornei
4.2. Sample Collection and Storage
4.3. Estimation of Total Phenols
4.4. Preparation of Enzyme Extracts and Optimization of Protocol for Microplate Reader
4.5. Polyphenol Oxidase Enzyme Assay
4.6. Peroxidase Enzyme Assay
4.7. Nematode Quantification
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Smiley, R.W.; Nicol, J.M. Nematodes which challenge global wheat production. In Wheat Science and Trade; Carver, B.F., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2009; Volume 10, pp. 171–187. [Google Scholar]
- Thompson, J.P.; Clewett, T.G.; Sheedy, J.G.; Reen, R.A.; O’Reilly, M.M.; Bell, K.L. Occurrence of root-lesion nematodes (Pratylenchus thornei and P. neglectus) and stunt nematode (Merlinius brevidens) in the northern grain region of Australia. Australas. Plant Path. 2010, 39, 254–264. [Google Scholar] [CrossRef]
- Fosu-Nyarko, J.; Jones, M.G.K. Advances in understanding the molecular mechanisms of root lesion nematode host interactions. Annu. Rev. Phytopathol. 2016, 54, 253–278. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.P.; Mackenzie, J.; Sheedy, G.H. Root-lesion nematode (Pratylenchus thornei) reduces nutrient response, biomass and yield of wheat in sorghum–fallow–wheat cropping systems in a subtropical environment. Field Crops Res. 2012, 137, 126–140. [Google Scholar] [CrossRef]
- Whish, J.P.M.; Thompson, J.P.; Clewett, T.G.; Lawrence, J.L.; Wood, J. Pratylenchus thornei populations reduce water uptake in intolerant wheat cultivars. Field Crops Res. 2014, 161, 1–10. [Google Scholar] [CrossRef]
- Baxter, R.I.; Blake, C.D. Pratylenchus thornei-a cause of root necrosis in wheat. Nematologica 1968, 14, 351–361. [Google Scholar]
- Vanstone, V.; Hollaway, G.; Stirling, G. Managing nematode pests in the southern and western regions of the Australian cereal industry: Continuing progress in a challenging environment. Australas. Plant. Path. 2008, 37, 220–234. [Google Scholar] [CrossRef]
- Thompson, J.P.; Clewett, T.G.; O’Reilly, M.M. Optimising initial population density, growth time and nitrogen nutrition for assessing resistance of wheat cultivars to root-lesion nematode (Pratylenchus thornei). Australas. Plant. Path. 2015, 44, 133–147. [Google Scholar] [CrossRef]
- Trudgill, D.L. Resistance to and tolerance of plant parasitic nematodes in plants. Annu. Rev. Phytopathol. 1991, 29, 167–192. [Google Scholar] [CrossRef]
- Thompson, J.P. Resistance to root-lesion nematodes (Pratylenchus thornei and P. neglectus) in synthetic hexaploid wheats and their durum and Aegilops tauschii parents. Aust. J. Agric. Res. 2008, 59, 432–446. [Google Scholar] [CrossRef]
- Thompson, J.P.; Brennan, P.S.; Clewett, T.G.; Sheedy, J.G.; Seymour, N.P. Progress in breeding wheat for tolerance and resistance to root-lesion nematode (Pratylenchus thornei). Australas. Plant. Path. 1999, 28, 45–52. [Google Scholar] [CrossRef]
- Thompson, J.P.; O’Reilly, M.M.; Clewett, T.G. Resistance to the root-lesion nematode Pratylenchus thornei in wheat landraces and cultivars from the West Asia and North Africa (WANA) region. Crop Pasture Sci. 2009, 60, 1209–1217. [Google Scholar] [CrossRef]
- Sheedy, J.G.; Thompson, J.P. Resistance to the root-lesion nematode Pratylenchus thornei of Iranian landrace wheat. Australas. Plant. Path. 2009, 38, 478–489. [Google Scholar] [CrossRef]
- Linsell, K.J.; Rahman, M.S.; Taylor, J.D.; Davey, R.S.; Gogel, B.J.; Wallwork, H.; Forrest, K.L.; Hayden, M.J.; Taylor, S.P.; Oldach, K.H. QTL for resistance to root lesion nematode (Pratylenchus thornei) from a synthetic hexaploid wheat source. Theor. Appl. Genet. 2014, 127, 1409–1421. [Google Scholar] [CrossRef] [PubMed]
- Zwart, R.S.; Thompson, J.P.; Sheedy, J.G.; Nelson, J.C. Mapping quantitative trait loci for resistance to Pratylenchus thornei from synthetic hexaploid wheat in the International Triticeae Mapping Initiative (ITMI) population. Austral. J. Agric. Res. 2006, 57, 525–530. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.P.; Haak, M.I. Resistance to root-lesion nematode (Pratylenchus thornei) in Aegilops tauschii Coss. the D-genome donor to wheat. Aust. J. Agric. Res. 1997, 48, 553–559. [Google Scholar] [CrossRef]
- Sheedy, J.G.; Thompson, J.P.; Kelly, A. Diploid and tetraploid progenitors of wheat are valuable sources of resistance to the root-lesion nematode Pratylenchus thornei. Euphytica 2012, 186, 377–391. [Google Scholar] [CrossRef]
- Zwart, R.S.; Thompson, J.P.; Godwin, I.D. Genetic analysis of resistance to root-lesion nematode (Pratylenchus thornei) in wheat. Plant Breed. 2004, 123, 209–212. [Google Scholar] [CrossRef]
- Zwart, R.S.; Thompson, J.P.; Milgate, A.W.; Bansal, U.K.; Williamson, P.M.; Raman, H.; Bariana, H.S. QTL mapping of multiple foliar disease and root-lesion nematode resistances in wheat. Mol. Breed. 2010, 26, 107–124. [Google Scholar] [CrossRef]
- Schmidt, A.; McIntyre, C.; Thompson, J.; Seymour, N.; Liu, C. Quantitative trait loci for root lesion nematode (Pratylenchus thornei) resistance in Middle-Eastern landraces and their potential for introgression into Australian bread wheat. Austral. J. Agric. Res. 2005, 56, 1059–1068. [Google Scholar] [CrossRef]
- Linsell, K.J.; Riley, I.T.; Davies, K.A.; Oldach, K.H. Characterization of resistance to Pratylenchus thornei (Nematoda) in wheat (Triticum aestivum): Attraction, penetration, motility, and reproduction. Phytopathology 2014, 104, 174–187. [Google Scholar] [CrossRef] [Green Version]
- Freeman, B.C.; Beattie, G.A. An overview of plant defenses against pathogens and herbivores. Plant Health Instr. 2008, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Veech, J.A. Phytoalexins and their role in the resistance of plants to nematodes. Nematology 1982, 14, 2–9. [Google Scholar]
- Dixon, R.A. Natural products and plant disease resistance. Nature 2001, 411, 843–847. [Google Scholar] [CrossRef] [PubMed]
- Sinha, M.; Singh, R.P.; Kushwaha, G.S.; Iqbal, N.; Singh, A.; Kaushik, S.; Kaur, P.; Sharma, S.; Singh, T.P. Current overview of allergens of plant pathogenesis related protein families. Sci. World J. 2014, 2014, 158. [Google Scholar] [CrossRef] [Green Version]
- Farkas, G.L.; Kiraaly, Z. Role of phenolic compounds in the physiology of plant diseases and disease resistance. J. Phytopathol. 1962, 44, 105–150. [Google Scholar] [CrossRef]
- Giebel, J. Mechanism of resistance to plant nematodes. Annu. Rev. Phytopathol. 1982, 20, 257–279. [Google Scholar] [CrossRef]
- Townshend, J.L. The pathogenicity of Pratylenchus penetrans to strawberry. Can. J. Plant Sci. 1963, 43, 75–78. [Google Scholar] [CrossRef] [Green Version]
- Acedo, J.R.; Rohde, R.A. Histochemical root pathology of Brassica oleracea capitata L. infected by Pratylenchus penetrans (Cobb) Filipjev and Schuurmans Stekhoyen (Nematoda: Tylenchidae). J. Nematol. 1971, 3, 62–68. [Google Scholar]
- Backiyarani, S.; Uma, S.; Sundararaju, P.; Mayilvaganan, M.; Saraswathi, M.S.; Arunkumar, G. Time course expression studies during Musa-Pratylenchus coffeae interaction. Indian J. Hortic. 2013, 70, 217–222. [Google Scholar]
- Vaganan, M.M.; Ravi, I.; Nandakumar, A.; Sarumathi, S.; Sundararaju, P.; Mustaffa, M. Phenylpropanoid enzymes, phenolic polymers and metabolites as chemical defenses to infection of Pratylenchus coffeae in roots of resistant and susceptible bananas (Musa spp.). Indian J. Exp. Biol. 2014, 52, 252–260. [Google Scholar]
- Mehta, U.; Kathiresan, T. Effect of Pratylenchus zeae infection on the expression of isozyme activities in resistant and susceptible sugarcane clones. Nematology 2005, 7, 677–688. [Google Scholar] [CrossRef]
- Kubalt, K. The role of phenolic compounds in plant resistance. Food Sci. Biotechnol. 2016, 97–108. [Google Scholar]
- Matern, U.; Kneusel, R.E. Phenolic compounds in plant disease resistance. Phytoparasitica 1988, 16, 153–170. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Mandal, S.M.; Chakraborty, D.; Dey, S. Phenolic acids act as signaling molecules in plant-microbe symbioses. Plant Signal Behav. 2010, 5, 359–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devi, A.N.; Ponnuswami, V.; Sundararaju, P.; Soorianathasundaram, K.; Sathiamoorthy, S.; Uma, S.; Van Den Bergh, I. Mechanism of resistance in banana cultivars against root-lesion nematode, Pratylenchus coffeae. Indian J. Nematol. 2007, 37, 138–144. [Google Scholar]
- Baldridge, G.D.; O’Neill, N.R.; Samac, D.A. Alfalfa (Medicago sativa L.) resistance to the root-lesion nematode, Pratylenchus penetrans: Defence-response gene mRNA and isoflavonoid phytoalexin levels in roots. Plant Mol. Biol. 1998, 38, 999–1010. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.P.; Zwart, R.S.; Butler, D. Inheritance of resistance to root-lesion nematodes (Pratylenchus thornei and P. neglectus) in five doubled-haploid populations of wheat. Euphytica 2012, 188, 209–219. [Google Scholar] [CrossRef]
- Saini, R.S.; Arora, Y.K.; Chawla, H.K.L.; Wagle, D.S. Total phenols and sugar content in wheat cultivars resistant and susceptible to Ustilago nuda (Jens.) Rostrup. Biochem. Physiol. Pflanz. 1988, 183, 89–93. [Google Scholar] [CrossRef]
- Zadoks, J.C.; Chang, T.T.; Konzak, C.F. A decimal code for the growth stages of cereals. Weed Res. 1974, 14, 415–421. [Google Scholar] [CrossRef]
- Iannucci, A.; Fragasso, M.; Platani, C.; Papa, R. Plant growth and phenol compounds in the rhizosphere soil of wild oat (Avena fatua L.). Front. Plant Sci. 2013, 509, 1–7. [Google Scholar]
- Rahman, M.S.; Linsell, K.J.; Taylor, J.D.; Hayden, M.J.; Collins, N.C.; Oldach, K.H. Fine mapping of root lesion nematode (Pratylenchus thornei) resistance loci on chromosomes 6D and 2B of wheat. Theor. Appl. Genet. 2020, 133, 635–652. [Google Scholar] [CrossRef] [PubMed]
- Mather, D.; (University of Adelaide). Personal communication, 2017.
- Valette, C.; Andary, C.; Geiger, J.P.; Sarah, J.L.; Nicole, M. Histochemical and cytochemical investigations of phenols in roots of banana infected by the burrowing nematode Radopholus similis. Phytopathology 1998, 88, 1141–1148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, C.; Rohde, R.A. Phenol accumulation related to resistance in tomato to infection by root-knot and lesion nematodes. J. Nematol. 1973, 5, 253–258. [Google Scholar]
- Taranto, F.; Pasqualone, A.; Mangini, G.; Tripodi, P.; Miazzi, M.M.; Pavan, S.; Montemurro, C. Polyphenol oxidases in crops: Biochemical, physiological and genetic aspects. Int. J. Mol. Sci. 2017, 18, 377. [Google Scholar] [CrossRef]
- Constabel, C.P.; Barbehenn, R. Defensive roles of polyphenol oxidase in plants. In Induced, Plant Resistance to Herbivory; Springer: Berlin, Germany, 2008; pp. 253–270. [Google Scholar]
- Pandey, V.; Awasthi, M.; Singh, S.; Tiwari, S.; Dwivedi, U. A comprehensive review on function and application of plant peroxidases. Biochem. Anal. Biochem. 2017, 6, 1–16. [Google Scholar] [CrossRef]
- Vaughn, K.C.; Duke, S.O. Function of polyphenol oxidase in higher plants. Physiol. Plant. 1984, 60, 106–112. [Google Scholar] [CrossRef]
- Siguemoto, E.S.; Gut, J.A.W. Validation of spectrophotometric microplate methods for polyphenol oxidase and peroxidase activities analysis in fruits and vegetables. Food Sci. Technol. 2017, 37, 148–153. [Google Scholar] [CrossRef] [Green Version]
- Talavera, M.; Vanstone, V.A. Monitoring Pratylenchus thornei densities in soil and roots under resistant (Triticum turgidum durum) and susceptible (Triticum aestivum) wheat cultivars. Phytoparasitica 2001, 29, 29–35. [Google Scholar] [CrossRef]
- Liu, Q.; Luo, L.; Zheng, L. Lignins: Biosynthesis and biological functions in plants. Int. J. Mol. Sci. 2018, 19, 335. [Google Scholar] [CrossRef] [Green Version]
- Melillo, M.; Bleve-Zacheo, T.; Zacheo, G. Role of peroxidase and esterase isoenzymes in pea roots infected with Heterodera goettingiana. Nematol. Mediterr. 1992, 20, 171–179. [Google Scholar]
- Miedes, E.; Vanholme, R.; Boerjan, W.; Molina, A. The role of the secondary cell wall in plant resistance to pathogens. Front. Plant Sci. 2014, 5, 358. [Google Scholar] [CrossRef] [Green Version]
- Dirk, D.W.; Elsen, A.; Mariama, K.; Dhakshinamoorthy, S. Phenols and lignin are involved in the defence response of banana (Musa) plants to Radopholus similis infection. Nematology 2014, 16, 565–576. [Google Scholar]
- Wuyts, N.; Swennen, R.; De Waele, D. Effects of plant phenylpropanoid pathway products and selected terpenoids and alkaloids on the behaviour of the plant-parasitic nematodes Radopholus similis, Pratylenchs penetrans and Meloidogyne incognita. Nematology 2006, 8, 89–101. [Google Scholar] [CrossRef]
- Rich, J.R.; Keen, N.T.; Thomason, I.J. Association of coumestans with the hypersensitivity of lima bean roots to Pratyenchus scriberni. Physiol. Plant Pathol. 1977, 10, 105–116. [Google Scholar] [CrossRef]
- Steinfath, M.; Strehmel, N.; Peters, R.; Schauer, N.; Groth, D.; Hummel, J.; Steup, M.; Selbig, J.; Kopka, J.; Geigenberger, P. Discovering plant metabolic biomarkers for phenotype prediction using an untargeted approach. Plant Biotechnol. J. 2010, 8, 900–911. [Google Scholar] [CrossRef]
- Fernandez, O.; Urrutia, M.; Bernillon, S.; Giauffret, C.; Tardieu, F.; Le Gouis, J.; Langlade, N.; Charcosset, A.; Moing, A.; Gibon, Y. Fortune telling: Metabolic markers of plant performance. Metabolomics 2016, 12, 158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.P. Treatments to eliminate root-lesion nematode (Pratylenchus thornei Sher & Allen) from a vertisol. Nematologica 1990, 36, 123–127. [Google Scholar]
- Sheedy, J.G.; McKay, A.C.; Lewis, J.; Vanstone, V.A.; Fletcher, S.; Kelly, A.; Thompson, J.P. Cereal cultivars can be ranked consistently for resistance to root-lesion nematodes (Pratylenchus thornei and P. neglectus) using diverse procedures. Australas. Plant. Path. 2015, 44, 175–182. [Google Scholar] [CrossRef]
- Thompson, J.; Clewett, T.; O’Reilly, M. Temperature response of root-lesion nematode (Pratylenchus thornei) reproduction on wheat cultivars has implications for resistance screening and wheat production. Ann. Appl Biol. 2015, 167, 1–10. [Google Scholar] [CrossRef]
- Wu, H.; Pratley, J.; Lemerle, D.; Haig, T. Laboratory screening for allelopathic potential of wheat (Triticum aestivum) accessions against annual ryegrass (Lolium rigidum). Aust. J. Agric. Res. 2000, 51, 259–266. [Google Scholar] [CrossRef]
- Thompson, J.P.; Sheedy, J.G.; Robinson, N.A. Resistance of wheat genotypes to root-lesion nematode (Pratylenchus thornei) can be used to predict final nematode population densities, crop greenness, and grain yield in the field. Phytopathology 2020, 110, 505–516. [Google Scholar] [CrossRef] [PubMed]
- Ainswort, E.A.; Gillespie, K.M. Estimation of total phenolic content and other oxidation substrates in plant tissues using Folin–Ciocalteu reagent. Nat. Protoc. 2007, 2, 875–877. [Google Scholar] [CrossRef]
- Bisswanger, H. Enzyme assays. Perspect. Sci. 2014, 1, 41–55. [Google Scholar] [CrossRef] [Green Version]
- Kar, M.; Mishra, D. Catalase, peroxidase, and polyphenoloxidase activities during rice leaf senescence. Plant Physiol. 1976, 57, 315–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitehead, A.G.; Hemming, J.R. A comparison of some quantitative methods of extracting small vermiform nematodes from soil. Ann. Appl. Biol. 1965, 55, 25–38. [Google Scholar] [CrossRef]
- Bybd, D., Jr.; Kirkpatrick, T.; Barker, K. An improved technique for clearing and staining plant tissues for detection of nematodes. J. Nematol. 1983, 15, 142. [Google Scholar]
- Fiske, M.L.; Thompson, J.P.; Clewett, T.G. Stain tubes—An aid for staining vesicular-arbuscular mycorrhizae. Australas. Plant. Path. 1989, 18, 24–25. [Google Scholar] [CrossRef]
- R Core Team. R: A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2017; Available online: https://www.R-project.org/ (accessed on 19 December 2018).
- VSN, International. GenStat for Windows, 17th ed.; VSN International: Hemel Hempstead, UK, 2014. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | ln (P. thornei/g of Dry Root + 1) (Back Transformed Mean)) | ln (P. thornei/kg of Soil and Root + 1) (Back Transformed Mean) |
|---|---|---|
| CPI133872 | 7.859 b (2588) | 8.295 b (4004) |
| GS50a | 7.783 b (2399) | 8.536 b (5095) |
| QT8343 | 8.074 ab (3210) | 8.733 b (6204) |
| Janz | 8.472 ab (4779) | 9.285 ab (10775) |
| Gatcher | 8.997 a (8079) | 10.018 a (22426) |
| Genotype | Origin | Rating [65] |
|---|---|---|
| CPI133872 | Synthetic hexaploid | R |
| CPI133872_Janz DH083 | CPI133872 derived line | R-MR |
| QT8343 | GS50a derived line | R-MR |
| GS50a | Selection from Gatcher | R-MR |
| Morocco 426 | Landrace | R-MR |
| USQW15008 | CPI133872_Janz_DH083 derived line | MR |
| CPI133859 | Synthetic hexaploid | MR |
| Iraq 43 | Landrace | MR |
| QT8447 | GS50a derived line | MR |
| Gauntlet | Cultivar | MR-MS |
| Suntop | Cultivar | MR-MS |
| Hartog | Cultivar | MS |
| Pelsart | Cultivar | MS |
| Gregory | Cultivar | MS-S |
| Wylie | Cultivar | MS-S |
| Batavia | Cultivar | S |
| Hume | Cultivar | S |
| Janz | Cultivar | S-VS |
| Gatcher | Cultivar | S-VS |
| Petrie | Cultivar | VS |
| Darwin | Cultivar | VS |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rahaman, M.M.; Zwart, R.S.; Thompson, J.P. Constitutive and Induced Expression of Total Phenol and Phenol Oxidases in Wheat Genotypes Ranging in Resistance/Susceptibility to the Root-Lesion Nematode Pratylenchus thornei. Plants 2020, 9, 485. https://doi.org/10.3390/plants9040485
Rahaman MM, Zwart RS, Thompson JP. Constitutive and Induced Expression of Total Phenol and Phenol Oxidases in Wheat Genotypes Ranging in Resistance/Susceptibility to the Root-Lesion Nematode Pratylenchus thornei. Plants. 2020; 9(4):485. https://doi.org/10.3390/plants9040485
Chicago/Turabian StyleRahaman, Md Motiur, Rebecca S. Zwart, and John P. Thompson. 2020. "Constitutive and Induced Expression of Total Phenol and Phenol Oxidases in Wheat Genotypes Ranging in Resistance/Susceptibility to the Root-Lesion Nematode Pratylenchus thornei" Plants 9, no. 4: 485. https://doi.org/10.3390/plants9040485