Assessment of Genetic Diversity of the “Acquaviva Red Onion” (Allium cepa L.) Apulian Landrace

 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. Establishment of Acquaviva Red Onion Germplasm Collection and Morphological Characterization
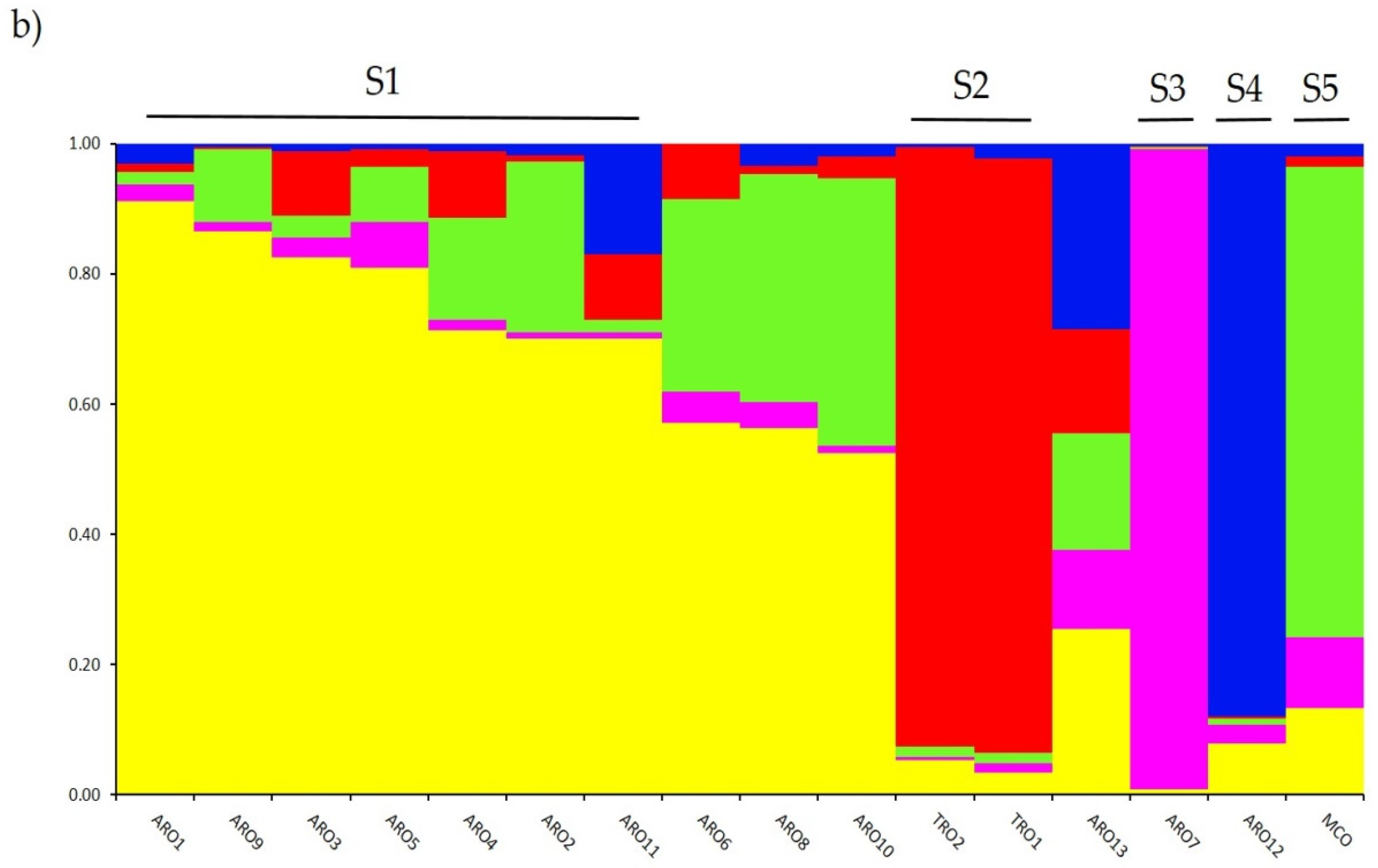
2.2. SSR Polymorphism and Genetic Relationships among Accessions
2.3. Analysis of Molecular Variance and Genetic Structure
2.4. Genetic Relationships among Populations
3. Discussion
4. Materials and Methods
4.1. Germplasm Collection, Plant Material, and DNA Extraction
4.2. SSR Analysis
4.3. Assessment of Genetic Diversity
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stearn, W.T. How many species of Allium are known? Kew Mag. 1992, 9, 180–182. [Google Scholar] [CrossRef]
- FAOSTAT. FAO Statistical Database. Available online: http://www.fao.org/2017 (accessed on 8 January 2019).
- Block, E. The chemistry of garlic and onion. Sci. Am. 1985, 252, 114–119. [Google Scholar] [CrossRef]
- Lee, B.; Jung, J.H.; Kim, H.S. Assessment of red onion on antioxidant activity in rat. Food Chem. Toxicol. 2012, 50, 3912–3919. [Google Scholar] [CrossRef]
- Lee, S.M.; Moon, J.; Chung, J.H.; Cha, Y.J.; Shin, M.J. Effect of quercetin-rich onion peel extracts on arterial thrombosis in rats. Food Chem. Toxicol. 2013, 57, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Yoshinari, O.; Shiojima, Y.; Igarashi, K. Anti-obesity effects of onion extract in zucker diabetic fatty rats. Nutrients 2012, 4, 1518–1526. [Google Scholar] [CrossRef]
- Akash, M.S.H.; Rehman, K.; Chen, S. Spice plant Allium cepa: Dietary supplement for treatment of type 2 diabetes mellitus. Nutrition 2014, 30, 1128–1137. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tian, W.X.; Ma, X.F. Inhibitory Effects of onion (Allium cepa L.) extract on proliferation of cancer cells and adipocytes via inhibiting fatty acid synthase. Asian Pac. J. Cancer Prev. 2012, 13, 5573–5579. [Google Scholar] [CrossRef] [PubMed]
- Lai, W.W.; Hsu, S.C.; Chueh, F.S.; Chen, Y.Y.; Yang, J.S.; Lin, J.P.; Lien, J.C.; Tsai, C.H.; Chung, J.G. Quercetin inhibits migration and invasion of SAS human oral cancer cells through inhibition of NF-kappaB and matrix metalloproteinase-2/-9 signaling pathways. Anticancer Res. 2013, 33, 1941–1950. [Google Scholar] [PubMed]
- Nicastro, H.L.; Ross, S.A.; Milner, J.A. Garlic and onions: Their cancer prevention properties. Cancer Prev. Res. 2015, 8, 181–189. [Google Scholar] [CrossRef] [Green Version]
- Forte, L.; Torricelli, P.; Boanini, E.; Gazzano, M.; Rubini, K.; Fini, M.; Bigi, A. Antioxidant and bone repair properties of quercetin-functionalized hydroxyapatite: An in vitro osteoblast-osteoclast-endothelial cell co-culture study. Acta Biomater. 2016, 32, 298–308. [Google Scholar] [CrossRef]
- Yamazaki, Y.; Iwasaki, K.; Mikami, M.; Yagihashi, A. Distribution of eleven flavor precursors, S-Alk(en)yl-L-cysteine derivatives, in seven Allium vegetables. Food Sci. Technol. Res. 2011, 17, 55–62. [Google Scholar] [CrossRef] [Green Version]
- Block, E. The organosulfur chemistry of the Genus Allium—Implications for the organic chemistry of sulphur. Angew. Chem. Int. Ed. Engl. 1992, 31, 1135–1178. [Google Scholar] [CrossRef]
- Griffiths, G.; Trueman, L.; Crowther, T.; Thomas, B.; Smith, B. Onions-A global benefit to health. Phytother. Res. 2002, 16, 603–615. [Google Scholar] [CrossRef]
- Schwimmer, S.; Weston, W.J. Enzymatic development of pyruvic acid in onion as a measure of pungency. J. Agric. Food Chem. 1961, 9, 301–304. [Google Scholar] [CrossRef]
- Ketter, C.A.T.; Randle, W.M. Pungency assessment in onions. In Tested Studies for Laboratory Teaching; Karcher, S.J., Ed.; Association for Biology Laboratory Education (ABLE): New York, NY, USA, 1998; Volume 19, pp. 177–196. [Google Scholar]
- Hanelt, P. Taxonomy, evolution, and history. In Onions and Allied Crops, Vol. I. Botany, Physiology and Genetics; Rabinowitch, H.D., Brewster, J.L., Eds.; CRC Press: Boca Raton, FL, USA, 1990; pp. 1–26. [Google Scholar]
- Rabinowitch, H.D.; Currah, L. Allium Crop Science: Recent Advances; CABI Publishing: Wallingford, UK, 2002. [Google Scholar]
- Mallor, C.; Carravedo, M.; Estopañan, G.; Mallor, F. Characterization of genetic resources of onion (Allium cepa L.) from Spanish secondary centre of diversity. Span. J. Agric. Res. 2011, 9, 144–155. [Google Scholar] [CrossRef]
- Ferioli, F.; D’Antuono, L.F. Evaluation of phenolics and cysteine sulfoxides in local onion and shallot germplasm from Italy and Ukraine. Genet. Resour. Crop Evol. 2016, 63, 601–614. [Google Scholar] [CrossRef]
- Petropoulos, S.A.; Fernandes, A.; Barros, L.; Ferreira, I.C.F.R.; Ntatsi, G. Morphological, nutritional and chemical description of ‘vatikiotiko’, an onion local landrace from Greece. Food Chem. 2015, 182, 156–163. [Google Scholar] [CrossRef] [Green Version]
- Liguori, L.; Adiletta, G.; Nazzaro, F.; Fratianni, F.; Di Matteo, M.; Albanese, D. Biochemical, antioxidant properties and antimicrobial activity of different onion varieties in the Mediterranean area. J. Food Meas. Charact. 2019, 13, 1232–1241. [Google Scholar] [CrossRef]
- Yoo, K.S.; Pike, L.; Crosby, K.; Jones, R.; Leskovar, D. Differences in onion pungency due to cultivars, growth environment, and bulb sizes. Sci. Hortic. 2006, 110, 144–149. [Google Scholar] [CrossRef]
- Beesk, N.; Perner, H.; Schwarz, D.; George, E.; Kroh, L.W.; Rohn, S. Distribution of quercetin-3, 4′-O-diglucoside, quercetin-4′-O-monoglucoside, and quercetin in different parts of the onion bulb (Allium cepa L.) influenced by genotype. Food Chem. 2010, 122, 566–571. [Google Scholar] [CrossRef]
- Caruso, G.; Conti, S.; Villari, G.; Borrelli, C.; Melchionna, G.; Minutolo, M.; Russo, G.; Amalfitano, C. Effects of transplanting time and plant density on yield, quality and antioxidant content of onion (Allium cepa L.) in southern Italy. Sci. Hortic. 2014, 166, 111–120. [Google Scholar] [CrossRef]
- Pérez-Gregorio, M.R.; Regueiro, J.; Simal-Gándara, J.; Rodrigues, A.S.; Almeida, D.P.F. Increasing the added-value of onions as a source of antioxidant flavonoids: A critical review. Crit. Rev. Food Sci. Nutr. 2014, 54, 1050–1062. [Google Scholar] [CrossRef] [PubMed]
- Pöhnl, T.; Schweiggert, R.M.; Carle, R. Impact of cultivation method and cultivar selection on soluble carbohydrates and pungent principles in onions (Allium cepa L.). J. Agric. Food Chem. 2018, 66, 12827–12835. [Google Scholar] [CrossRef] [PubMed]
- Tedesco, I.; Carbone, V.; Spagnuolo, C.; Minasi, P.; Russo, G.L. Identification and quantification of flavonoids from two southern Italian cultivars of Allium cepa L., Tropea (red onion) and Montoro (copper onion), and their capacity to protect human erythrocytes from oxidative stress. J. Agric. Food Chem. 2015, 63, 5229–5238. [Google Scholar] [CrossRef]
- Villano, C.; Esposito, S.; Carucci, F.; Frusciante, L.; Carputo, D.; Aversano, R. High-throughput genotyping in onion reveals structure of genetic diversity and informative SNPs useful for molecular breeding. Mol. Breed. 2019, 39, 5. [Google Scholar] [CrossRef]
- Mercati, F.; Longo, C.; Poma, D.; Araniti, F.; Lupini, A.; Mammano, M.M.; Fiore, M.C.; Abenavoli, M.R.; Sunseri, F. Genetic variation of an Italian long shelf-life tomato (Solanum lycopersicum L.) collection by using SSR and morphological fruit traits. Genet. Resour. Crop Evol. 2014, 62, 721–732. [Google Scholar] [CrossRef]
- González–Pérez, S.; Mallor, C.; Garcés–Claver, A.; Merino, F.; Taboada, A.; Rivera, A.; Pomar, F.; Perovic, D.; Silvar, C. Exploring genetic diversity and quality traits in a collection of onion (Allium cepa L.) landraces from north-west Spain. Genetika 2015, 47, 885–900. [Google Scholar] [CrossRef]
- Lotti, C.; Iovieno, P.; Centomani, I.; Marcotrigiano, A.R.; Fanelli, V.; Mimiola, G.; Summo, C.; Pavan, S.; Ricciardi, L. Genetic, bio-agronomic, and nutritional characterization of kale (Brassica oleracea L. var. acephala) diversity in Apulia, Southern Italy. Diversity 2018, 10, 25. [Google Scholar] [CrossRef] [Green Version]
- Bardaro, N.; Marcotrigiano, A.R.; Bracuto, V.; Mazzeo, R.; Ricciardi, F.; Lotti, C.; Pavan, S.; Ricciardi, L. Genetic analysis of resistance to Orobanche crenata (Forsk.) in a pea (Pisum sativum L.) low-strigolactone line. J. Plant Pathol. 2016, 98, 671–675. [Google Scholar]
- Wako, T.; Tsukazaki, H.; Yaguchi, S.; Yamashita, K.; Ito, S.; Shigyo, M. Mapping of quantitative trait loci for bolting time in bunching onion (Allium fistulosum L.). Euphytica 2016, 209, 537–546. [Google Scholar] [CrossRef]
- Dhaka, N.; Mukhopadhyay, A.; Paritosh, K.; Gupta, V.; Pental, D.; Pradhan, A.K. Identification of genic SSRs and construction of a SSR-based linkage map in Brassica juncea. Euphytica 2017, 213, 15. [Google Scholar] [CrossRef]
- Anandhan, S.; Mote, S.R.; Gopal, J. Evaluation of onion varietal identity using SSR markers. Seed Sci. Technol. 2014, 42, 279–285. [Google Scholar] [CrossRef]
- Mitrová, K.; Svoboda, P.; Ovesná, J. The selection and validation of a marker set for the differentiation of onion cultivars from the Czech Republic. Czech J. Genet. Plant Breed. 2015, 51, 62–67. [Google Scholar] [CrossRef] [Green Version]
- Di Rienzo, V.; Miazzi, M.M.; Fanelli, V.; Sabetta, W.; Montemurro, C. The preservation and characterization of Apulian olive germplasm biodiversity. Acta Hortic. 2018, 1199, 1–6. [Google Scholar] [CrossRef]
- Mallor, C.; Arnedo-Andrés, A.; Garcés-Claver, A. Assessing the genetic diversity of Spanish Allium cepa landraces for onion breeding using microsatellite markers. Sci. Hortic. 2014, 170, 24–31. [Google Scholar] [CrossRef]
- Rivera, A.; Mallor, C.; Garcés-Claver, A.; García-Ulloa, A.; Pomar, F.; Silvar, C. Assessing the genetic diversity in onion (Allium cepa L.) landraces from northwest Spain and comparison with the European variability. N. Z. J. Crop Hortic. 2016, 44, 103–120. [Google Scholar] [CrossRef]
- De Giovanni, C.; Pavan, S.; Taranto, F.; Di Rienzo, V.; Miazzi, M.M.; Marcotrigiano, A.R.; Mangini, G.; Montemurro, C.; Ricciardi, L.; Lotti, C. Genetic variation of a global germplasm collection of chickpea (Cicer arietinum L.) including Italian accessions at risk of genetic erosion. Physiol. Mol. Biol. Plants 2017, 23, 197–205. [Google Scholar] [CrossRef]
- Mazzeo, R.; Morgese, A.; Sonnante, G.; Zuluaga, D.L.; Pavan, S.; Ricciardi, L.; Lotti, C. Genetic diversity in broccoli rabe (Brassica rapa L. subsp. sylvestris (L.) Janch.) from Southern Italy. Sci. Hortic. 2019, 253, 140–146. [Google Scholar] [CrossRef]
- Jakše, M.; Martin, W.; McCallum, J.; Havey, M. Single nucleotide polymorphisms, indels, and simple sequence repeats for onion cultivar identification. J. Am. Soc. Hortic. Sci. 2005, 130, 912–917. [Google Scholar] [CrossRef]
- McCallum, J.; Thomson, S.; Pither-Joyce, M.; Kenel, F. Genetic diversity analysis and single-nucleotide polymorphism marker development in cultivated bulb onion based on expressed sequence tag–simple sequence repeat markers. J. Am. Soc. Hortic. Sci. 2008, 133, 810–818. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, S.; Pither-Joyce, M.; Wright, K.; Chen, L.; McCallum, J. Development of robust genomic simple sequence repeat markers for estimation of genetic diversity within and among bulb onion (Allium cepa L.) populations. Mol. Breed. 2012, 30, 1401–1411. [Google Scholar] [CrossRef]
- DeWoody, J.A.; Honeycutt, R.L.; Skow, L.C. Microsatellite markers in white tailed deer. J. Hered. 1995, 86, 317–319. [Google Scholar] [CrossRef] [PubMed]
- Khodadadi, M.; Hassanpanah, D. Iranian onion (Allium cepa L.) cultivars responses to inbreeding depression. World Appl. Sci. J. 2010, 11, 426–428. [Google Scholar]
- Abdou, R.; Bakasso, Y.; Saadou, M.; Baudoin, J.P.; Hardy, O.J. Genetic diversity of Niger onions (Allium cepa L.) assessed by simple sequence repeat markers (SSR). Acta Hortic. 2016, 1143, 77–90. [Google Scholar] [CrossRef]
- Pavan, S.; Lotti, C.; Marcotrigiano, A.R.; Mazzeo, R.; Bardaro, N.; Bracuto, V.; Ricciardi, F.; Taranto, F.; D’Agostino, N.; Schiavulli, A.; et al. A distinct genetic cluster in cultivated chickpea as revealed by genome-wide marker discovery and genotyping. Plant Genome 2017, 2017, 10. [Google Scholar] [CrossRef] [Green Version]
- Pavan, S.; Marcotrigiano, A.R.; Ciani, E.; Mazzeo, R.; Zonno, V.; Ruggieri, V.; Lotti, C.; Ricciardi, L. Genotyping-by-sequencing of a melon (Cucumis melo L.) germplasm collection from a secondary center of diversity highlights patterns of genetic variation and genomic features of different gene pools. BMC Genom. 2017, 18, 59. [Google Scholar] [CrossRef] [Green Version]
- Di Rienzo, V.; Sion, S.; Taranto, F.; D’Agostino, N.; Montemurro, C.; Fanelli, V.; Sabetta, W.; Boucheffa, S.; Tamendjari, A.; Pasqualone, A.; et al. Genetic flow among olive population within the Mediterranean basin. Peer J. 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Shepherd, L.D.; McLay, T.G. Two micro-scale protocols for the isolation of DNA from polysaccharide-rich plant tissue. J. Plant Res. 2011, 124, 311–314. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–14. [Google Scholar]
- Kuhl, J.C.; Cheung, F.; Qiaoping, Y.; Martin, W.; Zewdie, Y.; McCallum, J.; Catanach, A.; Rutherford, P.; Sink, K.C.; Jenderek, M.; et al. A unique set of 11,008 onion expressed sequence tags reveals expressed sequence and genomic differences between the monocot orders asparagales and poales. Plant Cell 2004, 16, 114–125. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Lee, H.R.; Hyun, J.Y.; Song, K.H.; Kim, K.H.; Kim, J.E.; Hur, C.G.; Harn, C.H. Marker development for onion genetic purity testing using SSR Finder. Korean J. Breed. Sci. 2012, 44, 421–432. [Google Scholar] [CrossRef]
- Schuelke, M. An economic method for fluorescent labeling of PCR fragments. Nat. Biotechnol. 2000, 18, 233–234. [Google Scholar] [CrossRef] [PubMed]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research: An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalinowski, S.T.; Taper, M.L.; Marshall, T.C. Revising how the computer program CERVUS accommodates genotyping error increases success in paternity assignment. Mol. Ecol. 2007, 16, 1099–1106. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Rosenberg, N.A.; Donnelly, P. Association mapping in structured populations. Am. J. Hum. Genet. 2000, 67, 170–181. [Google Scholar] [CrossRef] [Green Version]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Earl, D.; VonHoldt, B. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2011, 4. [Google Scholar] [CrossRef]
- Takezaki, N.; Nei, M.; Tamura, K. POPTREEW: Web version of POPTREE for constructing population trees from allele frequency data and computing some other quantities. Mol. Biol. Evol. 2014, 31, 1622–1624. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X. Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soluble Solid Content (Brix) | Pungency (µmol g−1 FW) | |||
|---|---|---|---|---|
| CODE | Mean | CV † (%) | Mean | CV † (%) |
| ARO1 | 6.25 D * | 5.65 | 5.84 ab * | 23.78 |
| ARO2 | 7.25 DC | 4.87 | 6.51 a | 22.98 |
| ARO3 | 7.50 BCD | 9.42 | 5.28 ab | 22.88 |
| ARO4 | 7.50 BCD | 0.00 | 6.97 a | 3.74 |
| ARO 5 | 7.50 BCD | 0.00 | 6.80 a | 9.68 |
| ARO6 | 6.25 D | 5.65 | 4.51 ab | 39.18 |
| ARO7 | 7.25 DC | 4.87 | 5.25 ab | 15.44 |
| ARO8 | 9.00 AB | 0.00 | 7.04 a | 3.49 |
| ARO9 | 8.25 ABC | 4.28 | 6.84 a | 0.15 |
| ARO10 | 7.00 DC | 0.00 | 5.94 ab | 6.57 |
| ARO11 | 9.50 A | 7.44 | 5.54 ab | 16.43 |
| ARO12 | 6.00 D | 0.00 | 4.91 ab | 9.70 |
| ARO13 | 9.50 A | 7.44 | 6.63 a | 24.93 |
| MCO | 6.00 D | 0.00 | 4.18 ab | 2.66 |
| TRO1 | 4.25 E | 8.31 | 2.80 b | 2.10 |
| TRO2 | 4.25 E | 8.31 | 4.28 ab | 4.79 |
| Locus. | Total Na | Size Range (bp) | PIC | Mean | |||||
|---|---|---|---|---|---|---|---|---|---|
| Na | Ne | Ho | He | I | Fis | ||||
| ACM91 | 4 | 189–205 | 0.40 | 2.63 | 1.72 | 0.38 | 0.39 | 0.66 | 0.04 |
| ACM101 | 4 | 229–241 | 0.52 | 2.94 | 2.37 | 0.53 | 0.56 | 0.92 | 0.06 |
| ACM132 | 11 | 186–248 | 0.67 | 5.38 | 2.78 | 0.55 | 0.61 | 1.20 | 0.09 |
| ACM138 | 5 | 242–272 | 0.66 | 3.69 | 2.82 | 0.62 | 0.61 | 1.09 | −0.02 |
| ACM147 | 2 | 264–266 | 0.37 | 1.94 | 1.83 | 0.44 | 0.44 | 0.62 | −0.01 |
| ACM152 | 4 | 228–244 | 0.25 | 2.38 | 1.41 | 0.25 | 0.27 | 0.45 | 0.07 |
| ACM235 | 4 | 286–298 | 0.41 | 2.81 | 1.77 | 0.44 | 0.41 | 0.72 | −0.06 |
| ACM446 | 6 | 108–120 | 0.56 | 3.50 | 2.48 | 0.49 | 0.58 | 1.01 | 0.16 |
| ACM449 | 8 | 120–140 | 0.66 | 4.88 | 2.82 | 0.62 | 0.61 | 1.18 | −0.03 |
| ACM463 | 5 | 202–210 | 0.47 | 3.38 | 1.95 | 0.46 | 0.48 | 0.83 | 0.05 |
| ACM504 | 2 | 188–192 | 0.33 | 1.94 | 1.64 | 0.30 | 0.37 | 0.54 | 0.20 |
| Mean | 5 | 0.48 | 3.22 | 2.15 | 0.46 | 0.48 | 0.84 | 0.05 | |
| Source | df | Sum of Squares | Variance Estimation | Variance (%) | ΦPT | P |
|---|---|---|---|---|---|---|
| Among populations | 15 | 458.63 | 1.16 | 13% | 0.134 | 0.001 |
| Within populations | 304 | 2272.99 | 7.50 | 87% | ||
| Total | 319 | 2731.62 | 8.66 |
| Code | Name | GPS Coordinates | Gene Bank † |
|---|---|---|---|
| ARO1 | Cipolla rossa di Acquaviva | 40°54′21.708″ N 16°49′1.631″ E | Di.S.S.P.A. |
| ARO2 | Cipolla rossa di Acquaviva | 40°53′14.28″ N 16°48′56.879″ E | Di.S.S.P.A. |
| ARO3 | Cipolla rossa di Acquaviva | 40°54′11.304″ N 16°49′13.079″ E | Di.S.S.P.A. |
| ARO4 | Cipolla rossa di Acquaviva | 40°54′3.348″ N 16°40′27.011″ E | Di.S.S.P.A. |
| ARO5 | Cipolla rossa di Acquaviva | 40°51′59.76″ N 16°53′0.527″ E | Di.S.S.P.A. |
| ARO6 | Cipolla rossa di Acquaviva | 40°52′48.72″ N 16°49′43.247″ E | Di.S.S.P.A. |
| ARO7 | Cipolla rossa di Acquaviva | 40°53′13.47″ N 16°50′23.783″ E | Di.S.S.P.A. |
| ARO8 | Cipolla rossa di Acquaviva | 40°53′18.816″ N 16°49′33.888″ E | Di.S.S.P.A. |
| ARO9 | Cipolla rossa di Acquaviva | 40°54′51.372″ N 16°49′3.504″ E | Di.S.S.P.A. |
| ARO10 | Cipolla rossa di Acquaviva | 40°54′1.188″ N 16°49′24.311″ E | Di.S.S.P.A. |
| ARO11 | Cipolla rossa di Acquaviva | 40°52′49.8″ N 16°49′48.575″ E | Di.S.S.P.A. |
| ARO12 | Cipolla rossa di Acquaviva | 40°52′38.892″ N 16°49′28.379″ E | Di.S.S.P.A. |
| ARO13 | Cipolla rossa di Acquaviva | 40°53′21.768″ N 16°49′29.711″ E | Di.S.S.P.A. |
| TRO1 | Cipolla rossa lunga di Tropea | - | Di.S.S.P.A. |
| TRO2 | Cipolla rossa tonda di Tropea | - | Di.S.S.P.A. |
| MCO | Cipolla ramata di Montoro | - | Di.S.S.P.A. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ricciardi, L.; Mazzeo, R.; Marcotrigiano, A.R.; Rainaldi, G.; Iovieno, P.; Zonno, V.; Pavan, S.; Lotti, C. Assessment of Genetic Diversity of the “Acquaviva Red Onion” (Allium cepa L.) Apulian Landrace. Plants 2020, 9, 260. https://doi.org/10.3390/plants9020260
Ricciardi L, Mazzeo R, Marcotrigiano AR, Rainaldi G, Iovieno P, Zonno V, Pavan S, Lotti C. Assessment of Genetic Diversity of the “Acquaviva Red Onion” (Allium cepa L.) Apulian Landrace. Plants. 2020; 9(2):260. https://doi.org/10.3390/plants9020260
Chicago/Turabian StyleRicciardi, Luigi, Rosa Mazzeo, Angelo Raffaele Marcotrigiano, Guglielmo Rainaldi, Paolo Iovieno, Vito Zonno, Stefano Pavan, and Concetta Lotti. 2020. "Assessment of Genetic Diversity of the “Acquaviva Red Onion” (Allium cepa L.) Apulian Landrace" Plants 9, no. 2: 260. https://doi.org/10.3390/plants9020260