Feasible Production of Lignans and Neolignans in Root-Derived In Vitro Cultures of Flax (Linum usitatissimum L.)

 ,
,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Establishment and Growth Dynamics of Callus Culture
2.2. Establishment and Growth Dynamics of Adventitious Root Culture
2.3. Phenolics Production in Callus and Adventitious Root Culture
2.4. Flavonoids Production in Callus and Adventitious Root Culture
2.5. Antioxidant Activity of Callus and Adventitious Root Cultures
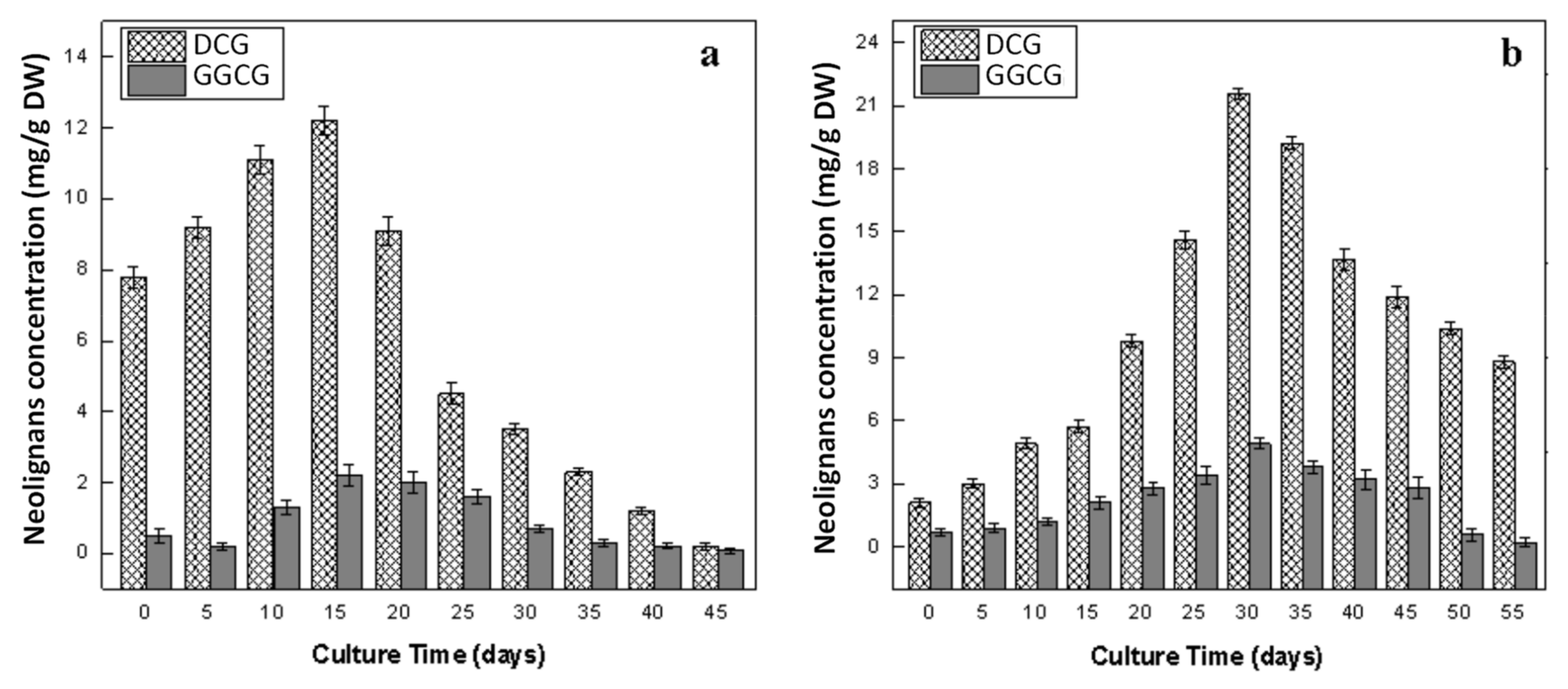
2.6. Production of Lignans in Callus and Adventitious Root Cultures
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Plant Material and Establishment of Callus Culture
3.3. Optimization and Growth Dynamics of Callus Culture
3.4. Establishment of Adventitious Root Culture
3.5. Optimization and Growth Dynamics of Adventitious Root Culture
3.6. Determination of Total Polyphenols Contents and Antioxidant Activity
3.7. HPLC (High Performance Liquid Chromatography) Analysis of Lignans and Neolignans
3.8. Statistical Analysis of Data
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lainé, E.; Hano, C.; Lamblin, F. Les lignanes phytoestrogènes du lin sont-ils des bienfaiteurs méconnus? Phytotherapie 2007. [Google Scholar] [CrossRef]
- Lamblin, F.; Hano, C.; Fliniaux, O.; Mesnard, F.; Fliniaux, M.-A.; Lainé, É. Interest of lignans in prevention and treatment of cancers. Med. Sci. 2008, 24. [Google Scholar] [CrossRef] [Green Version]
- Lainé, E.; Hano, C.; Lamblin, F. Lignans. In Chemoprevention of Cancer and DNA Damage by Dietary Factors; Knasmüller, S., DeMarini, D.M., Johnson, I.T.C.G., Eds.; Wiley-VCH: Weinheim, Germany, 2009; pp. 555–577. [Google Scholar]
- Szewczyk, M.; Abarzua, S.; Schlichting, A.; Nebe, B.; Piechulla, B.; Briese, V.; Richter, D.-U. Effects of extracts from Linum usitatissimum on cell vitality, proliferation and cytotoxicity in human breast cancer cell lines. J. Med. Plants Res. 2014, 8, 237–245. [Google Scholar]
- Wang, H.; Wang, J.; Guo, X.; Brennan, C.S.; Li, T.; Fu, X.; Chen, G.; Liu, R.H. Effect of germination on lignan biosynthesis, and antioxidant and antiproliferative activities in flaxseed (Linum usitatissimum L.). Food Chem. 2016, 205, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Bussmann, R.W.; Batsatsashvili, K.; Kikvidze, Z.; Paniagua-Zambrana, N.Y.; Khutsishvili, M.; Maisaia, I.; Sikharulidze, S.; Tchelidze, D. Linum usitatissimum L. Linaceae. Ethnobot. Mt. Reg. Far East. Eur. Ural. North. Cauc. Turk. Iran. 2019, 1–9. [Google Scholar]
- Oomah, B.D. Flaxseed as a functional food source. J. Sci. Food Agric. 2001, 81, 889–894. [Google Scholar] [CrossRef]
- Corbin, C.; Fidel, T.; Leclerc, E.A.; Barakzoy, E.; Sagot, N.; Falguiéres, A.; Renouard, S.; Blondeau, J.; Ferroud, C.; Doussot, J.; et al. Development and validation of an efficient ultrasound assisted extraction of phenolic compounds from flax (Linum usitatissimum L.) seeds. Ultrason. Sonochem. 2015, 26, 176–185. [Google Scholar] [CrossRef]
- Ramsay, A.; Fliniaux, O.; Quéro, A.; Molinié, R.; Demailly, H.; Hano, C.; Paetz, C.; Roscher, A.; Grand, E.; Kovensky, J.; et al. Kinetics of the incorporation of the main phenolic compounds into the lignan macromolecule during flaxseed development. Food Chem. 2017, 217. [Google Scholar] [CrossRef]
- Beejmohun, V.; Fliniaux, O.; Grand, É.; Lamblin, F.; Bensaddek, L.; Christen, P.; Kovensky, J.; Fliniaux, M.-A.; Mesnard, F. Microwave-assisted extraction of the main phenolic compounds in flaxseed. Phytochem. Anal. 2007, 18, 275–282. [Google Scholar] [CrossRef]
- Anjum, S.; Abbasi, B.H.; Doussot, J.; Favre-réguillon, A.; Hano, C.; Haider, B.; Doussot, J.; Favre-réguillon, A.; Hano, C. Effects of photoperiod regimes and ultraviolet-C radiations on biosynthesis of industrially important lignans and neolignans in cell cultures of Linum usitatissimum L. (Flax ). J. Photochem. Photobiol. B Biol. 2017, 167, 216–227. [Google Scholar] [CrossRef]
- Anjum, S.; Abbasi, B.H.; Hano, C. Trends in accumulation of pharmacologically important antioxidant-secondary metabolites in callus cultures of Linum usitatissimum L. Plant. Cell. Tissue Organ. Cult. 2017, 129. [Google Scholar] [CrossRef]
- Zahir, A.; Ahmad, W.; Nadeem, M.; Giglioli-Guivarc’h, N.; Hano, C.; Abbasi, B.H. In vitro cultures of Linum usitatissimum L.: Synergistic effects of mineral nutrients and photoperiod regimes on growth and biosynthesis of lignans and neolignans. J. Photochem. Photobiol. B Biol. 2018, 187, 141–150. [Google Scholar] [CrossRef]
- Ahmad, W.; Zahir, A.; Nadeem, M.; Garros, L.; Drouet, S.; Renouard, S.; Doussot, J.; Giglioli-Guivarc’h, N.; Hano, C.; Abbasi, B.H. Enhanced production of lignans and neolignans in chitosan-treated flax (Linum usitatissimum L.) cell cultures. Process. Biochem. 2019, 79, 155–165. [Google Scholar] [CrossRef]
- Nadeem, M.; Ahmad, W.; Zahir, A.; Hano, C.; Abbasi, B.H. Salicylic acid-enhanced biosynthesis of pharmacologically important lignans and neo lignans in cell suspension culture of Linum ussitatsimum L. Eng. Life Sci. 2019, 19, 168–174. [Google Scholar] [CrossRef] [Green Version]
- Zahir, A.; Nadeem, M.; Ahmad, W.; Giglioli-Guivarc’h, N.; Hano, C.; Abbasi, B.H. Chemogenic silver nanoparticles enhance lignans and neolignans in cell suspension cultures of Linum usitatissimum L. Plant. Cell Tissue Organ. Cult. 2019, 136, 589–596. [Google Scholar] [CrossRef]
- Ford, J.D.; Huang, K.S.; Wang, H.B.; Davin, L.B.; Lewis, N.G. Biosynthetic pathway to the cancer chemopreventive secoisolariciresinol diglucoside-hydroxymethyl glutaryl ester-linked lignan oligomers in flax (Linum usitatissimum) seed. J. Nat. Prod. 2001, 64, 1388–1397. [Google Scholar] [CrossRef]
- Dalisay, D.S.; Kim, K.W.; Lee, C.; Yang, H.; Rübel, O.; Bowen, B.P.; Davin, L.B.; Lewis, N.G. Dirigent Protein-Mediated Lignan and Cyanogenic Glucoside Formation in Flax Seed: Integrated Omics and MALDI Mass Spectrometry Imaging. J. Nat. Prod. 2015, 78, 1231–1242. [Google Scholar] [CrossRef]
- Corbin, C.; Drouet, S.; Markulin, L.; Auguin, D.; Lainé, É.; Davin, L.B.L.B.; Cort, J.R.J.R.; Lewis, N.G.N.G.; Hano, C. A genome-wide analysis of the flax (Linum usitatissimum L.) dirigent protein family: From gene identification and evolution to differential regulation. Plant. Mol. Biol. 2018, 97, 73–101. [Google Scholar] [CrossRef]
- Moinuddin, S.G.A.; Cort, J.R.; Smith, C.A.; Hano, C.; Davin, L.B.; Lewis, N.G. Linum Lignan and Associated Biochemical Pathways in Human Health and Plant Defense. In Plant Genetics and Genomics: Crops and Models; Springer: Berlin/Heidelberg, Germany, 2019; pp. 167–193. [Google Scholar]
- Von Heimendahl, C.B.I.; Schäfer, K.M.; Eklund, P.; Sjöholm, R.; Schmidt, T.J.; Fuss, E. Pinoresinol–lariciresinol reductases with different stereospecificity from Linum album and Linum usitatissimum. Phytochemistry 2005, 66, 1254–1263. [Google Scholar] [CrossRef]
- Hano, C.; Addi, M.; Bensaddek, L.; Crônier, D.; Baltora-Rosset, S.; Doussot, J.; Maury, S.; Mesnard, F.; Chabbert, B.; Hawkins, S.; et al. Differential accumulation of monolignol-derived compounds in elicited flax (Linum usitatissimum) cell suspension cultures. Planta 2006, 223, 975–989. [Google Scholar] [CrossRef]
- Hano, C.; Martin, I.; Fliniaux, O.; Legrand, B.; Gutierrez, L.; Arroo, R.R.J.; Mesnard, F.; Lamblin, F.; Lainé, E. Pinoresinol-lariciresinol reductase gene expression and secoisolariciresinol diglucoside accumulation in developing flax (Linum usitatissimum) seeds. Planta 2006, 224, 1291–1301. [Google Scholar] [CrossRef]
- Renouard, S.; Tribalatc, M.; Lamblin, F.; Mongelard, G.; Fliniaux, O.; Corbin, C.; Marosevic, D.; Pilard, S.; Demailly, H.; Gutierrez, L.; et al. RNAi-mediated pinoresinol lariciresinol reductase gene silencing in flax (Linum usitatissimum L.) seed coat: Consequences on lignans and neolignans accumulation. J. Plant. Physiol. 2014, 171, 1372–1377. [Google Scholar] [CrossRef] [PubMed]
- Corbin, C.; Drouet, S.; Mateljak, I.; Markulin, L.; Decourtil, C.; Renouard, S.; Lopez, T.; Doussot, J.; Lamblin, F.; Auguin, D.; et al. Functional characterization of the pinoresinol–lariciresinol reductase-2 gene reveals its roles in yatein biosynthesis and flax defense response. Planta 2017, 246, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Hemmati, S.; Von Heimendahl, C.B.I.; Klaes, M.; Alfermann, A.W.; Schmidt, T.J.; Fuss, E. Pinoresinol-Lariciresinol Reductases with Opposite Enantiospecificity Determine the Enantiomeric Composition of Lignans in the Different Organs of Linum usitatissimum L. Planta Med. 2010, 76, 928–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Markulin, L.; Corbin, C.; Renouard, S.; Drouet, S.; Gutierrez, L.; Mateljak, I.; Auguin, D.; Hano, C.; Fuss, E.; Lainé, E. Pinoresinol–lariciresinol reductases, key to the lignan synthesis in plants. Planta 2019, 249, 1695–1714. [Google Scholar] [CrossRef] [PubMed]
- Ghose, K.; Selvaraj, K.; McCallum, J.; Kirby, C.W.; Sweeney-Nixon, M.; Cloutier, S.J.; Deyholos, M.; Datla, R.; Fofana, B. Identification and functional characterization of a flax UDP-glycosyltransferase glucosylating secoisolariciresinol (SECO) into secoisolariciresinol monoglucoside (SMG) and diglucoside (SDG). BMC Plant. Biol. 2014, 14, 82. [Google Scholar] [CrossRef] [Green Version]
- Teponno, R.B.; Kusari, S.; Spiteller, M. Recent advances in research on lignans and neolignans. Nat. Prod. Rep. 2016, 33, 1044–1092. [Google Scholar] [CrossRef] [Green Version]
- Fofana, B.; Ghose, K.; McCallum, J.; You, F.M.; Cloutier, S. UGT74S1 is the key player in controlling secoisolariciresinol diglucoside (SDG) formation in flax. BMC Plant Biol. 2017, 17, 35. [Google Scholar] [CrossRef] [Green Version]
- Gabr, A.M.M.; Mabrok, H.B.; Ghanem, K.Z.; Blaut, M.; Smetanska, I. Lignan accumulation in callus and Agrobacterium rhizogenes-mediated hairy root cultures of flax (Linum usitatissimum). Plant. Cell Tissue Organ. Cult. 2016, 126, 255–267. [Google Scholar] [CrossRef]
- Gabr, A.M.M.; Mabrok, H.B.; Abdel-Rahim, E.A.; El-Bahr, M.K.; Smetanska, I. Determination of lignans, phenolic acids and antioxidant capacity in transformed hairy root culture of Linum usitatissimum. Nat. Prod. Res. 2018, 32, 1867–1871. [Google Scholar] [CrossRef]
- Markulin, L.; Corbin, C.; Renouard, S.; Drouet, S.; Durpoix, C.; Mathieu, C.; Lopez, T.; Auguin, D.; Hano, C.; Lainé, É. Characterization of LuWRKY36, a flax transcription factor promoting secoisolariciresinol biosynthesis in response to Fusarium oxysporum elicitors in Linum usitatissimum L. hairy roots. Planta 2019, 250, 347–366. [Google Scholar] [CrossRef] [PubMed]
- Sertse, D.; You, F.M.; Ravichandran, S.; Cloutier, S. The Complex Genetic Architecture of Early Root and Shoot Traits in Flax Revealed by Genome-Wide Association Analyses. Front. Plant. Sci. 2019, 10, 1483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siegień, I.; Adamczuk, A.; Wróblewska, K. Light affects in vitro organogenesis of Linum usitatissimum L. and its cyanogenic potential. Acta Physiol. Plant. 2013, 35, 781–789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Millam, S.; Obert, B.; Pret’ová, A. Plant cell and biotechnology studies in Linum usitatissimum—A review. Plant. Cell. Tissue Organ. Cult. 2005, 82, 93–103. [Google Scholar] [CrossRef]
- Reis, R.V.; Borges, A.P.P.L.; Chierrito, T.P.C.; de Souto, E.R.; de Souza, L.M.; Iacomini, M.; de Oliveira, A.J.B.; Gonçalves, R.A.C. Establishment of adventitious root culture of Stevia rebaudiana Bertoni in a roller bottle system. Plant. Cell Tissue Organ. Cult. 2011, 106, 329–335. [Google Scholar] [CrossRef]
- Murthy, H.N.; Hahn, E.J.; Paek, K.Y. Adventitious roots and secondary metabolism. Chin. J. Biotechnol. 2008, 24, 711–716. [Google Scholar] [CrossRef]
- Khan, M.A.; Abbasi, B.H.; Shah, N.A.; Yücesan, B.; Ali, H. Analysis of metabolic variations throughout growth and development of adventitious roots in Silybum marianum L.(Milk thistle), a medicinal plant. Plant. Cell Tissue Organ. Cult. 2015, 123, 501–510. [Google Scholar] [CrossRef]
- Cui, X.-H.; Chakrabarty, D.; Lee, E.-J.; Paek, K.-Y. Production of adventitious roots and secondary metabolites by Hypericum perforatum L. in a bioreactor. Bioresour. Technol. 2010, 101, 4708–4716. [Google Scholar] [CrossRef]
- Jeong, J.-A.; Wu, C.-H.; Murthy, H.N.; Hahn, E.-J.; Paek, K.-Y. Application of an airlift bioreactor system for the production of adventitious root biomass and caffeic acid derivatives of Echinacea purpurea. Biotechnol. Bioprocess. Eng. 2009, 14, 91–98. [Google Scholar] [CrossRef]
- Praveen, N.; Murthy, H.N. Production of withanolide-A from adventitious root cultures of Withania somnifera. Acta Physiol. Plant. 2010, 32, 1017–1022. [Google Scholar] [CrossRef]
- Fazal, H.; Abbasi, B.H.; Ahmad, N. Optimization of adventitious root culture for production of biomass and secondary metabolites in Prunella vulgaris L. Appl. Biochem. Biotechnol. 2014, 174, 2086–2095. [Google Scholar] [CrossRef] [PubMed]
- Janowicz, J.; Niemann, J.; Wojciechowski, A. The effect of growth regulators on the regeneration ability of flax (Linum usitatissimum L.) hypocotyl explants in in vitro culture. Biotechnol. J. Biotechnol. Comput. Biol. Bionanotechnol. 2012, 93, 135–138. [Google Scholar] [CrossRef] [Green Version]
- Rasool, R.; Ganai, B.A.; Kamili, A.N.; Akbar, S. Antioxidant potential in callus culture of Artemisia amygdalina Decne. Nat. Prod. Res. 2012, 26, 2103–2106. [Google Scholar] [PubMed]
- Raj, D.; Kokotkiewicz, A.; Drys, A.; Luczkiewicz, M. Effect of plant growth regulators on the accumulation of indolizidine alkaloids in Securinega suffruticosa callus cultures. Plant. Cell Tissue Organ. Cult. 2015, 123, 39–45. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Hahn, E.-J.; Murthy, H.N.; Paek, K. Adventitious root growth and ginsenoside accumulation in Panax ginseng cultures as affected by methyl jasmonate. Biotechnol. Lett. 2004, 26, 1619–1622. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhu, W.-H.; Hu, Q.; He, X.-W. Enhanced indole alkaloid production in suspension compact callus clusters of Catharanthus roseus: Impacts of plant growth regulators and sucrose. Plant. Growth Regul. 2001, 33, 33–41. [Google Scholar] [CrossRef]
- Abbasi, B.H.; Siddiquah, A.; Tungmunnithum, D.; Bose, S.; Younas, M.; Garros, L.; Drouet, S.; Giglioli-Guivarc’h, N.; Hano, C. Isodon rugosus (Wall. ex Benth.) codd in vitro cultures: Establishment, phytochemical characterization and in vitro antioxidant and anti-aging activities. Int. J. Mol. Sci. 2019, 20, 452. [Google Scholar] [CrossRef] [Green Version]
- Kikowska, M.; Thiem, B.; Sliwinska, E.; Rewers, M.; Kowalczyk, M.; Stochmal, A.; Oleszek, W. The Effect of Nutritional Factors and Plant Growth Regulators on Micropropagation and Production of Phenolic Acids and Saponins from Plantlets and Adventitious Root Cultures of Eryngium maritimum L. J. Plant. Growth Regul. 2014, 33, 809–819. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.; Abbasi, B.H. Production of commercially important secondary metabolites and antioxidant activity in cell suspension cultures of Artemisia absinthium L. Ind. Crop. Prod. 2013, 49, 400–406. [Google Scholar] [CrossRef]
- Lindsey, K.; Yeoman, M.M. The relationship between growth rate, differentiation and alkaloid accumulation in cell cultures. J. Exp. Bot. 1983, 34, 1055–1065. [Google Scholar] [CrossRef]
- Rice-Evans, C.A.; Miller, N.J. Antioxidant activities of flavonoids as bioactive components of food. Biochem. Soc. Trans. 1996, 24, 790–794. [Google Scholar] [CrossRef]
- Misra, N.; Misra, P.; Datta, S.K.; Mehrotra, S. In vitro biosynthesis of antioxidants from Hemidesmus indicus R. Br. cultures. Vitr. Cell. Dev. Biol. 2005, 41, 285–290. [Google Scholar] [CrossRef]
- Anjum, S.; Abbasi, B.H. Thidiazuron-enhanced biosynthesis and antimicrobial efficacy of silver nanoparticles via improving phytochemical reducing potential in callus culture of Linum usitatissimum L. Int. J. Nanomed. 2016, 11, 715. [Google Scholar]
- Andreazza, N.L.; Abreu, I.N.; Sawaya, A.; Eberlin, M.N.; Mazzafera, P. Production of imidazole alkaloids in cell cultures of jaborandi as affected by the medium pH. Biotechnol. Lett. 2009, 31, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Zahir, A.; Abbasi, B.H.; Adil, M.; Anjum, S.; Zia, M. Synergistic effects of drought stress and photoperiods on phenology and secondary metabolism of Silybum marianum. Appl. Biochem. Biotechnol. 2014, 174, 693–707. [Google Scholar] [CrossRef] [PubMed]
- Struijs, K.; Vincken, J.-P.; Doeswijk, T.G.; Voragen, A.G.J.; Gruppen, H. The chain length of lignan macromolecule from flaxseed hulls is determined by the incorporation of coumaric acid glucosides and ferulic acid glucosides. Phytochemistry 2009, 70, 262–269. [Google Scholar] [CrossRef]
- Corbin, C. Etude de la Régulation Transcriptionnelle de la Synthèse des Lignanes du lin (Linum usitatissimum L.). PhD Thesis, Université d’Orléans, Orléans, France, 2015. [Google Scholar]
- Attoumbré, J.; Charlet, S.; Baltora-Rosset, S.; Hano, C.; Raynaud-Le Grandic, S.; Gillet, F.; Bensaddek, L.; Mesnard, F.; Fliniaux, M.A. High accumulation of dehydrodiconiferyl alcohol-4-β-D-glucoside in free and immobilized Linum usitatissimum cell cultures. Plant. Cell Rep. 2006. [Google Scholar] [CrossRef]
- Attoumbre, J.; Hano, C.; Mesnard, F.; Lamblin, F.; Bensaddek, L.; Raynaud-Le Grandic, S.; Laine, É.; Fliniaux, M.-A.; Baltora-Rosset, S. Identification by NMR and accumulation of a neolignan, the dehydrodiconiferyl alcohol-4-$β$-D-glucoside, in Linum usitatissimum cell cultures. Comptes. Rendus Chim. 2006, 9. [Google Scholar] [CrossRef]
- Hano, C.; Addi, M.; Fliniaux, O.; Bensaddek, L.; Duverger, E.; Mesnard, F.; Lamblin, F.; Lainé, E. Molecular characterization of cell death induced by a compatible interaction between Fusarium oxysporum f. sp. linii and flax (Linum usitatissimum) cells. Plant. Physiol. Biochem. 2008, 46, 590–600. [Google Scholar] [CrossRef]
- Corbin, C.; Renouard, S.; Lopez, T.; Lamblin, F.; Lainé, E.; Hano, C. Identification and characterization of cis-acting elements involved in the regulation of ABA- and/or GA-mediated LuPLR1 gene expression and lignan biosynthesis in flax (Linum usitatissimum L.) cell cultures. J. Plant. Physiol. 2013, 170, 516–522. [Google Scholar] [CrossRef]
- Corbin, C.; Decourtil, C.; Marosevic, D.; Bailly, M.; Lopez, T.; Renouard, S.; Doussot, J.; Dutilleul, C.; Auguin, D.; Giglioli-Guivarc’h, N.; et al. Role of protein farnesylation events in the ABA-mediated regulation of the Pinoresinol-Lariciresinol Reductase 1 (LuPLR1) gene expression and lignan biosynthesis in flax (Linum usitatissimum L.). Plant. Physiol. Biochem. 2013, 72, 96–111. [Google Scholar] [CrossRef] [PubMed]
- Markulin, L.; Drouet, S.; Corbin, C.; Decourtil, C.; Garros, L.; Renouard, S.; Lopez, T.; Mongelard, G.; Gutierrez, L.; Auguin, D.; et al. The control exerted by ABA on lignan biosynthesis in flax (Linum usitatissimum L.) is modulated by a Ca 2+ signal transduction involving the calmodulin-like LuCML15b. J. Plant. Physiol. 2019, 236, 74–87. [Google Scholar] [CrossRef] [PubMed]
- Beejmohun, V.; Fliniaux, O.; Hano, C.; Pilard, S.; Grand, E.; Lesur, D.; Cailleu, D.; Lamblin, F.; Lainé, E.; Kovensky, J.; et al. Coniferin dimerisation in lignan biosynthesis in flax cells. Phytochemistry 2007, 68. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, M.; Abbasi, B.H.; Garros, L.; Drouet, S.; Zahir, A.; Ahmad, W.; Giglioli-Guivarc’h, N.; Hano, C. Yeast-extract improved biosynthesis of lignans and neolignans in cell suspension cultures of Linum usitatissimum L. Plant. Cell. Tissue Organ. Cult. 2018, 135, 347–355. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Rukh, G.; Ahmad, N.; Rab, A.; Ahmad, N.; Fazal, H.; Akbar, F.; Ullah, I.; Mukhtar, S.; Samad, N. Photo-dependent somatic embryogenesis from non-embryogenic calli and its polyphenolics content in high-valued medicinal plant of Ajuga bracteosa. J. Photochem. Photobiol. B Biol. 2019, 190, 59–65. [Google Scholar] [CrossRef]
- Renouard, S.; Hano, C.; Corbin, C.; Fliniaux, O.; Lopez, T.; Montguillon, J.; Barakzoy, E.; Mesnard, F.; Lamblin, F.; Lainé, E. Cellulase-assisted release of secoisolariciresinol from extracts of flax (Linum usitatissimum) hulls and whole seeds. Food Chem. 2010, 122, 679–687. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Conc. (mg/L) | Days Required | Percentage Callogenesis (%) | Fresh Weight (g/L) | Dry Weight (g/L) | |
|---|---|---|---|---|---|
| IAA | 0.5 | 9 ± 0.13 | 55.28 ± 1.33 | 103.01 ± 4.14 | 4.77 ± 0.034 |
| 1.0 | 10 ± 0.11 | 42.03 ± 1.62 | 96.21 ± 2.92 | 4.41 ± 0.061 | |
| 2.0 | 10 ± 0.21 | - | - | - | |
| 3.0 | - | - | - | - | |
| NAA | 0.5 | 8 ± 0.16 | 95.21 ± 3.09 | 323.11 ± 7.43 | 12.65 ± 0.24 |
| 1.0 | 7 ± 0.19 | 98.07 ± 2.24 | 392.76 ± 8.27 | 13.26 ± 0.44 | |
| 2.0 | 7 ± 0.14 | 82.83 ± 3.15 | 277.34 ± 6. 29 | 11.15 ± 0.63 | |
| 3.0 | 9 ± 0.20 | 76.34 ± 2.03 | 228.19 ± 4.95 | 10.03 ± 1.07 | |
| 2,4-D | 0.5 | - | - | - | - |
| 1.0 | 10 ± 0.43 | 65.21 ± 2.09 | 146.01 ± 3.65 | 5.23 ± 0.32 | |
| 2.0 | 8 ± 0.61 | 74.09 ± 2.05 | 189.09 ± 3.22 | 6.01 ± 0.22 | |
| 3.0 | 7 ± 0.10 | 88.31 ± 2.85 | 206.04 ± 2.97 | 9.21 ± 0.50 | |
| IBA | 0.5 | 9 ± 0.25 | 42.53 ± 2.32 | 79.21 ± 4.61 | 3.81 ± 0.02 |
| 1.0 | 10 ± 0.12 | 38.47 ± 1.20 | 62.87 ± 4.22 | 3.02 ± 0.04 | |
| 2.0 | - | - | - | - | |
| 3.0 | - | - | - | - | |
| Control | MS0 | - | - | - | - |
| Conc. (mg/L) | Days Required | Percentage Rooting (%) | Fresh Weight (g/L) | Dry Weight (g/L) | |
|---|---|---|---|---|---|
| IAA | 0.5 | 11 ± 0.24 | 65.36 ± 2.32 | 61.23 ± 2.44 | 3.67 ± 0.08 |
| 1.0 | 12 ± 0.21 | 62.28 ± 1.48 | 56.12 ± 3.79 | 3.21 ± 0.03 | |
| 2.0 | - | - | - | - | |
| 3.0 | - | - | - | - | |
| NAA | 0.5 | 8 ± 0.13 | 100.00 ± 2.99 | 123.21 ± 4.57 | 10.52 ± 0.16 |
| 1.0 | 10 ± 0.36 | 86.52 ± 3.26 | 92.76 ± 2.38 | 7.26 ± 0.16 | |
| 2.0 | 10 ± 0.14 | 80.32 ± 2.65 | 77.34 ± 3. 34 | 4.15 ± 0.08 | |
| 3.0 | - | - | - | - | |
| IBA | 0.5 | 9 ± 0.45 | 78.52 ± 4.57 | 83.21 ± 3.65 | 4.81 ± 0.10 |
| 1.0 | 10 ± 0.22 | 72.59 ± 4.92 | 79.87 ± 5.32 | 4.62 ± 0.08 | |
| 2.0 | 11± 0.38 | 67.62 ± 3.25 | 61.25 ± 4.45 | 3.71 ± 0.06 | |
| 3.0 | - | - | - | - | |
| Control | MS0 | - | - | - | - |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anjum, S.; Komal, A.; Drouet, S.; Kausar, H.; Hano, C.; Abbasi, B.H. Feasible Production of Lignans and Neolignans in Root-Derived In Vitro Cultures of Flax (Linum usitatissimum L.). Plants 2020, 9, 409. https://doi.org/10.3390/plants9040409
Anjum S, Komal A, Drouet S, Kausar H, Hano C, Abbasi BH. Feasible Production of Lignans and Neolignans in Root-Derived In Vitro Cultures of Flax (Linum usitatissimum L.). Plants. 2020; 9(4):409. https://doi.org/10.3390/plants9040409
Chicago/Turabian StyleAnjum, Sumaira, Amna Komal, Samantha Drouet, Humera Kausar, Christophe Hano, and Bilal Haider Abbasi. 2020. "Feasible Production of Lignans and Neolignans in Root-Derived In Vitro Cultures of Flax (Linum usitatissimum L.)" Plants 9, no. 4: 409. https://doi.org/10.3390/plants9040409